
H.I. El- Marakby
Fish Nutrition Department, Central Laboratory for Aquaculture Research, Sharqia, Egypt
A.M. Eid
Department of Animal Production, Faculty of Agriculture, Suez Canal University, Egypt
A.E. Abdelghany
Fish Nutrition Department, Central Laboratory for Aquaculture Research, Sharqia, Egypt
M. Abdel -Tawwab
Fish Ecology Department, Central Laboratory for Aquaculture Research, Egypt
Journal of Fisheries and Aquatic Science
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 87-96







ABSTRACT
This study was carried out to investigate the ability of striped mullet (Mugil cephalus) to select phytoplankton at different feeding regimes. Striped mullet (35.1±1 g) was stocked at a rate of 1 fish/m2 for 14 weeks. The first treatment (control) received only organic and inorganic fertilizers. The second treatment (T1) received organic and inorganic fertilizers as in the first treatment and supplemental feed at a rate of 3% of fish body weight (bw). The third treatment (T2) received only supplemental feed at a rate of 5% of fish bw. Each treatment was represented by three replicates. The maximum final weight was obtained at T1 and T2 without significant differences, while the minimum one was obtained at control treatment. The fresh weight of stomach content in control treatment was the highest one. Phytoplankton occurrence in gut content of striped mullet was higher in control sharing 70-80% of total gut content, while it was significantly lower in T2 (p<0.05). Detritus consisted mainly of scraps of macrophytes, organic particles including feed particles and mud. The composition percentage of detritus in fish stomach was significantly differed and ranked as T2>T1>control. Zooplankton sharing to gut content was generally small in T2 and the highest content was obtained at control. Whenever zooplankton occurred, it was not exceed 10.0% of the total components in fish stomach, and it consisted of parts of animals especially cladocera, copepoda and rotifers. Sometimes, parts of insects and zoobenthos were found. The main phytoplankton genera found in fish stomach are belonged to Cyanobacteria, Chlorophyceae, Bacillariophyceae and Euglenophyceae. Bacillariophyceae represented the main phytoplankton division, while Cyanobacteria followed by Chlorophyceae were the less abundant divisions at all treatments. Euglenophyceae represented moderate abundance in fish stomach content. Striped mullet selected Bacillariophyceae and Euglenophyceae at all treatments, meanwhile Cyanobacteria was sometimes selected at T1 and T2. Chlorophyceae was not selected at all and it was gut entered with water and mechanically swallowed together with other foodstuffs. This result indicates that striped mullet naturally does not consume food at random but have the ability to select and choose the preferred foodstuff and could use supplemental feed efficiently.
PDF Abstract XML References
How to cite this article
H.I. El- Marakby, A.M. Eid, A.E. Abdelghany and M. Abdel -Tawwab, 2006. The Impact of Striped Mullet, Mugil cephalus on Natural Food and Phytoplankton Selectivity at Different Feeding Regimes in Earthen Fishponds. Journal of Fisheries and Aquatic Science, 1: 87-96.
DOI: 10.3923/jfas.2006.87.96
URL: https://scialert.net/abstract/?doi=jfas.2006.87.96
DOI: 10.3923/jfas.2006.87.96
URL: https://scialert.net/abstract/?doi=jfas.2006.87.96
INTRODUCTION
Organic and/or inorganic fertilizers are commonly used in pond fertilization and artificial feeds are also applied. Many investigations have shown positive correlation between pond fertilization and fish production (Boyd, 1984; Ahmad et al., 2001; Abdel-Tawwab et al., 2002; Ibrahim et al., 2003). The increase in fish productivity in fertilized ponds has been attributed to an increase in primary productivity that in turn increases zooplankton production, thereby producing greater yields of fish.
Striped mullet (Mugil cephalus) is one of the best known members of Mugilidae and has commercial value in many countries (Oren 1981). It is an euryhaline marine teleost that inhabits coastal waters but enters estuaries and rivers where it feeds on microalgae, filamentous algae, diatoms, detritus associated with sand and mud (Harrison and Senou 1997) and it is widely cultured in both brackish and freshwater semi-intensive fishponds as a secondary species to improve sediment quality (Sarig, 1981). As adults they eat all available food, microalgae including epiphytic and benthic forms and decaying plant detritus. They feed at the lowest trophic level, being able to utilize the direct grazing or plant detritus food chains as energy sources (Odum, 1970; Brusle, 1981). Porter et al. (1996) indicated that grey mullets are able to reduce the organic matter in the sediment underneath the cages. Thus, Katz et al. (2002) and Lupatsch et al. (2003) used striped mullet (Mugil cephalus) to remove the organic matter deposited on the sea floor due to the deposition of solid waste from undigested feed.
