
Natchimuthu Karmegam
Department of Botany, Government Arts College, Salem 636 007, Tamil Nadu, India
Kuppuraj Rajasekar
Faculty of Science, Vinayaka Missions University, Salem 636 308, Tamil Nadu, India
Journal of Environmental Science and Technology
Year: 2012 | Volume: 5 | Issue: 2 | Page No.: 91-108







ABSTRACT
The present study has been conducted on the enrichment of biogas slurry vermicompost with microbial inoculants Azotobacter chroococcum and Bacillus megaterium, optimization of inoculum level and time inoculation during vermicomposting along with the survival rate of each microbial inoculant, the total microbial population and their correlation during the storage. On 0, 10, 20 and 30th day of inoculation showed decrease in the viable population of A. chroococcum and B. megaterium inoculated at the rate of 30, 35 and 40 mL-1 175 g of substrate, towards the progression of storage period of vermicompost, uniformly. The change of A. chroococcum and B. megaterium in the vermicompost (at 30, 35 and 40 mL-1 175 g substrate) with reference to the storage period (180 days) showed highly significant negative correlation (p<0.001). In all the four treatments, the viable population of A. chroococcum and B. megaterium at the rate of 30, 35 and 40 mL-1 175 g substrate from 0th day (after harvest) onwards showed statistically significant decline with that of storage period of vermicompost. This trend was observed uniformly for both the microbial inoculants, inoculated on 0, 10, 20 and 30th day of vermicomposting. In the present study, total microbial population in A. chroococcum and B. megaterium inoculated vermicompost was high during the initial phases of storage and then total microbial population declined towards the end. The vermicompost inoculated on 20th day of vermicomposting with A. chroococcum at the rate of 35 mL-1 175 g of substrate showed 11, 9, 8, 6, 3 and 0x107 cfu g-1 population of total microorganisms, respectively during 0, 15, 45, 75, 105 and 135th day of storage. Similar trend of results were obtained for B. megaterium inoculated vermicompost suggesting that the overall maintenance of total microbial population in vermicompost is similar in the vermicomposts with any microbial inoculants.
PDF Abstract XML References Citation
Received: August 20, 2011;
Accepted: December 06, 2011;
Published: January 23, 2012
How to cite this article
Natchimuthu Karmegam and Kuppuraj Rajasekar, 2012. Enrichment of Biogas Slurry Vermicompost with Azotobacter chroococcum and Bacillus megaterium. Journal of Environmental Science and Technology, 5: 91-108.
DOI: 10.3923/jest.2012.91.108
URL: https://scialert.net/abstract/?doi=jest.2012.91.108
DOI: 10.3923/jest.2012.91.108
URL: https://scialert.net/abstract/?doi=jest.2012.91.108
INTRODUCTION
The global scientific community today is searching for a technology which should be economically viable, environmentally sustainable and socially acceptable. Vermiculture technology combines all these virtues and qualities together (Sinha et al., 2010; Ansari, 2011). Vermicomposting is a controlled biological process that involves the joint action of earthworms and microorganisms to biotransform organic wastes. Vermicomposting is generally more efficient than composting because earthworms feed on organic matter and create conditions that favor the colonization and degradation of partially-decomposed and fragmented organic material by aerobic microorganisms (Edwards, 2004). Worms have been found to proliferate Actinomycetes, Azotobacter, Rhizobium, Nitrobacter and phosphate solubilizing bacteria significantly in their end products. The study conducted by Singh (2009) indicated higher values of Azotobacter (the nitrogen fixing bacteria) and the Actinomycetes (the bacteria that increase biological resistance in plants against pests and diseases) in vermicompost as compared to the conventional aerobic and anaerobic composts (Singh, 2009). Combining composting and vermicomposting also yields good results (Alidadi et al., 2007). Vermistabilization is known to bring down the level of pathogens in the raw material (Kumar and Shweta, 2011a). It is known fact that from municipal solid waste including municipal sewage sludge can be vermicomposted with earthworms (Zularisam et al., 2010; Paul et al., 2011).
Number of researchers has observed increased proliferation of a variety of microorganisms in the gut of earthworms, viz., bacteria, fungi and actinomycetes. The organic matter that passes through earthworm gut results in increased levels of microbial populations, activity and respiration and of enzymatic activity and micro and macro nutrients (Kalam et al., 2004; Prakash and Karmegam, 2010). Biofertilizers are the Plant Growth Promoting Microorganisms (PGPMs) which by several mechanisms augment plant growth (Mostafa and Abo-Baker, 2010; Abo-Baker and Mostafa, 2011; Yadav et al., 2011). On the soil, plant growth promoting microorganisms like Azospirillum, Azotobacter, Phosphobacteria and Rhizobia could contribute to solubilize and/or to acquire essential minerals, making scarce nutrients more available to the plant. Bacteria of the genus Azospirillum are associative nitrogen-fixing rhizobacteria that are found in close association with plant roots. They are able to exert beneficial effects on plant growth and yield of many agronomic crops under a variety of environmental and soil conditions. Soil bacteria of the genera Rhizobium, Azorhizobium and Bradyrhizobium (collectively referred to as rhizobia) interact with leguminous plants to form nitrogen-fixing nodules (Barassi et al., 2007; Saikia and Jain, 2007). Recent evidences on the use of biofertilizers for enriching the vermicompost showed promising results on the enhanced quality of vermicompost (Kumar and Singh, 2001; Kaushik et al., 2008).
The process of vermicomposting results in the increase of microbial diversity and activity dramatically and the vermicompost produced could be a definitive source of plant growth regulators produced by interactions between microorganisms and earthworms which could contribute significantly to increased plant growth, flowering and yields (Arancon and Edwards, 2009; Jayashree et al., 2011). So, the addition or enrichment of microbial inoculants such as biofertilizers definitely would provide an increased plant growth and yield.
