
Ashraf M.A.S. Goda
Fish Nutrition Laboratory, National Institute of Oceanography and Fisheries (NIOF), Cairo, Egypt
Journal of Fisheries and Aquatic Science
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 248-263







ABSTRACT
Two feeding trails were conducted with Nile tilapia Oreochromis niloticus fingerlings to determine the effect of dietary phytase level with partial or complete replacement of fish meal (FM) by soybean meal (SBM) on growth, feed utilization, nutrients digestibility as well as dietary phosphorus discharge. A total of 960 Nile tilapia fingerlings with an average body weight of 18.8±0.3 g were divided in the twenty-experimental treatments (duplicate for each treatment). The experiment was conducted for 13 week. Five basal diets were formulated to be isonitrogenous 30% crude protein and isocaloric 19.5 MJ kg-1 diet gross energy. The control diet (diet 1) was prepared with FM as the main protein source. Diets 2-5 each SBM were substituted, for an isonitrogenous amount of FM control diet, to provide 25% (diet 2), 50% (diet 3), 75% (diet 4) or 100% (diet 5) of the FM crude protein. Another three groups of similar previous five basal diets were formulated and supplemented with 1000, 1500, 2000 FTU kg-1 diet, respectively of phytase enzyme. The present results suggest that a 25% substitution of dietary FM with SBM in Nile tilapia fingerlings diets has enhance growth performance and feed efficiency. With increasing dietary SBM levels; comparably lower growth indices were obtained compared to 100%-FM control diet. Irrespective of dietary SBM levels, supplementation of microbial phytase at 1000 FTU kg-1 level has demonstrated to be efficient for increasing dietary phosphorus supply for Nile tilapia fingerlings, thereby reducing the phosphorus effluent from aquaculture facilities. This may have to be taken into consideration for ongoing feeding strategies for the control of waste discharge.
PDF Abstract XML References Citation
How to cite this article
Ashraf M.A.S. Goda, 2007. Effect of Dietary Soybean Meal and Phytase Levels on Growth, Feed Utilization and Phosphorus Discharge for Nile tilapia Oreochromis niloticus (L.). Journal of Fisheries and Aquatic Science, 2: 248-263.
DOI: 10.3923/jfas.2007.248.263
URL: https://scialert.net/abstract/?doi=jfas.2007.248.263
DOI: 10.3923/jfas.2007.248.263
URL: https://scialert.net/abstract/?doi=jfas.2007.248.263
INTRODUCTION
Aquaculture feeds typically represent up to 70% of variable operating production costs, depending on the intensity of the operation (Muzinic et al., 2006). Nowadays, a priority area of research in aquaculture nutrition is the reduction and possible elimination of Fish Meal (FM) and fish oil will fully expected change in the future with more limitation on the use of these ingredients (Craig, 2004). The replacement of fish meal with plant or grain by-products will become increasingly important for the development of low cost fish feeds. One of the major problems associated with the use of plant by-products in fish feed is the presence of anti-nutritional factors, like phytic acid or phytates and enzyme inhibitors. Phytic acid (1,2,3,5/4,6-hexakis (dihydrogen phosphate) myoinositol) is the major phosphorus (P) storage compound in plant seeds and can account for up to 70% of total P (Lall, 1991; Eeckhout and De Paepe, 1994). Soluble inorganic and cellular P (P bound in nucleic acids, phosphorylated proteins, phospholipids, phosphor-sugars) represents the remaining P (Baruah et al., 2004).
Because of the high density of negatively charged phosphate groups, phytate chalets with mineral cations like potassium (K), magnesium (Mg), calcium (Ca), zinc (Zn), iron (Fe), copper (Cu) and forms poorly soluble complexes (Satoh et al., 1989; Hossain and Jauncey, 1993; Papatryphon et al., 1999; Riche et al., 2001; Goda et al., 2002; Nwanna, 2004; Yoo et al., 2005). Phytates also forms complexes with proteins and Amino Acid (AA). The amino groups present on the side chain of the AA are one of functional groups involved in protein-phytate interaction, thereby decreasing the digestibility of proteins and availability of AA (Riche et al., 2001). These salts of phytic acid are known as phytins and their availability and digestibility to monogastric animals including fish is very limited due to the lack of intestinal phytase (Pointillart et al., 1987). Both phytate-protein (binary complex) and phytate-mineral-protein (ternary complex) complexes can occur for tilapia (Reddy et al., 1989). Riche and Garling (2004) cited that in acidic environments, such as the tilapia stomach (pH 1-2), half of the phosphate moieties are negatively charged. This favours binding of soluble proteins at ε-amino groups on lysine (Lys), imidazole groups on histidine (His) and gaunidyl groups on arginine (Arg). In alkaline environments, such as tilapia intestine (pH 8.5-8.8), ternary complexes are favoured. Both complexes are resistant to proteolytic digestion (Vaintraub and Bulmaga, 1991; Caldwell, 1992). Finally, dietary phytate-phosphorus compound is excreted into the aquatic environment. However, a poor degradation of phytates leads to detrimental effects on the aquatic environment like eutrophication effects on freshwater or saltwater bodies, causing algal growth (Persson, 1991).
Soybean is considered the best plant protein for meeting the essential amino acid requirements of tilapia and other fish species commercially grown. Soybean meal (SBM) contains anti-nutritional factors (ANFs) notably; phytic acid, protease inhibitors and haemagglutins, tannins, gossypols, anti-vitamins and anti-enzymes which reduce their biological value and nutrient utilization and often result in histopathological abnormalities in fish (Wassef et al., 1988; Liener, 1994; Tacon, 1997). Therefore, the reduction or elimination of the levels of ANFs in soybean meal would improve its nutrient bioavailability, digestibility and absorption by fishes. However, reports of growth performance and feed efficiency in tilapia species fed soy products are conflicting. Poor performance is generally noticed in direct relationship to level of dietary SBM incorporation (Shiau et al., 1990; El-Dahhar and El-Shazly, 1993; Abdelghany, 2003; El-Sayed, 2006).
Tilapias as a group of fish species are highly suitable for low cost aquaculture. It is will documented that, tilapias lack the intestinal enzyme phytase (EC 3.1.3.8) that hydrolyze phytate. Phytase is a specific enzyme for phytate hydrolysis. However, exogenous phytase has been used successfully to hydrolyze phytate and increasing nutrient digestibility (Mroz et al., 1994; Teskeredñi� et al., 1995; Riche and Brown, 1996; Riche et al., 2001; Liebert and Portz, 2005). Phytase pre-treated diets fed to rainbow trout Oncorhynchus mykiss (Cain and Garling, 1995; Vielma et al., 2002) and Atlantic salmon, Salmo salar (Storebakken et al., 1998) resulted in an increased growth which attributed to improved protein utilization as demonstrated in terrestrial animals (Mroz et al., 1994; Sebastian et al., 1997; Martin et al., 1998). Addition of microbial phytase has been reported to improve the utilization and bioavailability of plant phosphorus in fish diets and, hence, there is less discharge into the aquatic environment, thereby causing less water pollution problems (Forster et al., 1999; Riche et al., 2001; Robinson et al., 2002; Liebert and Portz, 2005).