In Abbassa fishponds, striped mullet is one of the most important components of fish polyculturing system. Recently, fish intensification is one of the most important goals in Egypt throughout ponds or cages culture. To intensify striped mullet, it should be understanding its food habit and to which extent it could use artificial feed to maximize its growth. Therefore, this work was carried out to study the food habit of striped mullet (M. cephalus) and the contribution of phytoplankton to the natural food throughout the investigation of stomach contents in fishponds received either organic and inorganic fertilizers and/or artificial feed.
MATERIALS AND METHODS
Earthen ponds (surface area 1000 m2 each) at Central Laboratory for Aquaculture Research, Abbassa, Sharqia were used in this study. The ponds had been drained, cleaned and refilled with new freshwater from El-Wadi canal derived from El-Ismailia canal. The water level was adjusted at 1.0 m depth. Before experiment starting, six ponds were fertilized with cow manure at a rate of 300 kg/acre, which distributed at different places on the pond’s bottom. Fish was collected from mullet nursery pond and the average initial weight was 35.1±1 g. Fish stocked in the experimental ponds at a rate of 1 fish/m2 on September 9th and reared for 14 weeks. Chicken manure had been applied in fertilized ponds at a rate of 10 kg/acre at biweekly intervals. Inorganic fertilizers of urea (46.5% N) and superphosphate (15.5% P2O5) were also applied biweekly at a rate of 1 and 5 kg/acre, respectively. The inorganic fertilizers were dissolved and broadcasted on the water surface of fishponds.
Three treatments with three replications were assigned to evaluate the acceptability of striped mullet for natural food and/or supplemental feed. The first three ponds received only organic and inorganic fertilizers (control), the second three ponds (T1) received organic and inorganic fertilizers as in control ponds and supplementary feed at a rate of 3% of fish body weight as well and the third three ponds (T2) received only supplementary feed at a rate of 5% of fish body weight. The used diet contained 35% crude protein, 9.6% total lipids, 4.6% crude fiber, 15.5% ash and 407.7 kcal/100 g gross energy. The feed was provided to fed-ponds twice daily.
Every two weeks, 25 fish from each pond were sampled and individual length and weight were measured, then 20 fish were returned to the pond, meanwhile 5 fish were captured and immediately placed in 10% formalin. According to the weight of fish samples, the amounts of given feed, in fed ponds, were readjusted. Fish were dissected and the length of digestive tract was measured and the ratio between length of digestive tract to total fish length (L ratio) was calculated. The stomach fullness degree was estimated and fullness degree of fish stomachs was categorized to empty, ¼ full, ½ full, ¾ full, full and gorged as described by Abdelghany (1993). The stomach contents was freshly weighed and transferred to a fixed volume of distilled water. Three 1-mL subsamples from each stomach were then transferred to Sedgwick-Rafter counting cell and phytoplankton was identified and counted. The phytoplankton in samples was identified to genus according to Prescott (1961). The composition of the phytoplankton community in the stomach was then compared with that of fishpond to evaluate grazing selectivity, which was calculated according to Ivlev (1961).
Water samples for chemical analyses were collected weekly between 08:30 and 09:30 h at 30 cm depth from each pond. Dissolved oxygen and temperature were daily monitored in site at 30 cm depth with a YSI model 58 oxygen meter (Yellow Spring Instrument Co., Yellow Springs, Ohio, USA). Water conductivity was measured with a YSI model 33 conductivity meter (Yellow Spring Instrument Co., Yellow Springs, Ohio, USA). The pH value and free ammonia were measured colorimetrically by using Hach kits (Hach Co., Loveland, Colorado, USA). Total alkalinity and total hardness was measured by titration. Orthophosphate was colorimetrically determined by stannous chloride method and nitrate was colorimetrically determined by phenoldisulphonic acid method (Boyd, 1984).
In the same time, water samples (1-liter bottle) were collected for phytoplankton determination at the same depth (30 cm) and preserved by adding 1 mL of 4% formalin. Samples were allowed to settle for 15 days and the supernatant was siphoned to 50 mL. Water samples for zooplankton determination was collected by plankton net (50 μm) throughout filtering 20 L and soonly preserved by adding 1 mL of 10% formalin. The counts of phytoplankton and zooplankton were performed using Sedgwick-Rafter cell under a binocular microscope using suitable magnification.