The studies on the survivability of the biofertilizers, Azotobacter chroococcum and Bacillus megaterium are available (Sekar and Karmegam, 2010). However, The studies on the microbial enrichment of vermicompost with reference to the amount of inoculum required, time of inoculation, survival rate of inoculated microorganisms in vermicompost during storage and the relation of total microbial population with that of inoculated microorganisms are not documented. Hence, the present study has been undertaken to enrich biogas slurry vermicompost with the biofertilizers, A. chroococcum and B. megaterium and to optimize the biofertilizer inoculum level and time of inoculation during enrichment process of vermicompost and to assess the survival rate of A. chroococcum and B. megaterium in relation to total microbial population in the vermicompost.
MATERIALS AND METHODS
Collection of biogas slurry: Biogas slurry for the study was collected from the biogas plant situated at Kottagoundanpatty, Salem (Dt.), Tamil Nadu and used for the preparation of vermibed. One week aged biogas slurry was collected, air dried and stored in polyethylene bags until use.
Collection of earthworms: The earthworm, Eudrilus eugeniae Kinberg for the study, originally collected from culture bank of the Department of Biology, Gandhigram Rural University, Tamil Nadu, India was mass multiplied in cowdung and used for the study.
Mass multiplication of biofertilizers: The cultures of Azotobacter chroococcum (MTCC 446) and Bacillus megaterium (MTCC 453) were procured from Microbial Type Culture Collection (MTCC), Chandigarh and used for the present study. The organisms were revealed in the suggested broth medium and sub-cultured in Jensons and Nutrient Agar media, respectively. A loop full of A. chroococcum and B. megaterium was transferred, respectively to 100 mL of respective selective medium and incubated. After incubation, 10 mL of the inoculum was transferred to 1000 mL of respective broth and kept in shaking incubator for mass multiplication.
Enrichment of vermicompost with biofertilizers: For enrichment studies, four different vermicomposting trials, each with six replicates were carried out by preparing the vermibeds. In this, the vermicompost was collected from all the vermibeds after 40 days and subjected to physico-chemical and microbiological analysis as per the standard procedures. E. eugeniae was introduced in all the vermibeds. The mass multiplied biofertilizer organisms at the rate of 30, 35 and 40 mL-1 175 g of vermibed substrates were added to each of the experiments, T1, T2, T3 and T4 on 0, 10, 20 and 30th day, respectively to find out the optimum inoculum level and time of inoculation that results in the maintenance of 1x107 viable cells per gram of vermicompost during storage.
Statistical analyses: Data were subjected for Analysis of Variance (ANOVA) followed by Duncan’s multiple-range test to differentiate the significant difference between different treatments at the probability level of p<0.05 using SPSS® computer software for Windows (version 9.05). The relation between viable cell counts in different carrier materials and incubation days were carried out using Microcal Origin Computer Software (Version 6.1) and correlation coefficient (r) was calculated to know the level of significance of correlation.
RESULTS AND DISCUSSION
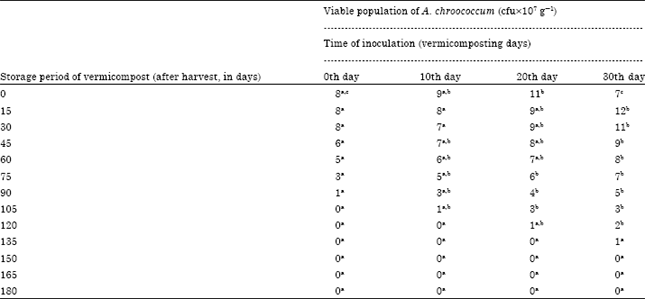
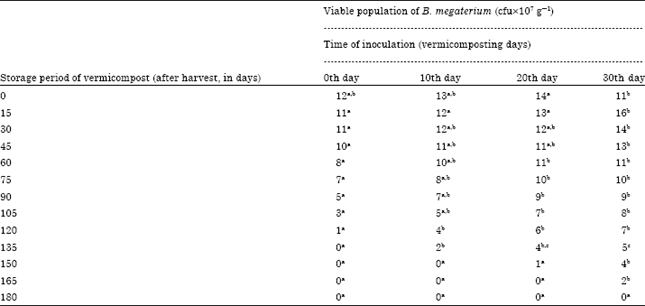
Microbial enrichment of vermicompost: Enrichment studies with A. chroococcum: A. chroococcum inoculated at the rate of 30 mL-1 175 g of substrate showed the viable cells in 10-7 dilution upto 75, 90 and 105th day of storage in 10, 20 and 30th day of inoculation, respectively (Table 1). A. chroococcum inoculated on 0, 10 and 20th day showed decline in viable population during storage period. In 30th day of inoculation, the viable population initially increased, then decreased. The viable population of 5, 9 and 7 cfu x107 g-1 were observed on 0, 15 and 30th day of storage of vermicompost which received A. chroococcum inoculum on 30th day of vermicomposting. Then, colonies in the same vermicompost showed decline upto 105 days of storage. The change of A. chroococcum population in vermicompost which received the inoculum level of 30 mL-1 175 g substrate during the storage period showed negative correlation. The 30th day of inoculation of A. chroococcum during vermicomposting showed viable population in 107 dilution for a maximum storage period i.e., 105 days. The decrease of A. chroococcum population showed significant negative correlation with storage period (p<0.001) (Fig. 1). The viable population of A. chroococcum inoculated at the rate of 35 mL-1 175 g of substrate in different intervals showed decrease in viable population during the storage of vermicompost. The 30th day inoculation of A. chroococcum showed survival upto 105 days whereas, the A. chroococcum inoculated on 10 and 20th day showed the viable population upto 90 and 105th day, respectively.