Egyptian aquaculture has developed rapidly in recent years. Tilapia is one of most widely cultured species in Egypt. The total aquaculture production of tilapia increased from 24,916 Metric Tons (mt) in 1990 to 486,000, mt year-1(GAFRD, 2006) and accounted for 55% of total production (876,000 mt year-1). One of the major problems confronting Egyptian fish production is the increasing cost and short supply of fish meal. Thus, replacing portions of FM with SBM in fish diets is a promising economical alternative for the aquaculture business. Otherwise, by applying phytase in SBM-based diets, it will not only reduce fish diet costs, but also reduce environmental pollution due to lower discharges of phosphorus and other minerals from fish farms.
Two growth and feed efficiency measurement trails were conducted with a completely randomized factorial design (4x5) experiment: The purpose of first trail (laboratory study) was to determine the influence of dietary phytase levels with partial or complete replacement of FM by SBM of different experimental diets on apparent nutrients digestibility as well as phosphorus utilization and discharges. The second trail (field study) was conducted to determine the effect of previous experimental test diets on growth, feed utilization and carcass composition of Nile tilapia O. niloticus fingerlings in concrete net-pen culture.
MATERIALS AND METHODS
Growth Trial
This study was performed at the Experimental Fish Farm, El-Kanater-El-Khayria, National Institute of Oceanography and Fisheries (NIOF), Kalubiya Governorate, Egypt. A total of 960 Nile tilapia O. niloticus fingerlings with an average weight of 18.8±0.3 g were obtained from Nawa Farm, Kalubiya Governorate, Egypt. In March-15-2005, the fingerlings of Nile tilapia were stocked into four cement ponds (40 m3 each). Each pond was divided into ten equal pens by netting (4 m3 each) and each pen was stocked with 24 fish (Duplicate pen for each treatment). Concrete ponds were supplied with freshwater from the Darawa irrigation Baranch, Kalubiya, Governorate, Egypt. The turnover rate of water was about 0.3 m3 day-1 pond-1 and fish was held under natural light conditions (12:12 h light: dark schedule). Fish were acclimatized to experimental conditions for 2 weeks prior to the start of the experiment. During adaptation period, fish was fed a commercial tilapia diet (28% CP, ZooControl Company, 6 October City, Cairo, Egypt).
Five basal diets were formulated to be isonitrogenous 30% crude protein and isocaloric 19.5 MJ kg-1 diet gross energy. The control diet (diet 1) was prepared with FM as the main protein source. Diets 2-5 each SBM was substituted, for an isonitrogenous amount of FM control diet, to provide 25% (diet 2), 50% (diet 3), 75% (diet 4) or 100% (diet 5) of the FM crude protein (Table 1). Crystalline L-lysine and DL-Methionine (Met) were added to the experimental diets containing SBM to cover Lysine and total sulphur amino acid requirements for O. niloticus (NRC, 1993).
Another three groups of similar previous five basal diets were formulated and supplemented with 1000, 1500, 2000 FTU kg-1 diet of phytase enzyme (Phytasonovo®). The enzyme product was kindly provided from Novo Nordisk, Denmark (1 g of Phytasonovo® content 2500 FTU). One unit of phytase (FTU) is defined as the quantity of enzyme that liberates one micro mol of inorganic orthophosphorus (Pi) minuet from 5.1 mmol L-1 sodium phytate at pH 5.5 and 37°C (Engelen et al., 1994). The proper amount of phytase was dissolved in 40 mL of distilled water. The twenty-experimental test diets were processed in laboratory by blending the dry ingredients with the proper amount of water phytase solution into a homogenous mixture for 10 min. Pellets of 2 mm were made in Sprout- Waldron laboratory pellet mill (CPM, California Pellet Mill Co., San Francisco, California, CA, USA). The pelleting temperature did not exceed 40°C. After pelleting, all diets were dried at 45°C for 48 h, packed in cellophane bags and cooled at 4°C prior to use. The FM (999 Con-Kix Fish Meal, Triple Nine Fish Protein a.m.b.a., Thyborøn, Denmark) used in this study was obtained from the ZooControl Company, 6 October City, Cairo, Egypt. All feed-grade ingredients including SBM and yellow corn were already purchased from local markets (ISLAMIC company (APICO), Giza, Egypt).
Fish were fed at a level of 3% of body weight twice a day (9.00 and 12.00 h) for 13 week (90 days). Fish were fed 7 days a week, except for days fish were weighed. From each experimental treatment fish were weighed collectively every two weeks, average fish weight was calculated and the amount of daily diet was adjusted accordingly. However, at the beginning of the experiment, three groups of 5 fish each were sacrificed and used for assessment of their initial proximate body composition. In addition, at the end of the experimental, random samples of 5 fish from each treatments (n = 3) were used to assess the final proximate body composition.
| Table 1: | Formulation and proximate analysis of experimental basal diets |
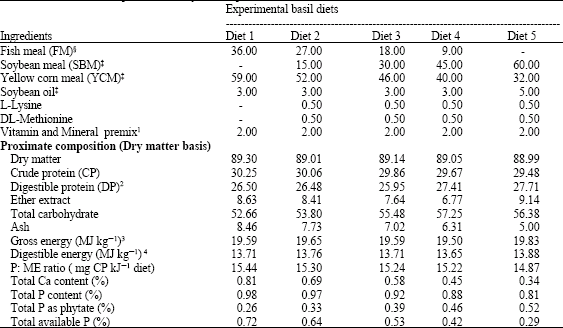 | |
| Diet 1: Fish Meal 100% (FM), Diet 2: FM 75%: SBM: 25%, Diet 3: FM 50%: SBM: 50% , Diet 4: FM 25%: SBM: 75%, Diet 5: Soybean meal 100% (SBM), §999 Con-Kix Fish Meal, Triple Nine Fish Protein a.m.b.a., Thyborøn, Denmark, ‡Supply from local markets (Animal Production Islamic Company (APICO), Dokki-El-Giza, Egypt). 1Vitamin and mineral mix (supplements per kg of the mixed feed): vitamin A, 45000 IU, vitamin D3, 4500 IU; vitamin E, 400 mg, vitamin B1, 30 mg, vitamin B2, 40 mg; vitamin B6, 40 mg; vitamin B12, 80 mcg; vitamin K3, 15 mg; ascorbic acid, 750 mg; nicotinic acid, 300 mg, Ca-pantothenate, 100 mg; folic acid, 10 mg; biotin, 3 mg; inositol, 500 mg; p-amino benzoic acid, 200 mg, Ca, 2.1 g; Fe, 250 mg; Mn, 40 mg; Zn, 60 mg; I, 4 mg; Cu, 12 mg; Se, 0.3 mg, Co, 2 mg, 2Digestible protein was calculated using values of 90.2%, 96.2 and 75.1% for crude protein content of FM, SBM and YC, respectively according to Sklan et al., 2004, 3Calculated using gross calorific values of 23.6, 17.2 and 39.5 kJ g-1 for protein, carbohydrate and fat, respectively according to Brett (1973), 4DE was calculated as 70% of calculated dietary GE (Hepher et al., 1983) | |
Digestibility Study
The apparent digestibility coefficient (ADC) was measured in a thermoregulated recirculation-glass water system. A total of 600 Nile tilapia O. niloticus fingerlings with an average weight of 18.2±0.5 g were randomly stocked into 60 glass aquaria (70x45x40 cm each) with a capacity of 45 L of water. The glass aquaria's were filled with dechlorinated tap water. Water was continuously aerated to supply oxygen. After two weeks of fish acclimation, the experimental fish were stocked as 10 fish aquaria-1 (three replicate for each treatment). The aquaria were cleaned daily from fecal matters and replacing 100% of the water. All fish in each aquaria were collectively weighed every week and the new feeding rate was re-adjusted. The water quality indices of experimental aquaria were measured at 09.00 h according to procedures of AOAC (1995). Temperature was maintained at 28±1.0°C. Dissolved Oxygen (DO) was measured daily using YSI model 56 oxygen meter (Yellow Springs Instrument, Yellow Springs, OH, USA)and pH at 8.00 h. Ammonia concentration was maintained at less than 0.3 mg L-1. Fish was maintained on an artificial light photoperiod of 12:12 h light: dark schedule.