The obtained data were subjected to Kolmogrov-Smirnov and Cochran’s tests for normality and homogeneity, respectively. Some data did not show homogeneity of variance. So, the data sets were analyzed among treatment levels with Kruskal-Wallis one-way analysis of variance (ANOVA). Mean separations were determined using Tukey’s Test to determine which treatment had greater values. Statistical analyses were conducted by using SPSS software program version 10 (SPSS, Richmond, USA) as described by Dytham (1999).
Results
The variations of the investigated parameters of water quality are presented in (Table 1). Water temperature showed remarkable decreasing where maximum value was recorded in September (28.7°C) and gradually decreased to reach the lowest one in December (12.30°C; Fig. 1). Turbidity, dissolved oxygen, nitrate and phosphate concentrations were significantly affected by feeding regimes (Table 1). Water conductivity, pH value, free ammonia, total alkalinity and total hardness were insignificantly affected by feeding regimes of striped mullet in this study (p>0.05).
Phytoplankton occurrence was positively affected by feeding regimes and it was significantly higher at control and T1 than that of T2 (Table 1; p<0.05). The most abundant genera of Bacillariophyceae were Navicula, Nitzschia and Synedra. Among Chlorophyceae, Pediastrum, Scenedesmus and Tetraedron were found to be the most abundant genera.
| Table 1: | Changes in physico-chemical and biological parameters (mean±SE and range in parenthesis) in earthen fishponds cultured with striped mullet at different feeding regimes |
 | |
| The same letter (s) in the same row is not significantly different at p<0.05. Control: ponds received only organic and inorganic fertilizers, T1: ponds received organic and inorganic fertilizers and 3% artificial feed and T2: ponds received 5% artificial feed only | |
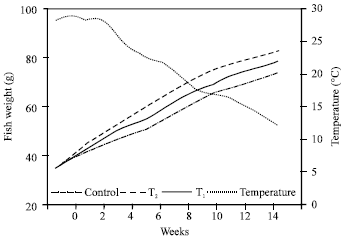 | |
| Fig. 1: | Changes in water temperature (oC) and the weight of striped mullet (g) reared in only fertilized earthen ponds (control), earthen ponds received both of fertilizers and supplemental feed at a rate of 3% BW (T1) and earthen ponds received supplemental feed only at a rate of 5% BW (T2) |
Among Cyanobacteria, Anabaena and Merismopedia were the most dominant genera. Euglenophyceae was only represented by the genera Euglena and Phacus in this investigation.
Zooplankton abundance was positively affected by feeding regime and it was greater at control and T1 than that of T2 (Table 1; p<0.05). The dominant cladocera included Moina, Bosmania, Diaphanosoma and Daphnia. Cyclops was the most common copepod and common rotifers included Brachionus and Keratella.
The growth of striped mullet was gradually increased up to the end of the experimental period (Fig. 1). The maximum final weight was obtained at T1 and T2 without significant differences (78.5 and 83.2 g, respectively; p>0.05), while the minimum one was obtained at control treatments (74.0 g). Figure 2 indicated that the average fresh weight of stomach content in control treatment was the highest one, while the lowest ones were obtained at T1 and T2 treatments with insignificant differences (p>0.05) (Fig. 2).