| Table 1: | The viable population of A. chroococcum inoculated at the rate of 30 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
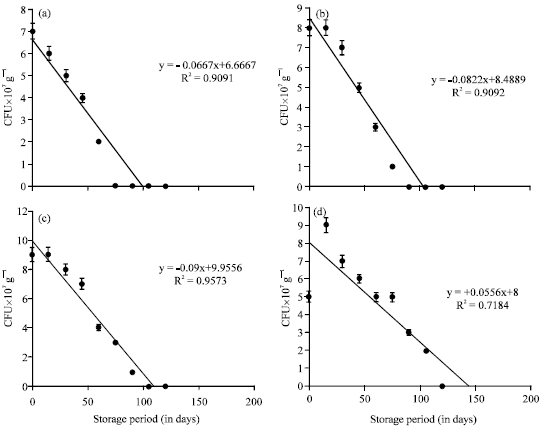 | |
| Fig. 1(a-d): | The change of A. chroococcum population in the vermicompost (at 30 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of A. chroococcum during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
| Table 2: | The viable population of A. chroococcum inoculated at the rate of 35 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
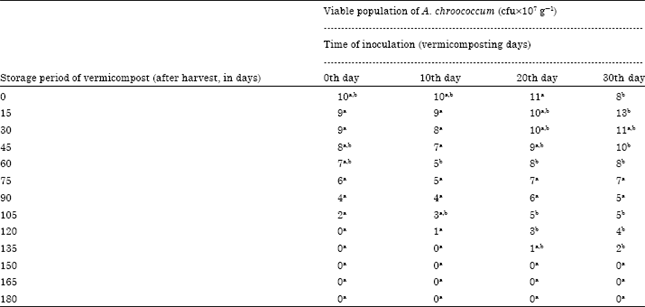
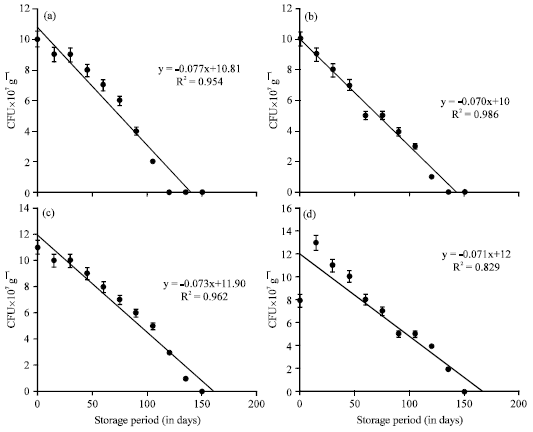
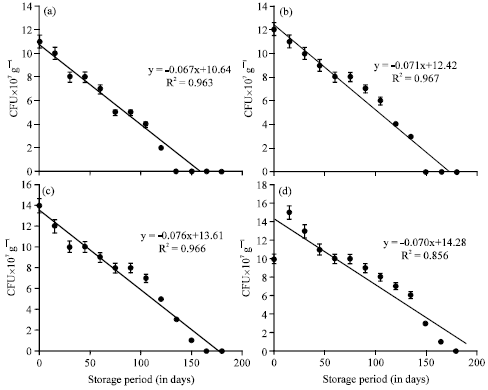
The 0th day inoculation showed viable population upto 90 days of storage period (Table 2). The decrease of viable population upon storage of vermicompost which received A. chroococcum inoculum at different intervals were negatively correlated (Fig. 2) A. chroococcum population at 35 mL-1 175 g substrate, inoculated on 30th day showed negative correlation with a R2 value of 0.682 (y = -0.067+11.17) which was highly significant at 0.1% level. The viable population of A. chroococcum inoculated at the rate of 40 mL-1 175 g of substrate on the 0th day of storage was 10, 10, 11 and 8x107 g-1 cfu in the vermicompost, inoculated on 0, 10, 20 and 30th day of vermicomposting, respectively (Table 3). The population of A. chroococcum inoculated on 0 and 10th day of vermicomposting showed 2 and 1 x107 g-1 cfu, on 105 and 120th day of storage, respectively. There after no viable cells were observed upto 180 days. On 20 and 30th day of inoculation the survival of A. chroococcum was observed upto 135th day and the subsequent observations showed no viable population upto 180 days. The change of A. chroococcum population in vermicompost (at 40 mL-1 175 g of substrate) with reference to the storage period (180 days) showed negative correlation with the storage period (p<0.001, Fig. 3).
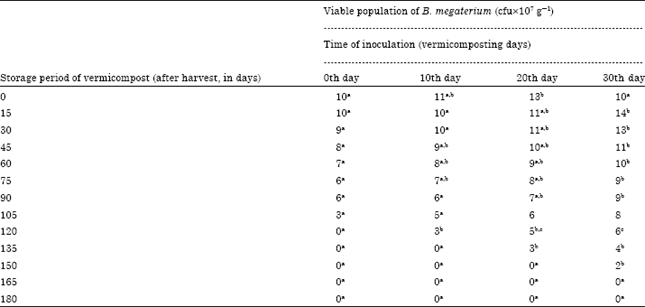
Enrichment studies with B. megaterium: The viable population of B. megaterium inoculated at the rate of 30 mL-1 175 g of substrate in different intervals showed decrease in viable population during the storage of vermicompost. The 30th day inoculation of B. megaterium showed survival upto 150 days whereas, the B. megaterium inoculated on 10 and 20th day showed the viable population upto 120th and 135th day, respectively; 0th day inoculation showed viable population upto 105 days of storage period (Table 4).
The decrease of viable population upon storage of vermicompost which received B. megaterium inoculum at different intervals was negatively correlated (Fig. 4). B. megaterium population at 30 mL-1 175 g substrate, inoculated on 30th day showed negative correlation with a R2 value of 0.850 (y = -0.072+14) which was highly significant at 0.1% level. The B. megaterium inoculated at the rate of 35 mL-1 175 g substrate showed the viable cells in 10-7 dilution upto 135, 150 and 165th day of storage in 10, 20 and 30th day of inoculation, respectively (Table 5).