Fish were fed to satiation three times a day (9:00, 11:00 and 14:00 h). Mean maximum intake was empirically determined as 4.4 g aquaria/day. To optimize feed utilization during the experimental period, fish were offered 80% on (a dry matter basis) of their estimated maximum consumption rate. This rate corresponded with the suggested feeding rates for tilapia (NRC, 1993; Riche et al., 2001). This feeding trial lasted for 21 days. The twenty-experimental diets, as described for growth experiment, were formulated as shown in Table 1 using chromic oxide (Cr2O3) as an external marker at a level of 0.05% of diet. Feaces samples were collected according to procedure of Nwanna (2004) using sedimentation technique for two weeks. The samples were collected every 6 h after last feeding by siphoning with a 5 mm diameter tube. The collected samples were dried at 105°C for 24 h, pooled for each treatment and analyzed for proximate composition according to methodology of AOAC (1995). The gross energy was determined with an adiabatic bomb calorimeter (Model 1272, Parr Instruments Inc., Moline, IL, USA) using benzoic acid as the slandered substance. The following equation determined the apparent digestibility coefficients (ADC) of the experimental diets according to Austreng et al. (1988):
 |
Growth and Feed Utilization Indices
Weight gain (WG), specific growth rate (SGR), feed conversion ratio (FCR), protein productive value (PPV), fat retention (FR) and energy retention (ER) were calculated using the following equations:
WG = Final body weight (g) - Initial body weight (g).
SGR = [(ln FBW - ln IBW)] period days-1x100,
where:
| FBW is final body weight (g); IBW is initial body weight (g); ln | = | natural logarithmic. |
| FCR | = | feed intake (g)/weight gain (g). |
| PPV | = | (Retained protein (g) /protein intake (g)) x100. |
| FR | = | (Retained fat (g) /fat intake (g)) x100. |
| ER | = | (Retained energy (kJ) /energy intake (kJ)) x100. |
Water Phosphorus Samples
Water phosphorus (P) samples were collected for the last 15 days of digestibility experiment. After the last meal provided, water samples from aquaria inlet (blank sample) and from all experimental aquaria's outlet were collected, (100 mL each) and were stabilized with 0.5 mL concentrated sulfuric acid (Bureau and Cho, 1999). Total phosphorus content was determined in all water samples taken by using the total phosphorus vanadomolybophosphoric acid colorimetric method (APHA, 1985).
The total waste discharge of phosphorus was calculated according to Cho et al. (1994) as:
Total P Discharge (g kg/ fish) = Total solid waste (TSW) + Total dissolved waste (TDW)
Where:
Total solid waste (TSW) = (Feed intake (kg)x ( (1- ADC) + Feed wastage ( kg , FW) .
Total dissolved waste (TDW) = Feed phosphorus consumed x ADC - body phosphorus retained.
ADC = Apparent digestibility coefficients.
Analytical Procedures
Proximate composition of fish, diets and feaces samples were determined according to standard methods of AOAC (1995). Analyses of samples were made on dry matter basis after drying in an oven at 105°C for 24 h. Ash was determined by incineration at 550°C for 12 h, crude protein by micro-Kjeldhal method, % N x 6.25 (using a Kjeltech autoanalyzer, Model 1030, Tecator, Höganäs, Sweden), crude fat by Soxhlet extraction with diethyl ether (40-60°C). The concentration of chromic oxide was measured by spectro-photomerically according to Furukawa and Tsukahtara (1966). Gross energy (GE) content of diet and carcass samples was calculated according to Brett (1973) by applying the factor of 23.6, 39.5 and 17.2 kJ g-1 for crude protein, crude fat and total carbohydrate, respectively. The Digestible Energy (DE) was estimated by applying the coefficient of 70% to convert determined GE to DE according to Hepher et al. (1983). Phytic acid was determined in all ingredients according to procedure of Wheeler and Ferrel (1971).
Statistical Analysis
The data of two growth and feed efficiency trails were analyzed with ANOVA by a completely randomized factorial design (4x5) experiment using MSTAT version 4 software (MSTAT, 1987). Duncan’s multiple range test was used to compare differences between individual means when significant ANOVA F values were determined (Duncan, 1955). Treatment effects were considered significant at (p≤0.05). The percentage values were transformed to their arc sign values before analysis. The original percentages values though, are listed in the Table 1-5.
RESULTS
Over the 13 week feeding periods, net-pen water quality demonstrated that water temperature ranged from 27.5-28.1°C, Dissolved Oxygen (DO) from 5.3 to 6.4 mg L-1, pH from 6.5-8.2, alkalinity from 165-170 mg L-1 and ammonia (NH3) from 0.22-0.28 mg L-1.