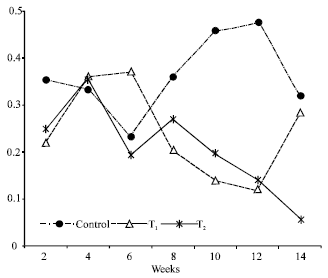 | |
| Fig. 2: | Average fresh weight of stomach contents (g/stomach) of striped mullet reared in only fertilized earthen ponds (control), earthen ponds received both of fertilizers and supplemental feed (T1) and earthen ponds received supplemental feed only (T2) |
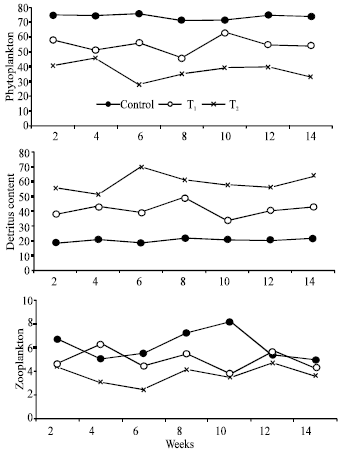 | |
| Fig. 3: | Stomach contents (%) of striped mullet reared in only fertilized earthen ponds (Control), earthen ponds received combination of fertilizers and supplemental feed (T1) and earthen ponds received supplemental feed only (T2) |
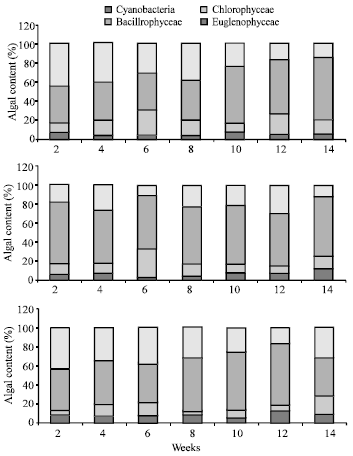 | |
| Fig. 4: | Phytoplankton contents (%) in stomach of striped mullet reared in only fertilized earthen ponds (control), earthen ponds received combination of fertilizers and supplemental feed (T1) and earthen ponds received supplemental feed only (T2) |
Considering the fluctuation of phytoplankton abundance in fish stomach, Fig. 3 shows that phytoplankton content in gut content of striped mullet was higher in control sharing 70-80% of total gut content, while it was significantly lower in T2 (p<0.05) (Fig. 3). Detritus consisted mainly of scraps of macrophytes, organic particles and mud. Feed particles were also found in the stomach of fish reared in T1 and T2 ponds. However, the composition percentage of detritus in fish stomach was significantly differed and ranked as T2>T1>control (Fig. 3; p<0.05). Zooplankton sharing to gut content was generally small in T2 and the high zoo content was obtained at control (Fig. 3; p>0.05). Whenever zooplankton occurred, it was not exceed 10.0% of the total components in fish stomach and it consisted of parts of animals especially cladocera, copepoda and rotifers. Sometimes, parts of insects and zoobenthos were found.
The main phytoplankton genera found in fish stomach are belonged to Cyanobacteria, Chlorophyceae, Bacillariophyceae and Euglenophyceae. Figure 4 shows that the abundance of phytoplankton divisions was fluctuated from treatment to another. Bacillariophyceae represented the main phytoplankton division, while Cyanobacteria followed by Chlorophyceae were the less abundant divisions at all treatments. Euglenophyceae represented moderate abundance in fish stomach content.
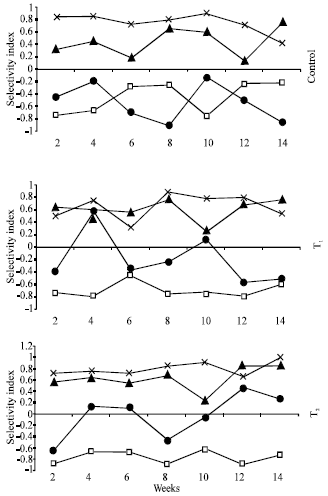 | |
| Fig. 5: | Selectivity index of striped mullet reared in only fertilized earthen ponds (control), earthen ponds received combination of fertilizers and supplemental feed (T1) and earthen ponds received supplemental feed only (T2) |
The most frequently genera represented in fish stomach in all treatments were Anabaena, Anabaenopsis, Coleospharium, Merismopedia, Microcystis, Nodularia, Oscillatoria, Spirulina (Cyanobacteria), Actinostrum, Chlorella, Closterium, Coelastrum, Eudrina, Pandorina, Pediastrum, Scenedesmus, Shereoderia, Staurastrum. (Chlorophyceae), Amphora, Cocconeis, Cymatopleura, Cymbella, Gyrosigma, Melosira, Navicula, Pinnularia, Synedra. (Bacillariophyceae) and Euglena, Phacus (Euglenophyceae).
Regarding the complex nature of the feeding habit of striped mullet in earthen fishponds with different feeding regimes, it has been necessary to calculate the selectivity index, which might throw some light on fish’s food preference. According to Ivlev’s equation (Ivlev, 1961), values of selectivity index are between +1 and –1. Positive values indicate a positive selectivity of a certain kind of food while negative ones indicate a negative selectivity. Striped mullet selected Bacillariophyceae and Euglenophyceae at all treatments, meanwhile Cyanobacteria was sometimes selected at T1 and T2 treatments (Fig. 5). Chlorophyceae was not selected at all and it was gut interred with water and mechanically swallowed together with other foodstuffs. This result indicates that striped mullet does not consume food at random but have the ability to select and choose the preferred foodstuff.