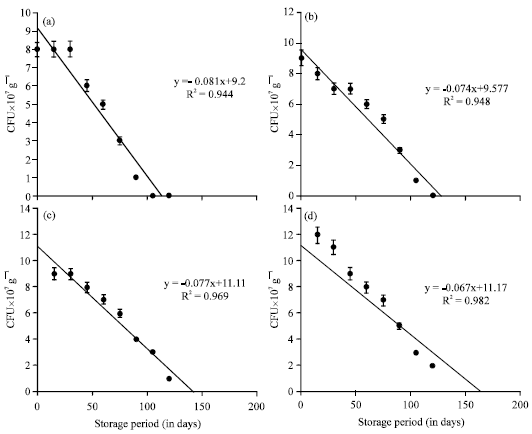 | |
| Fig. 2(a-d): | The change of A. chroococcum population in the vermicompost (at 35 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of A. chroococcum during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
| Table 3: | The viable population of A. chroococcum inoculated at the rate of 40 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
 | |
| Fig. 3(a-d): | The change of A. chroococcum population in the vermicompost (at 40 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of A. chroococcum during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
| Table 4: | The viable population of B. megaterium inoculated at the rate of 30 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
 | |
| Fig. 4(a-d): | The change of B. megaterium population in the vermicompost (at 30 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of B. megaterium during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
| Table 5: | The viable population of B. megaterium inoculated at the rate of 35 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
B. megaterium inoculated on 0, 10 and 20th day showed a decline in viable population during storage period. In 30th day of inoculation, the viable population initially increased, then decreased.
 | |
| Fig. 5(a-d): | The change of B. megaterium population in the vermicompost (at 35 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of B. megaterium during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
The viable population of 10, 15 and 13 cfux107 g-1 were observed on 0, 15 and 30th day of storage of vermicompost which received B. megaterium inoculum on 30th day of vermicomposting. Then, colonies in the same vermicompost showed decline upto 165 days of storage. The change of B. megaterium population in vermicompost which received the inoculum level of 35 mL-1 175 g substrate during the storage period showed negative correlation. The 30th day of inoculation of B. megaterium during vermicomposting showed viable population in 10-7 dilution for a maximum storage period i.e., 165 days. The decrease of B. megaterium population showed significant negative correlation with storage period (p<0.001) (Fig. 5).
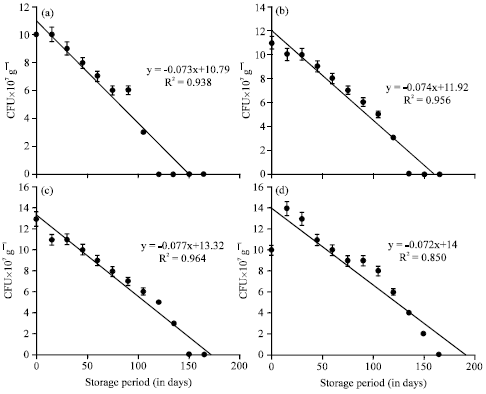
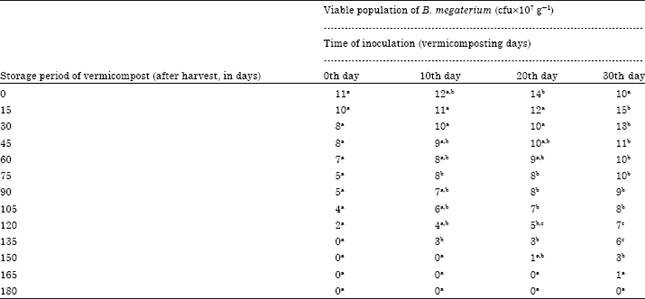
The viable population of B. megaterium inoculated at the rate of 40 mL-1 175 g of substrate on the 0th day of storage was 12,13, 14 and 11x107 g-1 cfu in the vermicompost, inoculated on 0, 10, 20 and 30th day of vermicomposting, respectively (Table 6). The population of B. megaterium inoculated on 0th day and 10th day of vermicomposting showed 1 and 2x107 g-1 cfu on 120 and 135th day, respectively there after no viable cells were observed upto 180 days. On 20 and 30th day of inoculation the survival of B. megaterium was observed upto 150 and 165th day, respectively and the subsequent observations showed no viable population upto 180 days. The change of B. megaterium population in vermicompost (at 40 mL-1 175 g of substrate) with reference to the storage period (180 days) showed that negative correlation with storage period (p<0.001, Fig. 6). The viable population of B. megaterium inoculated at the rate of 40 mL-1 175 g of substrate on 30th day of vermicomposting showed 14, 11, 9, 7, 4 and 2x10-7 g-1 cfu during 30, 60, 90, 120, 150 and 165th day, respectively which was negatively significant at p<0.001 (y = -0.081x +14.29, R2 = 0.876) (Fig. 6). On 0th, 10th and 20th day of inoculation, decrease in the viable population of B. megaterium towards the progression of storage period of vermicompost was observed.