Experimental diets (Table 1) were formulated to contain 30% crude protein and gross energy (DE) was 19.5 MJ kg-1 diets. Proximate composition of all test diets was almost similarly closely related. The proximate analysis of diets showed that, with increasing the dietary SBM level up to 100%, the total phosphorus decreased linearly ranged from 980 mg P 100 g-1 to 810 mg P 100 g-1 diets, whereas dietary phosphorus as phytate was increased ranged from 260 mg P 100g-1 to 520 mg P 100g-1 diets and the dietary available P was decreased ranged from 720 mg P 100 g-1 to 290 mg P100 g-1 diets Growth indices (Table 2) showed that final body weight (FBW), feed intake (FI), weight gain (WG) and specific growth rate (SGR) were all significantly higher (p≤0.05) when Nile tilapia fed 25%-SBM diet. The trend of variation was towards decrease with increasing dietary SBM levels. However, fish fed 75%-SBM diet recorded the lowest significant (p≤0.05) values of FBW, FI, WG and SGR. Therefore, a small inclusion of plant protein in tilapia diets results in increased growth efficiency. Irrespective of dietary SBM levels, fish fed diet supplemented with phytase at 1000 FTU kg-1 showed the highest (p≥0.05) values of FBW, FI, WG and SGR, meanwhile the lowest (p≤0.05) value was recorded with the diet supplemented with phytase at 1500 FTU kg-1.
The highest (p≤0.05) feed conversion ratio (FCR) was shown when Nile tilapia fed 75%-SBM diet; whereas fish fed either 25%-SBM or 100%-FM diets recorded the better significant (p≤0.05) FCR values. No significant effect was observed for FCR with different dietary phytase levels. However, the higher feed intake (p≤0.05) value was observed for Nile tilapia fed diet supplemented with phytase at a level of 1000 FTU kg-1.
The influence of different experimental diets, on protein productive value (PPV), fat retention (FR) and energy retention (ER) of tilapia are shown in Table 2. The highest (p<0.05) values of PPV, FR and ER were observed for fish fed diets with 100%-FM, 50%-SBM and 100%-FM, respectively. Irrespective of dietary FM replacement, fish fed dietary phytase treated had not showed any differences in PPV and ER, while FR was observed the highest values (p≥0.05) with fish fed dietary phytase treated at level of 2000 FTU kg-1 .
The proximate compositions of final whole fish are shown in Table 3. No significant difference (p≤0.05) in whole body contents of moisture%, crude protein (CP), ether extract (EE), ash and gross energy (GE) were observed as affected by different dietary FM replacement levels with SBM or with different dietary phytase levels.
The apparent digestibility coefficients (ADCs) of nutrients for different experimental diets are illustrated in Table 4. The ADC of CP, GE and P were noticed to be the highest with Nile tilapia when fed 50%-SBM diet, whereas fish fed with 100%-FM diet showed the highest significant (p≤0.05) value for apparent digestibility of EE. Irrespective of dietary FM replacement levels by SBM, the higher ADC (p≤0.05) values for CP, EE and GE were observed for diet supplemented with 1000 FTU kg-1 phytase. Meanwhile, ADC of phosphorus has improved to a highest significant level for both dietary In contrast, the lowest ADC of protein was showed for diet supplemented with 2000 FTU kg-1 phytase, while, control diet was recorded the lowest ADC value of gross energy, lipid and phosphorus.
| Table 2: | Effect of dietary fish meal (FM) replacement level by SBM and phytase supplementation level on growth performance and feed utilization indices (mean±SD) for Nile tilapia fingerlings |
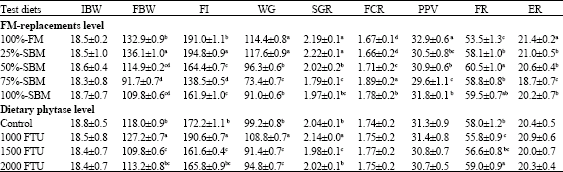 | |
| IBW: Initial Body Weight (g), FBW: Final Body Weight (g); FI: Feed Intake; WG: Weight Gain; SGR: Specific Growth Rate (% days), FCR: Feed Conversion Ratio, PPV: Protein Productive Value; FR: Fat Retention, ER: Energy Retention. Mean in the same column sharing the same superscript are not significantly different (p≥0.05) | |
| Table 3: | Effect of dietary fish meal (FM) replacement level by SBM and phytase supplementation level on proximate body composition (mean±SD) of Nile tilapia fingerlings |
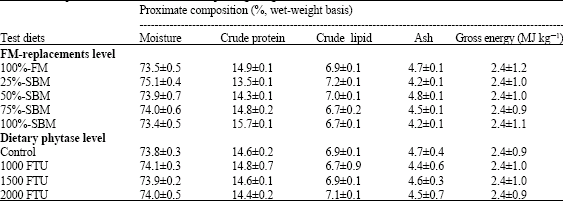 | |
| Mean in the same column sharing the same superscript are not significantly different (p≥0.05) | |
| Table 4: | Effect of dietary fish meal (FM) replacement level by SBM and phytase supplementation level on apparent digestibility coefficient (ADC) of different dietary nutrients content (mean±SD) for Nile tilapia fingerlings |
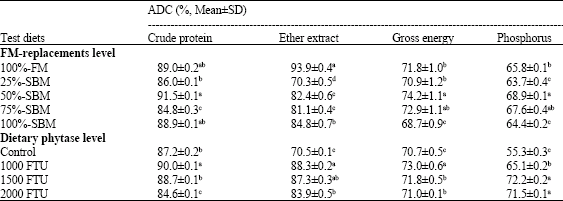 | |
| Mean in the same row sharing the same superscript are not significantly different (p≥0.05) | |
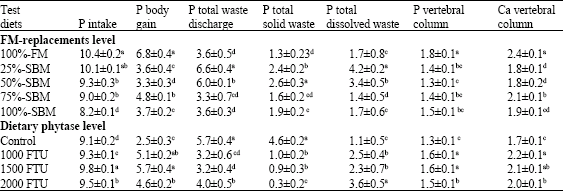
The P intake, P body gain, P total waste discharge, P total solid waste, P total dissolved waste, P vertebral column and Ca vertebral column are presented in Table 5. Nile tilapia fed 100%-FM control diet showed the highest (p≥0.05) values for P intake, P body gain, P vertebral column and Ca vertebral column associated with lowest values of P total waste discharge and P total solid waste, while the lower significant (p≥0.05) values of P body gain, P vertebral column and Ca vertebral column were observed for fish fed 50%-SBM diet.
| Table 5: | Effect of dietary fish meal (FM) replacement level by SBM and phytase supplementation level on dietary phosphors budget (P g kg fish) for Nile tilapia fingerlings |
 | |
| Mean in the same column sharing the same superscript are not significantly different (p≥0.05) | |
Irrespective of dietary FM replacement levels by SBM, Nile tilapia fed diets supplemented with different phytase levels showed the highest values of P intake, P body gain, P total dissolved waste, P vertebral column and Ca vertebral column associated with lowest (p≥0.05) values of P total waste discharge and P total solid waste compared with fish fed control diet (Table 5).
Regarding to dietary phytase supplemented, tilapia fed the diet with 1500 FTU kg-1 phytase showed the highest values of P intake, P body gain and P vertebral column. The highest value of P total dissolved waste was shown for fish fed phytase level of 2000 FTU kg-1 diet associated with the lowest value of P total solid waste
DISCUSSION
In the present study, over the 13 week feeding period all water quality guidelines values were found within the acceptable ranges recommended by Papoutsoglou and Tziah (1996) for tilapia culture and indicated no pose of any stress factors to the fish during the experiment.