DISCUSSION
The obtained results are much as expected where feed supplementation to fishponds resulted in significantly higher growth compared to fertilization alone. However, fish growth was high at T2 and T1 due to considerable consumption of supplemental food. In this concern, Green (1992) found that growth of Nile tilapia was significantly smaller in fertilization-alone ponds than in feed-alone or combined fertilizer-feed treatments. Also, Abdelghany and Ahmad (2002) found that the growth of Nile tilapia, common carp and silver carp in combined fertilized-feed ponds were better than fertilization alone. Additionally, artificial feed may be provided to increase fish growth and production above the possible in fertilized ponds.
Brusle (1981) and Wells (1984) reported that striped mullet is a plankton feeder. Moreover, Harrison and Senou (1997) reported that striped mullet is herbivorous fish where it feeds on microalgae, filamentous algae, diatoms and detritus associated with sand and mud and it may use artificial diets. Larson and Shanks (1996) found that juvenile striped mullet (M. cephalus) actively ingested marine snow in the laboratory and assimilated organic matter and total amino acids from marine snow with efficiencies of 49.2% and 55.8%, respectively. Lupatsch et al. (2003) indicated that striped mullet could have chosen as a potential aquaculture bioremediator and it is able to reduce the organic matter in the sediments underneath the cages. Jana et al. (2004) found that striped mullet (M. cephalus) could efficiently use periphyton colonized the bamboo substrates in inland saline groundwater. In addition to the previous reviews, striped mullet in this study could efficiently use the supplemental feed with and without the natural food in freshwater earthen ponds. These conflicting results may be differed as the experimental condition, fish size and aquacultural technique differed. Moreover, growth of fish is variable with many factors like species, age (or body weight), genetic background, water temperature, diet types, feeding levels and protein sources (NRC, 1993).
In control (fertilize-alone) ponds, the growth of striped mullet was achieved throughout fish grazing and accumulation of phytoplankton nutrients. On the other side, the impact of fish on phytoplankton depends on fish filter feeding and phytoplankton growth rates as well as the predatory pressure on phytoplankton (Turker et al., 2003). In fertilize-feed ponds (T1) or feed-alone ponds (T2), striped mullet was efficiently consumed the supplemented feed.
The fluctuation in phytoplankton composition and occurrence in fish stomach resulted from the fluctuation of phytoplankton composition and occurrence in fishponds. It is clear that fish in control treatment consumed much more phytoplankton than other two treatments because the natural food is the only available food source. On the other hand, phytophagous fish consume great amounts of food and the intensity of feeding is affected by the filtration rate of food components, which depends on the density of phytoplankton in the water mass and the condition of fish.
The fish growth is slow as compared with other experimentsand the fish yield is not promising. This result may be because of the continuous decrease in water temperature (Fig. 1) although striped mullet could survive in temperature range of 7-26 oC (Harrison and Senou 1997). Certainly, the low temperature will affect the biological cycle in the pond and the physiological status of cultured fish that was reflected by high FCR. In conclusion, it is noticed that striped mullet could utilize both natural and/or supplemental feed. This result gave the possibility to intensify striped mullet in ponds or cage culture. Further work is needed concerning the quality and quantity of supplementary feed required by striped mullet at intensive fish culture.
REFERENCES
- Jana, S.N., S.K. Garg and B.C. Patra, 2004. Effect of periphyton on growth performance of grey mullet, Mugil cephalus (Linn.), in inland saline groundwater ponds. J. Applied Ichthyol., 20: 110-117.
Direct Link - Katz, T., B. Herut, A. Genin and D.L. Angel, 2002. Gray mullets ameliorate organically enriched sediments below a fish farm in the oligotrophic Gulf of Aqaba (Red Sea). Marine Ecol. Prog. Series, 234: 205-214.
Direct Link - Larson, E.T. and A.L. Shanks, 1996. Consumption of marine snow by two species of juvenile mullet and its contribution to their growth. Marine Ecol. Prog. Series, 130: 19-28.
Direct Link - Lupatsch, I., T. Katz and D.L. Angel, 2003. Assessment of the removal efficiency of fish farm effluents by grey mullets: A nutritional approach. Aqua. Res., 34: 1367-1377.
Direct Link - Turker, H., A.G. Eversole and D.E. Burne, 2003. Effect of Nile tilapia, Oreochromis niloticus (L.), size on phytoplankton filtration rate. Aqua. Res., 34: 1087-1091.
Direct Link - Wells, R.D., 1984. The food of grey mullet Mugil cephalus L. in lake Waahi and the Waikato River at Huntly, New Zealand. J. Marine Freshwater Res., 18: 13-19.
Direct Link