| Table 6: | The viable population of B. megaterium inoculated at the rate of 40 mL per 175 g of substrate in different intervals along the storage period of vermicompost. Values are rounded of mean values of three replicates |
 | |
| The values with same superscript letters between columns are not significantly different at 5% level (p<0.05) by Duncan’s Multiple range test | |
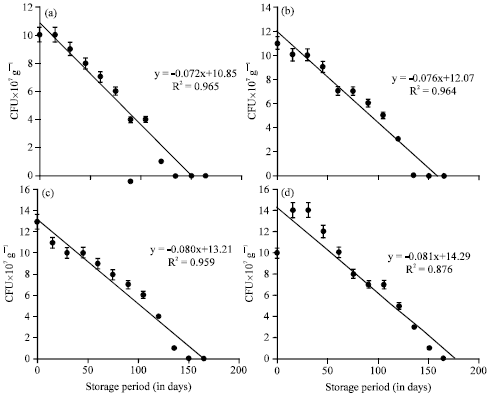 | |
| Fig. 6(a-d): | The change of B. megaterium population in the vermicompost (at 40 mL-1 175 g of substrate) during storage period (180 days). Error bars Indicate±SD; Time of inoculation of B. megaterium during vermicomposting: (a) 0th day, (b) 10th day, (c) 20th day and (d) 30th day |
Whereas, the 30th day inoculation of B. megaterium showed increase in viable population within 15 days and declined gradually towards the termination of the study. Among the different time of inoculation of B. megaterium, the 30th day showed viable population of 2x107 g-1 cfu upto 165 days. In all the four treatments, the viable population of B. megaterium from 0th day onwards showed statistically significant decline with that of storage period of vermicompost (Table 6, Fig. 6).
Earthworm activity is closely associated with microbial activity. Lavelle (1983), is of the opinion that there may exist competition between microorganisms and earthworms for easily digestible and energy rich substrates. Such competition may depend on availability of nutrients in the medium. Contrary to this, earthworms may derive benefit from microorganisms when they have to survive on materials rich in cellulose or hemicellulose. So, there exists mutualistic relation between earthworms and microorganisms. The enrichment of vermicompost with nutrients and microorganisms using different organic and inorganic materials and microbial inoculants is now popularizing, due to the advantage of using the ‘enriched vermicompost’ (Singh and Sharma, 2002; Anilkumar et al., 2007; Hashemimajd and Golchin, 2009). But these studies have not described the standardized protocol for enriching vermicompost with microbial inoculants such as biofertilizers.
The enrichment of vermicompost was done in the present study by inoculating the biofertilizer inoculants, A. chroococcum and B. megaterium at the rate of 30, 35 and 40 mL-1 175 g of substrate. This was done to find out the optimum level of inoculum required for the maintenance of 1x107 viable cells in the vermicompost. The inoculation was done on 0, 10, 20 and 30th day of vermicomposting to assess whether the time of inoculation had any effect on survival rate of biofertilizer inoculants during storage. At the same time, the survival rates of each inoculant were correlated with total microbial population in the vermicompost to study the influence of other microbial groups on biofertilizer inoculants. The change of A. chroococcum and B. megaterium in the vermicompost (at 30, 35 and 40 mL-1 175 g substrate) with reference to the storage period (180 days) showed highly significant negative correlation (p<0.001). In all the four treatments, the viable population of A. chroococcum and B. megaterium at the rate of 30, 35 and 40 mL-1 175 g substrate from 0th day (after harvest) onwards showed statistically significant decline with that of storage period of vermicompost.
The enrichment of vermicompost with the addition of nutrient rich substrates was demonstrated by Daniel et al. (2010) where their results reveal that the leaves of Gliricidia sepium and Leucaena leucocephala can be converted into microbial-and nutrient-rich vermicompost using E. fetida. Hashemimajd and Golchin (2009) studied the effect of iron-enriched vermicompost on growth and nutrition of tomato and reported that total and available forms of iron in iron-enriched vermicomposts as well as in tomato tissues increased by an increase in the proportion of iron refuse in vermicompost. Some of the authors tried microbial inoculants for hastening the process of vermicomposting or for enriching the vermicompost. The inoculation of microbial consortia like ‘jeevamrutha’ and cow dung together with organic substrates significantly enhances the microbial density throughout the process of decomposition during vermicomposting (Veeresh et al., 2010). The inoculation of consortium of microorganisms Aspergillus niger, P. sajor-caju, Azotobacter chroococum, Trichoderma harzianum not only accelerated vermicomposting of crop residues and farm yard manure but also enriched the quality of product (Singh and Sharma, 2002). During the incubation period the inoculated bacterial strains proliferated rapidly, fixed nitrogen and solublised added and native phosphate (Kumar and Singh, 2001).
Press-mud alone and in combination with other by-products of sugar processing industries was pre-decomposed for 30 days by inoculation with combination of Pleurotus sajorcaju, Trichoderma viridae, Aspergillus niger and Pseudomonas striatum, followed by vermicomposting for 40 days with the native earthworm, Drawida willsi. The study conducted by Prabha et al. (2007) reported that the enrichment generally had a significant effect on the nutrient content, especially for N, P, K, Mg and Mn. Eudrilus compost, when treated with Azospirillum and P-solubilising organisms, gave a N-content of 2.08% which was significantly higher than the N-content of uninoculated Eudrilus compost (1.8%). The nitrogen was enriched appreciably by Azospirillum. The enrichment increased progressively when Azospirillum inoculation was supplemented with phosphate solubilising culture, a beneficial additive to obtain good quality compost, rich in N (Tiwari et al., 1989). An increase in N-content due to microbial inoculation was reported by Rasal et al. (1988). The P-contents were significantly higher when inoculated with Azospirillum and P-solubilising organisms (1.76%) than in uninoculated compost (0.72%). The mechanism of conversion of insoluble P by P-solubilising organisms to available forms include altering the solubility of inorganic compounds to the ultimate soluble form by production of acids and H2S under aerobic and anaerobic conditions and by mineralizing organic compounds, with the release of inorganic phosphate (Rasal et al., 1988).
Kumar and Shweta (2011b) studied the enhancement of wood waste decomposition by microbial inoculation prior to vermicomposting. The timber wastes which were inoculated with different combinations of the fungi Phanerochete chrysosporium, Trichoderma reesei, Aspergillus niger and the bacteria Azotobacter chroococcum (MTCC 3853) and Bacillus cereus (MTCC 4079) and incubated at 28-30°C in a mechanical composter. The inoculation enhanced the degradation of timber wastes, increased total nitrogen and improved the quality and enhanced production of vermicompost generated with the native earthworm Drawida willsi Michelsen. Their study showed that microbial predecomposition of timber wastes to produce quality vermicompost is a feasible technology.