The total phosphorus in all experimental test diets (Table 1) was adequate to cover Nile tilapia phosphors P requirement (0.81 g 100 g-1 diet) according to Miranda et al. (2000), but dietary non-phytate P was kept in all diets below the P recommendations to detect the possible phytase supplementation action in increase dietary available P generated compared with control diet. Therefore, differences in growth and other performance indices can be referred to either variation of their dietary protein sources or their phytase supplementation levels. However, the results showed that a 25% substitution of dietary FM with SBM in Nile tilapia fingerlings diets has enhance growth performance and feed efficiency. With increasing dietary SBM levels; comparably lower growth indices were obtained compared to 100%-FM control diet. These results agreed with the findings of Riche and Garling (2004) who showed a linear decrease in Nile tilapia weight gain, relative to the control diet (FM-based diet), with dietary SBM beyond 250 g kg-1 CP. Dabrowski et al. (1989) attributed the increase of growth performance at lower substitution (250 g kg-1 CP) levels of plant protein in tilapia diets due to improved essential amino acid patterns with plant proteins incorporated. Riche et al. (2001) stated that inclusion of high levels of SBM can exacerbate problems not only related to the essential amino acid balance but also with regarded to the possible effects of anti-nutritional factors. Generally, tilapia has been shown to respond favorably to the dietary replacement of FM with SBM. Many studies have shown considerable success in partially replacing FM with SBM and other soybean products in the diets of various tilapia species (El-Sayed and Tacon, 1997). Other studies showed that higher inclusion levels of SBM reduce growth performance and feed utilization (Riche et al., 2001; Riche and Garling, 2004). Meanwhile, some authors reported that total replacement is possible (Viola et al., 1988: Shiau et al., 1990; El-Saidy and Gaber, 1997, 2002; Goda et al., 2007). These conflicting results may be due to the variation of environmental conditions (e.g., controlled laboratory conditions, vs. pond culture conditions), water quality (e.g., green water ponds vs. clear water in glass aquaria or fiberglass tanks); feed processing (pelleted vs. extruded) and to differences in the nutritional quality of the SBM used within the experimental test diets (El-Sayed, 2006; Amaya et al., 2007). The dietary SBM inclusion level depending upon a variety of different factors, including fish species and size, SBM source and processing method, aqua-feed processing, manufacturing method and culture system employed. One of the major factors that influenced the dietary inclusion of SBM in tilapia feeds is the dietary protein level. Shiau et al. (1987) reported that for O. niloticus X O. aureus hybrids FM could be partially replaced by SBM within diets containing sub-optimal protein levels (24%), whereas at optimum protein levels (32%) the dietary replacement of FM with 30% SBM significantly depressed fish performance. In another series of feeding trials with hybrid tilapia Viola et al. (1988) evaluated 30% CP diets containing 55-60% SBM without animal feedstuffs along with various supplements such as oil, methionine, lysine and phosphorus. Phosphorus supplementation of SBM based-diet was required to achieve weight gain and feed efficiency responses similar to that of fish feed a control diet containing 35% FM. The growth results in the present study for Nile tilapia after being fed different SBM-based diets, clearly indicate that with increasing dietary levels of SBM up to 100% CP, a number of nutrition P-deficiency symptoms were observed, such as relatively inferior WG and SGR that were consequently associated with the decrease of available of P which was considered insufficient for optimal growth of tilapia. Therefore, the total P as phytate in different experimental diets (Table 1) that ranged from 0.26-0.52% appeared to cause the lower growth indices recorded in Nile tilapia. Goda et al. (2002) reported that Nile tilapia fed either SBM or full-fat soybean meal basal-diets supplemented with dicalcium phosphate to level of P requirement, showed the highest significant (p<0.05) values in FBW and FCR compared with the control diet (100%-FM based-diet with no P supplementation). The same tendency was observed with common carp Cyprinus carpio (Schäfer et al., 1995) and Nile tilapia (Dato-Cajegas and Yakupitiyage, 1996).
The present results showed that the dietary phytase supplementation with 1000 FTU kg-1 level for Nile tilapia fingerlings was adequate to improve growth performance and nutrient utilization. The trend of these results suggested that more nutrients in the soybean meal were liberated with these levels of phytase addition, which may lead to significant improvement in growth rate. Yan et al. (2002) reported that dephosphorylation of phytate in the stomach of channel catfish (Ictalurus punctatus) increased with the level of phytase supplementation. This further explains that nutrient bioavailability and liberation in plant feed ingredients depend largely on the levels of phytase inclusion.
The highest significant (p≤0.05) value of feed conversion ratio (FCR) was showed when Nile tilapia fed 75%-SBM diet (1.89); while fish fed either 25%-SBM or 100%-FM diets recorded the better significant (p≤0.05) FCR values (1.66 and 1.67, respectively). The increase FCR values with increasing dietary SBM levels in the present study indicate that the plant protein-based diets were not suggested to be high performance diets for Nile tilapia. The highest levels of plant ingredients in the experimental diets result in higher content of insoluble carbohydrates (such as cellulose) as well as elevated levels of indigestible phytates, both of which are known to depress performance in fish (NRC, 1993; Cain and Garling, 1995; Riche and Brown, 1996; Francis et al., 2001; Portz and Liebert, 2004). However, the highest significant (p≤0.05) feed intake (190.60 g kg-1 90 days-1) was observed for Nile tilapia fed diet supplemented with phytase at a level of 1000 FTU kg-1 diet. There is some evidence to indicate that dietary phytase supplementation has increase feed intake of fish fed diets with P at sub-optimal or requirement levels. In this context, the present data agreed with the findings of Rodehutscord and Pfeffer (1995) that showed increased feed intake in rainbow trout (Oncorhynchus mykiss) fed a diet containing phytase at a sub-optimal P level. They concluded that increased availability of P due to phytase was the reason for increased feed intake and resulted in higher growth performance. Higher feed intake was observed in catfish Ictalurus punctatus fed diets containing phytase at least 250 FTU kg-1 (Li and Robinson, 1997) and at least 500 FTU kg-1 phytase (Jackson et al., 1996). Similarly, Hauler and Carter (1997) showed that phytase in a plant protein based-diet with a slightly deficient level of available phosphorus (5 g kg-1) increased feed intake in Atlantic salmon, Salmo salar.
As regards to proximate composition of final whole fish (Table 3), these results agreed with the findings of Abdelghany (2003) who reported that substituting of dietary herring FM with soybean flour (0, 25, 50, 75 and 100%) had no affect on final body fish protein, ash and moisture contents of red tilapia Oreochromis niloticus x O. mossambicus.