However, the above studies were focused on either the enhancement of vermicomposting process or the nutrients. These studies have not described the standardization of amount of inoculum and time of inoculation for the maintenance of 1x107 g-1 viable population of A. chroococcum and B. megaterium and their survival during storage in comparison with the total microbial population. From the results of this study, it is concluded that the biofertilizer inoculants, A. chroococcum and B. megaterium at the rate 35 mL-1 175 g substrate on 30th day of vermicomposting is the optimum inoculation level and time for the maintenance of 1x107 viable cells in the vermicompost is maximum number of days during storage. The study also reveals that the microbial inoculants inoculated at the later stage of vermicomposting survive for long period.
TOTAL MICROBIAL POPULATION IN THE VERMICOMPOST
Total microbial population in the vermicompost inoculated with A. chroococcum at 30, 35 and 40 mL-1 175 g substrate: The total microbial population in A. chroococcum inoculated vermicompost was high during the initial phases of storage and then total microbial population (1x107 cfu g-1) declined towards the end (Table 7). The bioinoculation on 0th day of vermicomposting with A. chroococcum at the rate of 30 mL-1 175 g substrate showed 14, 11, 6, 1 and 0x107 g-1 cfu population of total microorganisms, respectively during 0, 30, 60, 90 and 120th day of storage. No viable population of total microorganisms in 10-7 dilution on 105, 120, 135 and 150th day of storage was observed, respectively in 0, 10, 20 and 30th day inoculation of A. chroococcum at the rate of 30 mL-1 175 g substrate. On 20th day of vermicomposting with A. chroococcum at the rate of 35 mL-1 175 g of substrate showed 15, 11, 8, 4, 1 and 0x107 g-1 cfu population of total microorganisms, respectively during 15, 45 75, 105, 135 and 165th day of storage (Table 7). Similar observations were made for the inoculum level of 40 mL-1 175 g substrate.
| Table 7: | The total microbial population change in A. chroococcum and B. megaterium inoculated vermicompost during storage period. Values are rounded of mean values of three replicates |
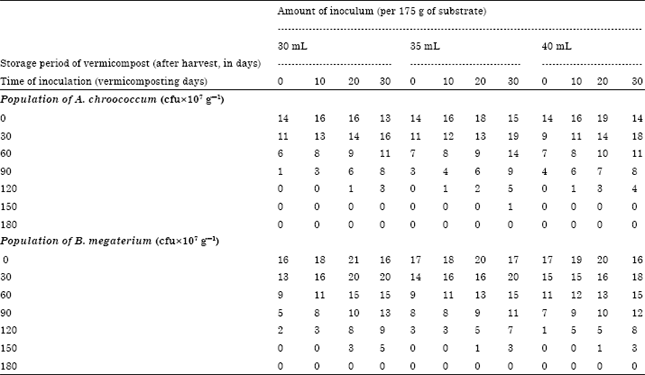 | |
Total microbial population in the vermicompost inoculated with B. megaterium at 30, 35 and 40 mL-1 175 g substrate: B. megaterium inoculated at the rate of 30 mL-1 175 g of substrate on 0th day of vermicomposting showed 16, 13, 9, 5, 2 and 0x107 g-1 cfu population of total microorganisms, respectively during 0, 30, 60, 90, 120 and 150th day of storage. The total microbial population in B. megaterium inoculated vermicompost was high during the initial phases of storage and then total microbial population (1x107 g-1 cfu) declined towards the end (Table 7). The microbial inoculants inoculated on 20th day of vermicomposting with B. megaterium at the rate of 35 mL-1 175 g of substrate showed 17, 15, 11, 7, 3 and 0x10-7 g-1 cfu population of total microorganisms, respectively during 15, 45 75, 105, 135 and 165th day of storage. B. megaterium at the rate of 35 mL-1 175 g of substrate inoculated on 20th day of vermicomposting showed 16, 18, 15, 12, 8, 3 and 0x10-7 g-1 cfu population of total microorganisms, respectively during 0, 30, 60, 90, 120, 150 and 165th day of storage.
CORRELATION OF TOTAL MICROBIAL POPULATION WITH STORAGE PERIOD
The change of total microbial population in A. chroococcum inoculated vermicompost (at 30 mL-1 175 g substrate) as a function of storage period (180 days) showed negative correlation in all the four treatments that received A. chroococcum during 0, 10, 20 and 30th day of vermicomposting. The vermicompost inoculated with A. chroococcum on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient (r): -0.9840 (y = -0.1344x+14.4). Similar results were recorded for 10th day inoculation: r = -0.9794, y = -0.1119x+15.231; for 20th day inoculation: r = -0.9794, y = -0.1224x+16.909; for 30th day inoculation: r = -0.9541, y = -0.1152x+17.833.
The total microbial population change in A. chroococcum inoculated vermicompost (at 35 mL-1 175 g substrate) as a function of storage period (180 days) showed negative correlation in all the four treatments that received A. chroococcum during 0, 10, 20 and 30th day of vermicomposting. The vermicompost inoculated with A. chroococcum on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient of r = -0.9913, y = -0.1089x+13.422. Similar results were recorded for 10th day inoculation: r = -0.9757, y = -0.1079x+14.874; for 20th day inoculation: r = -0.9948, y = -0.1188x+16.818; for 30th day inoculation: r = -0.9542, y = -0.127x+20.731. The vermicompost inoculated with A. chroococcum at the rate of 40 mL-1 175 g substrate on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient of r = -0.991, y = -0.1089x+13.422. Similar results were recorded for 10th day inoculation: r = -0.9710, y = -0.1008x+14.421; for 20th day inoculation: r = -0.994, y = -0.1255x+18.136; for 30th day inoculation: r = -0.9558, y = -0.1193x+18.679.
Similar trend of results were observed for B. megaterium during vermicomposting. The vermicompost inoculated with B. megaterium (at 30 mL-1 175 g substrate) on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient of r = -0.9142, y = -0.0861x+12.132. Similar results were recorded for 10th day inoculation: r = -0.942, y = -0.1011x+14.637; for 20th day inoculation: r = -0.972, y = -0.1051x+16; for 30th day inoculation: r = -0.9538, y = -0.1088x+17.484.
B. megaterium inoculated at 35 mL-1 175 g substrate on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient of r = -0.9913, y = -0.1089x+13.422. Similar results were recorded for 10th day inoculation: r = -0.9757, y = -0.1079x+14.874; for 20th day inoculation: r = -0.9948, y = -0.1188x+16.818; for 30th day inoculation: r = -0.9543, y = -0.127x+20.731. The vermicompost inoculated with B. megaterium at 40 mL-1 175 g substrate on 0th day of vermicomposting showed significant (p<0.001) negative correlation with a correlation coefficient of r = -0.9913, y = -0.1089x+13.422. Similar results were recorded for 10th day inoculation: r = -0.9790, y = -0.1008x+14.421; for 20th day inoculation: r = -0.9941, y = -0.1255x+18.136; for 30th day inoculation: r = -0.9558, y = -0.1193x+18.679.
CORRELATION OF POPULATION DYNAMICS BETWEEN TOTAL MICROFLORA AND INDIVIDUAL MICROBIAL INOCULANTS
Total microbial population vs A. chroococcum: Total microbial population in the vermicompost inoculated with A. chroococcum on 0, 10, 20 and 30th day of vermicomposting at the rate 30, 35 and 40 mL-1 175 g each showed a significant positive correlation (Table 8). Similar positive correlation was recorded for different time of inoculation and with different inoculum levels as indicated in the table. The viable population of A. chroococcum inoculated (35 and 40 mL-1 175 g substrate) on 0th day of vermicomposting was significantly correlated with total microbial population recorded during storage (Table 8).
Total microbial population vs B. megaterium: The viable population of B. megaterium inoculated (30 mL-1 175 g) on 0th day of vermicomposting was significantly correlated with total microbial population recorded during storage (r = 0.9534; y = 0.6328x+0.648; p<0.001). Similar positive correlation was recorded for different time of inoculation and with different inoculum levels (Table 8). In the present study, total microbial population in A. chroococcum and B. megaterium inoculated vermicompost was high during the initial phases of storage and then total microbial population declined towards the end suggesting that the overall maintenance of total microbial population in vermicompost is similar in the vermicomposts with any microbial inoculant.
| Table 8: | Correlation of population dynamics between total microflora and A. chroococcum and B. megaterium during storage of enriched vermicompost (180 days) |
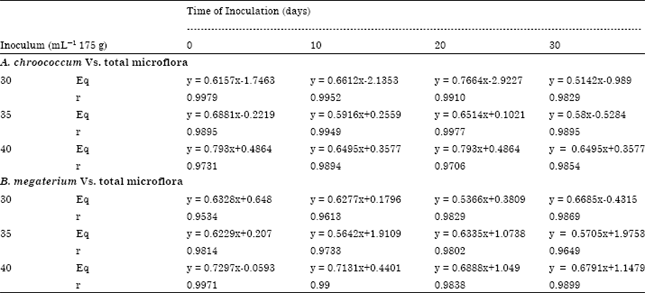 | |
| Eq: Regression equation; r: Correlation co-efficient | |
The current study results on the change of total microbial population in A. chroococcum and B. megaterium inoculated vermicompost (at 30, 35 and 40 mL-1 175 g substrate) as a function of storage period (180 days) showed negative correlation in all the four treatments received the microbial inoculants during 0, 10, 20 and 30th day of vermicomposting. The same trend of results was obtained in the total microbial population change in B. megaterium inoculated vermicompost which also showed negative correlation with storage period in all treatments. The results on the correlation of total microbial population with the individual microbial inoculants, A. chroococcum and B. megaterium in respective vermicompost during storage period showed significant positive correlation, i.e., the increase/decrease of individual inoculants and total microbial population were parallel. These results clearly show that the population of microbial inoculants and the total microbial population in the vermicompost are dependents of each other. However, the competition between these two groups for nutrients requires further insight. There are many studies focusing the increase of microbial population in earthworm excreted or processed material than the parent material. Recent developments in the country as well as at the global level is the application of detritivorous epigeic earthworms for organic manure/vermicompost production from biodegradable organic materials recovered from agricultural lands, agro-based industries and municipal solid waste. This field of study is closely associated with earthworm microbe interaction. The quality of the manure or vermicompost depends on microorganisms associated with the process of decomposition. Earthworm activity is closely associated with microbial activity.
Lavelle (1983), is of the opinion that there may exist competition between microorganisms and earthworms for easily digestible and energy rich substrates. Such competition may depend on availability of nutrients in the medium. Contrary to this, earthworms may derive benefit from microorganisms when they have to survive on materials rich in cellulose or hemicellulose. So, there exists mutualistic relation between earthworms and microorganisms. Tiunov and Scheu (2004) have shown that earthworms deprive easily available carbon to microorganisms and availability of carbon increases effective mobilization of N and P by earthworms. Earthworms are mainly responsible for fragmentation and conditioning of the substrate, increasing surface area for microbial activity and significantly altering biological activity of the process (Dominguez et al., 2003). The survival and increase of microbial population in vermicasts and worm-worked compost (vermicompost) falls in line with regard to the present study results.