The present results indicated that Nile tilapia had a similar or slightly higher apparent digestibility coefficient (ADC) of crude protein with increase dietary substitution of FM with SBM. These data agreed with the findings of Shiau et al. (1987, 1989). However, irrespective of dietary FM replacement levels, these data indicated that phytase supplementation to the experimental diets had improved the ADC of protein, gross energy and lipid. The same trend was observed for ADC of phosphorus. The improvement in the ADC of protein and gross energy agreed with the findings of Portz and Liebert (2004) who reported similar improvement mainly for diets with 1000 and 2000 FTU kg-1 phytase supplementation. They added that the apparent P digestibility and utilization was increased significantly (p≥0.05) for diet supplemented with 4000 FTU kg-1. However, the inclusion rate of phytase in present study may plays an important role in releasing phytate-phosphorus in SBM-based diets. Phytase supplementation in diets for many fish species have previously been shown to be effective in releasing P (Cain and Garling, 1995; Rodehutscord and Pfeffer, 1995; Storebakken et al., 1998; Van Weerd et al., 1999; Vielma et al., 1998, 2000; Papatryphon et al., 1999; Cheng and Hardy, 2004; Baruah et al., 2007). Generally, the effect of phytase on digestibility may depend on a variety of dietary factors such as source and concentration of phytate in the diet, source and concentration of protein in the diet (Selle et al., 2000); digestibility of protein source, calcium and phosphorus levels (Sugiura et al., 2001). Furthermore, methods used for adding phytase during diet manufacture, such as ingredient pretreatment or direct supplementation of the diet with phytase may also have effects on fish growth performance and feed utilization efficiency.
A possible explanation for reduced availability of nutrients from diets containing phytase treated in the present study (at the level of 2000 FTU kg-1) is Nile tilapia feeding behavior. Riche and Garling (2004) reported that tilapias are preferential grazers with palatine teeth and pharyngeal gill rankers to grind and filter material. This process creates small particles with high surface to volume ratio increasing the potential for leaching of soluble components (Hanley, 1987). Phytic acid decreases solubility of nutrients (Reddy et al., 1989) and its removal may enhance leaching. Another possibility is reduced energy availability. Tilapia amylase activity is high for fish allowing them to utilize carbohydrates better than other fish species (Anderson et al., 1991). A reduction of amylase activity may have decreased available energy. Alternatively, phytic acid may serve in a protective capacity. Phytic acid binds to the globular proteins glycinin and β-conglycinin (Reddy et al., 1989), both identified as ANFs in fish (Kaushik et al., 1995; Rumsey et al., 1995). Additionally, phytic acid may protect sensitive AA, such as Lys, from degradation during processing or pelleting.
The data in Table 5 observed that, with increasing dietary phytase levels; comparably lower phosphorus effluent was showed compared to control diet. These results suggested that phytase addition in diets could reduce mineral excretion into the culture environment which invariably means more mineral deposition in the tissues of the fish. The minimization of the minerals effluent into the environment would reduce environmental pollution associated with fish culture effluents. Sugiura et al. (2001) reported that excretion of P in the faeces of fish fed a low-ash diet containing phytase-treated SBM was reduced by 95-98% compared with P excretion by fish that consumed commercial diet without phytase. These ascertain supports the observation from the present study that the total waste of P effluent and P total solid waste were significantly reduced by phytase treatment of the different experimental diets. de la Higuera (2001) reported that when fish are fed diets deficient in a nutrient, two opposite responses are possible: when the deficiency is mild, increasing feed intake will compensate by increasing intake of the nutrient but when the deficiency is severe, an inhibitory response is observed. Decreased feed intake in fish fed severely deficient diets may prevent or delay the onset of metabolic disorder. The present results indicated that the inclusion of SBM in the diets led to a decreased feed intake in Nile tilapia (sub-optimal phosphorus case) (Table 2) and P intake (Table 5), thus the increased available phosphorous due to phytase was stimulated the maximum feed intake, resulted in a large increase in feed intake and therefore growth. The same trend was reported by Rodehutscord and Pfeffer (1995) for rainbow trout, (Oncorhynchus mykiss).
In conclusion, the results of the present study suggested that a small substitution of dietary FM with SBM in Nile tilapia diets has resulted in higher growth rate and feed utilization efficiency. However inclusion of high levels of SBM in the diets can exacerbate problems not only related to the essential amino acid balance but also with regard to the possible effects of ant-nutritional factor, especially phytates. Therefore, addition of phytase to different diets can improve fish growth rate, feed conversion and feed digestibility that tend to confer superiority for the diet supplemented with 1000 FTU kg-1 of phytase. Nevertheless, the supplementation of microbial phytase has demonstrated to be efficient for increasing dietary phosphorus supply for Nile tilapia fingerlings, thereby reducing the phosphorus effluent from aquaculture facilities. This may have to be taken into consideration for ongoing feeding strategies for the control of waste discharge.
ACKNOWLEDGMENTS
The author would like to express his sincerest gratitude to Prof. Dr. Elham A. Wassef, Fish Nutrition Laboratory, National Institute of Oceanography and Fisheries (NIOF), Alexandria, Egypt, for her help. The author would like to thank Dr. Ehab R. El Haroun, Animal Production Department, Faculty of Agriculture, Cairo University, Egypt and Dr. Mohamed A. S. El-Sherbiny, Food Technology Center Laboratory, Agriculture Research Center, Ministry of Agriculture, Egypt for Financial support of the digestion trails, providing the microbial phytase and Laboratory analyses of in-feed phosphors and phytase activity.
REFERENCES
- Abdelghany, A.E., 2003. Replacement of herring fish meal by soybean flour in practical diets for red tilapia, Oreochromis niloticus x O. mossambicus, grown in concrete tanks. J. Applied Aquacult., 14: 69-87.
CrossRefDirect Link - Amaya, E.A., D.A. Davis and D.B. Rouse, 2007. Replacement of fish meal in practical diets for the pacific white shrimp (Litopenaeus vannamei) reared under pond conditions. Aquaculture, 262: 393-401.
Direct Link - Anderson, J., B.S. Capper and N.R. Bromage, 1991. Measurement and predication of digestible energy values in feedstuffs for the herbivorous fish tilapia (Oreochromis niloticus Linn.). Br. J. Nutr., 66: 37-48.
Direct Link - Baruah, K., N.P. Sahu, A.K. Pal and D. Debnath, 2004. Dietary Phytase: An ideal approach for a cost effective and low-polluting aquafeed. NAGA, World Fish Center Q., 27: 15-19.
Direct Link - Baruah, K., N.P. Sahu, A.K. Pal, K.K. Jain, D. Debnath and S.C. Mukherjee, 2007. Dietary microbial phytase and citric acid synergistically enhances nutrient digestibility and growth performance of Labeo rohita (Hamilton) juveniles at sub-optimal protein level. Aquacult. Res., 38: 109-120.
CrossRefDirect Link - Brett, J.R., 1973. Energy expenditure of Sockeye salmon Oncorhynchus nerka, during sustained performance. J. Fish. Res. Board Can., 30: 1799-1809.
CrossRefDirect Link - Bureau, D.P. and C.Y. Cho, 1999. Phosphorus utilization by rainbow trout (Oncorhynchus mykiss): Estimation of dissolved phosphorus waste output. Aquaculture, 179: 127-140.
Direct Link - Cain, K.D. and D.L. Garling, 1995. Pretreatment of soybean-meal with phytase for salmonid diets to reduce phosphorus concentrations in hatchery effluents. Progr. Fish-Cult., 57: 114-119.
Direct Link - Caldwell, R.A., 1992. Effect of calcium and phytic acid on the activation of trypsinogen and the stability of trypsin. J. Agric. Chem., 40: 43-46.
CrossRefDirect Link - Cho, C.Y., J. Hynes, K. Wood and H. Yoshida, 1994. Development of high nutrient-dense, low pollution diets and prediction of aquaculture wastes using biological approaches. Aquaculture, 124: 293-305.
Direct Link - Dabrowski, K., P. Poczyczynski, G. Kock and B. Berger, 1989. Effect of partially or totally replacing fish meal protein by soybean meal protein on growth, food utilization and proteolytic enzyme activities in rainbow trout (Salmo gairdneri). New in vivo test for exocrine pancreatic secretion. Aquaculture, 77: 29-49.
- De la Higuera, M., 2001. Effects of Nutritional Factors and Feed Characteristics on Feed Intake. In: Feed Intake in Fish, Houlihan, D., T. Boujard and M. Jobling (Eds.). Blackwell Science, Oxford, pp: 250.
CrossRef - Dato-Cajegas, C.R.S. and A. Yakupitiyage, 1996. The needs for dietary mineral supplementation for Nile tilapia Oreochromis niloticus cultured in a semi-intensive system. Aquaculture, 144: 227-237.
Direct Link - Eeckhout, W. and M. De Paepe, 1994. Total phosphorus, phytate-phosphorus and phytase activity in plant feedstuffs. Anim. Feed Sci. Technol., 47: 19-29.
CrossRefDirect Link - El-Dahhar, A.A. and K. El-Shazly, 1993. Effect of essential amino acids (methionine and lysine) and treated oil in fish diet on growth performance and feed utilization of Nile tilapia, Tilapia nilotica (L.). Aquacult. Fish. Manage., 24: 731-739.
Direct Link - El-Saidy, D.M.S.D. and M.M.A. Gaber, 2002. Complete replacement of fish meal by soybean meal with dietary L-lysine supplementation for Nile tilapia Oreochromis niloticus (L.) fingerlings. J. World Aquacult. Soc., 33: 297-306.
CrossRefDirect Link - El-Sayed, A.F.M., 2006. Tilapia Culture. CAB International, Wallingford, England, ISBN: 9781845930165, Pages: 277.
Direct Link - Engelen A.J., F.C. van der Heeft, P.H.G. Randsdorp and E.L.C. Smit, 1994. Simple and rapid determination of phytase activity. J. AOAC. Int., 77: 760-764.
Direct Link - Forster, I., D.A. Higgs, B.S. Dosanjh, M. Rowshandeli and J. Parr, 1999. Potential for dietary phytase to improve the nutritive value of canola protein concentrate and decrease phosphorus output in rainbow trout (Oncorhynchus mykiss) held in 11°C fresh water. Aquaculture, 179: 109-125.
CrossRefDirect Link - Francis, G., H.P.S. Makkar and K. Becker, 2001. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture, 199: 197-227.
CrossRefDirect Link - Farukawa, A. and H. Tsukahara, 1966. On the acid digestion method for the determination of chromic oxide as an index substance in the study of digestibility of fish feed. Bull. Jpn. Soc. Sci. Fish., 32: 502-506.
CrossRefDirect Link - Goda, A.M., E.R. El-Haroun and M.A.K. Chowdhury, 2007. Effect of totally or partially replacing fish meal by alternative protein sources on growth of African catfish Clarias gariepinus (Burchell, 1822) reared in concrete tanks. Aquacult. Res., 38: 279-287.
CrossRefDirect Link - Jackson, L.S., M.H. Li and E.H. Robinson, 1996. Use of microbial phytase in channel catfish Ictalurus punctatus diets to improve utilization of phytate phosphorus. World Aquacult. Soc., 27: 309-313.
CrossRefDirect Link - Kaushik, S.J., J.P. Cravedi, J.P. Lalles, J. Sumpter, B. Fauconneau and M. Laroche, 1995. Partial or total replacement of fish meal by soybean protein on growth, protein utilization, potential estrogenic or antigenic effects, cholesterolemia and flesh quality in rainbow trout, Oncorhynchus mykiss. Aquaculture, 133: 257-274.
Direct Link - Li, M.H. and E.H. Robinson, 1997. Microbial phytase can replace inorganic phosphorus supplements in channel catfish Ictalurus punctatus diets. J. World Aquac. Soc., 28: 402-406.
CrossRefDirect Link - Liebert, F. and L. Portz, 2005. Nutrient utilization of Nile tilapia Oreochromis niloticus fed plant based low phosphorus diets supplemented with graded levels of different sources of microbial phytase. Aquaculture, 248: 111-119.
Direct Link - Liener, I.E., 1994. Implications of antinutritional components in soya bean foods. Crit. Rev. Food Sci. Nutr., 34: 31-67.
PubMedDirect Link - Martin, E.A., J.V. Nolan, Z. Nitsan and D.J. Farrell, 1998. Strategies to improve the nutritive value of rice bran in poultry diets. IV. Effects of addition of fish meal and a microbial phytase to duckling diets on bird performance and amino acid digestibility. Br. Poult. Sci., 39: 612-621.
CrossRefPubMedDirect Link - Mroz, Z., A.W. Jongbloed and P.A. Kemme, 1994. Apparent digestibility and retention of nutrients bound to phytate complexes as influenced by microbial phytase and feeding regimen in pigs. J. Anim. Sci., 72: 126-132.
Direct Link - NRC., 1993. Nutrient Requirements of Fish. National Academy Press, Washington, DC, USA, ISBN-13: 9780309048910, Pages: 128.
Direct Link - Papatryphon, E., R.A. Howell and J.H. Jr. Soares, 1999. Growth and mineral absorption by striped bass Morone saxatilis fed a plant feedstuff based diet supplemented with phytase. J. World Aquacult. Soc., 30: 161-173.
Direct Link - Papoutsoglou, S.E. and G. Tziah, 1996. Blue tilapia (Oreochromis aureus) growth rate in relation to dissolved oxygen concentration under recirculated water conditions. Aquacul. Eng., 15: 181-192.
Direct Link - Pointillart, A., A. Fourdin and N. Fontaine, 1987. Importance of cereal phytase activity for phytate phosphorus utilization by growing pigs fed diets containing triticale or corn. J. Nutr., 113: 907-913.
PubMed - Riche, M. and P.B. Brown, 1996. Availability of phosphorus from feedstuffs fed to rainbow trout, Oncorhynchus mykiss. Aquaculture, 142: 269-282.
CrossRefDirect Link - Riche, M. and D.L. Garling Jr, 2004. Effect of phytic acid on growth and nitrogen retention in tilapia Oreochromis niloticus L. Aquacult. Nutr., 10: 389-400.
CrossRefDirect Link - Riche, M., N.L. Trottier, P.K. Ku and D.L. Garling, 2001. Apparent digestibility of crude protein and apparent availability of individual amino acids in tilapia (Oreochromis niloticus) fed phytase pretreated soybean meal diets. Fish Physiol. Biochem., 25: 181-194.
CrossRefDirect Link - Robinson, E.H., M.H. Li and B.B. Manning, 2002. Comparison of microbial phytase and dicalcium phosphate for growth and bone mineralization of pond-raised channel catfish, Ictalurus punctatus. J. Applied Aquacult., 12: 81-88.
CrossRefDirect Link - Rumsey, G.L., J.G. Endres, P.R. Bowser, K.A. Earnst-Koons, D.P. Anderson and A.K. Siwicki, 1995. Soy Protein in Diets of Rainbow Trout: Effects on Growth, Protein Absorption, Gastrointestinal Histology and Nonspecific Serologic and Immune Response. In: Nutrition and Utilization Technology in Aquaculture, Lim, C.E. and J. Sessa (Eds.). AOCS Press, Champaign, Illinois, USA., pp: 166-188.
- Satoh, S., W.E. Poe and R.P. Wilson, 1989. Effect of supplemental phytate and/or tricalcium phosphate on weight gain, feed efficiency and zinc content in vertebrae of channel catfish. Aquaculture, 80: 155-161.
CrossRefDirect Link - Schafer, A., W.M. Koppe, K.H. Meyer-Burgdorff and K.D. Gunther, 1995. Effects of a microbial phytase on the utilization of native phosphorus by carp in a diet based on soybean meal. Water Sci. Technol., 31: 149-155.
CrossRefDirect Link - Sebastian, S., S.P. Touchburn, E.R. Chavez and P.C. Lague, 1997. Apparent digestibility of protein and amino acids in broiler chickens fed a corn-soybean diet supplementation with microbial phytase. Poult. Sci., 76: 1760-1769.
PubMedDirect Link - Shiau, S.Y., J.L. Chunag and C.L. Sun, 1987. Inclusion of soybean meal in tilapia (Oreochromis niloticus x O. aureus) diets at two protein levels. Aquaculture, 65: 251-261.
CrossRefDirect Link - Shiau, S.Y., C.C. Kwok, J.Y. Hwang, C.M. Chen and S.L. Lee, 1989. Replacement of fishmeal with soybean meal in male tilapia (Oreochromis niloticus x O. aureus) fingerling diets at a suboptimal protein level. J. World Aquacult. Soc., 20: 230-235.
CrossRefDirect Link - Shiau, S., S. Lin, S. Yu, A. Lin and C. Kwok, 1990. Defatted and full-fat soybean meal as partial replacements for fish meal in tilapia (Oreochromis niloticus H O. aureus) diets at low protein level. Aquaculture, 86: 401-407.
Direct Link - Sklan, D., T. Prag and I. Lupatsch, 2004. Apparent digestibility coefficients of feed ingredients and their prediction in diets for tilapia Oreochromis niloticus H Oreochromis aureus (Teleostei, Cichlidae). Aquact. Res., 35: 358-364.
Direct Link - Storebakken, T., K.D. Shearer and A.J. Roem, 1998. Availability of protein, phosphorus and other elements in fish meal, soy-protein concentrate and phytase-treated soy-protein-concentrate-based diets to Atlantic salmon, Salmo salar. Aquaculture, 161: 365-379.
CrossRefDirect Link - Sugiura, S.H., J. Gabaudan, F.M. Dong and R.W. Hardy, 2001. Dietary microbial phytase supplementation and the utilization of phosphorus, trace minerals and protein by rainbow trout [Oncorhynchus mykiss (Walbaum)] fed soybean meal-based diets. Aquacult. Res., 32: 583-592.
CrossRefDirect Link - Tacon, A.G.J., 1997. Fish meal replacers: Review of antinutrients within oilseeds and pulses-a limiting factor for the aquafeed green revolution? Proceedings of the Workshop of the CIHEAM Network on Technology of Aquaculture in the Mediterranean, Volume 22, June 24-26, 1997, CIHEAM, Apodo, Spain, pp: 153-182.
- Vaintraub, I.A. and V.P. Bulmaga, 1991. Effect of phytate on the in vitro activity of digestive proteinases. J. Agric. Food Chem., 39: 859-861.
Direct Link - Van Weerd, J.H., Khalaf and T. Aartsen, 1999. Balance trials with African catfish Clarias gariepinus fed phytase-treated soybean meal-based diets. Aquacult. Nutr., 5: 135-142.
CrossRefDirect Link - Vielma, J., S.P. Lall, J. Koskela, J. Schoner and P. Mattila, 1998. Effects of dietary phytase and cholecalciferol on phosphorus bioavailability in rainbow trout (Oncorhynchus mykiss). Aquaculture, 163: 309-323.
CrossRefDirect Link - Vielma, J., T. Makinen, P. Ekholm and J. Koskela, 2000. Influence of dietary soy and phytase levels on performance and body composition of large rainbow trout (Oncorhynchus mykiss) and algal availability of phosphorus load. Aquaculture, 183: 349-362.
CrossRefDirect Link - Vielma, J., K. Ruohonen and M. Peisker, 2002. Dephytinization of two soy proteins increases phosphorus and protein utilization by rainbow trout, Oncorhynchus mykiss. Aquaculture, 204: 145-156.
CrossRefDirect Link - Viola, S., Y. Arieli and G. Zohar, 1988. Animal-protein-free feeds for hybrid tilapia (Oreochromis niloticus x O. aureus) in intensive culture. Aquaculture, 75: 115-125.
CrossRefDirect Link - Wassef, E.A., G.H. Palmer and M.G. Poxton, 1988. Protease digestion of the meals of ungerminated and germinated soya beans. J. Agric. Food Chem., 44: 201-214.
CrossRefDirect Link - Wheeler, E.L. and R.E. Ferrel, 1971. A method of phytic acid determination in wheat and wheat fractions. Cereal Chem., 48: 312-320.
Direct Link - Yan, W., R.C. Reigh and Z. Xu, 2002. Effects of fungal phytase on utilization of dietary protein and minerals and dephosphorylation of phytic acid in the alimentary tract of channel catfish Ictalurus punctatus fed an all-plant-protein diet. J. World Aquac. Soc., 33: 10-22.
CrossRefDirect Link - Yoo, G.Y., X. Wang, S. Choi, K. Han, J.C. Kang and S.C. Bai, 2005. Dietary microbial phytase increased the phosphorus digestibility in juvenile Korean rockfish Sebastes schlegeli fed diets containing soybean meal. Aquaculture, 243: 315-322.
CrossRefDirect Link