The enhanced survival rate of A. brasilense, A. chroococcum, B. megaterium and R. leguminosarum in the present study might be due to the presence of nutrients and other factors reported in the following studies. There are reports that earthworms influence the abundance and activity of soil microorganisms (Brown et al., 2000; Scheu et al., 2002), either by changes in the physical structure of the soil, e.g., soil aggregation (Winding et al., 1997) or by direct interactions when microorganisms are ingested together with mineral soil and organic material. During gut passage, some microbes are digested while others survive and may be stimulated by the rich nutrient supply (Winding et al., 1997; Brown et al., 2000). The correlation between the physico-chemical parameters and microbial populations of the casts of P. corethrurus showed that the establishment of microbial population requires optimum moisture, organic carbon and nitrogen content (Karmegam and Daniel, 2000). The incubation of vermicasts (45 days) showed significant correlation with that of the increase in fungal population (r = 0.720; p<0.05), decrease in moisture content (r = -0.984; p<0.001) and the decrease in moisture content statistically had no effect on the total fungal population in the vermicasts of P. ceylanensis (Prakash et al., 2008). Further, their study showed that the total microbial population, viz., bacteria, fungi and actinomycetes were found to be many-fold higher than in the initial vermibed substrate and in substrate without earthworms (control). Similar studies by Sekar and Karmegam (2009) and Prakash and Karmegam (2010) with different vermibed substrates revealed that the microbial population increased in vermicompost than in compost. These studies well support well the findings of the present study which showed the similar results where the vermicompost served as a substrate for the survival and viability of the biofertilizer inoculants, A. chroococcum and B. megaterium for long period.
CONCLUSION
In the present study, the inoculum level of A. chroococcum and B. megaterium at the rate of 35 mL-1 175 g of vermibed substrate is sufficient to maintain 1x107 viable cells up to 160 days after harvesting of vermicompost. The inoculum of biofertilizer organisms into vermibed on 30th day showed increased survival rate and hence, the optimized inoculation of 35 mL of inoculum per 175 g of substrate on 30th of vermicomposting is helpful for the maintenance of sufficient viable population for more than five months in the enriched vermicompost.
ACKNOWLEDGMENT
The authors sincerely thank Prof. Thilagavathy Daniel, Department of Biology, Gandhigram Rural Institute, Gandhigram, India for her suggestions and constant encouragements.
REFERENCES
- Abo-Baker, A.A. and G.G. Mostafa, 2011. Effect of bio-and chemical fertilizers on growth, sepals yield and chemical composition of Hibiscus sabdariffa at new reclaimed soil of south valley area. Asian J. Crop Sci., 3: 16-25.
CrossRefDirect Link - Alidadi, H., A.R. Parvaresh, M.R. Shahmansouri, H. Pourmoghadas and A.A. Najafpoor, 2007. Combined compost and vermicomposting process in the treatment and bioconversion of sludge. Pak. J. Biol. Sci., 10: 3944-3947.
CrossRefPubMedDirect Link - Ansari, A.A., 2011. Worm powered environmental biotechnology in organic waste management. Int. J. Soil Sci., 6: 25-30.
CrossRefDirect Link - Barassi, C.A., R.J., Sueldo, C.M. Creus, L.E. Carrozzi, E.M. Casanova and M.A. Pereyra, 2007. Azospirillum spp., a dynamic soil bacterium favourable to vegetable crop production. Dyn. Soil Dyn. Plant, 1: 68-82.
Direct Link - Brown, G.G., I. Barois and P. Lavelle, 2000. Regulation of soil organic matter dynamics and microbial activityin the drilosphere and the role of interactionswith other edaphic functional domains. Eur. J. Soil Biol., 36: 177-198.
CrossRefDirect Link - Dominguez, J., R.W. Parmelee and C.A. Edwards, 2003. Interactions between Eisenia andrei (Oligochaeta) and nematode populations during vermicomposting. Pedobiologia, 47: 53-60.
CrossRefDirect Link - Hashemimajd, K. and A. Golchin, 2009. The effect of iron-enriched vermicompost on growth and nutrition of tomato. J. Agric. Sci. Technol., 11: 613-621.
Direct Link - Jayashree, S., J. Rathinamala and P. Lakshmanaperumalsamy, 2011. Determination of heavy metal removal efficiency of Chrysopogon zizanioides (Vetiver) using textile wastewater contaminated soil. J. Environ. Sci. Technol., 4: 543-551.
CrossRefDirect Link - Kalam, A., J. Tah and A.K. Mukherjee, 2004. Pesticide effects on microbial population and soil enzyme activities during vermicomposting of agricultural waste. J. Environ. Biol., 25: 201-208.
PubMed - Kaushik, P., Y.K. Yadav, N. Dilbaghi and V.K. Garg, 2008. Enrichment of vermicomposts prepared from cow dung spiked solid textile mill sludge using nitrogen fixing and phosphate solubilizing bacteria. Environmentalist, 28: 283-287.
CrossRefDirect Link - Kumar, R. and Shweta, 2011. Removal of pathogens during vermin-stabilization. J. Environ. Sci. Technol., 4: 621-629.
CrossRefDirect Link - Mostafa, G.G. and A.A. Abo-Baker, 2010. Effect of bio- and chemical fertilization on growth of sunflower (Helianthus annuus L.) at South valley area. Asian J. Crop Sci., 2: 137-146.
CrossRefDirect Link - Padmavathiamma, P.K., L.Y. Li and U.R. Kumari, 2008. An experimental study of vermin-biowaste composting for agricultural soil improvement. Bioresour. Technol., 99: 1672-1681.
CrossRefDirect Link - Prakash, M. and N. Karmegam, 2010. Vermistabilization of pressmud using Perionyx ceylanensis Mich. Bioresour. Technol., 101: 8464-8468.
CrossRefDirect Link - Sekar, K.R. and N. Karmegam, 2009. Suitability of vermicasts as carrier material for a biofertilizer, Azosprillum brasilense (MTCC 4036). Dyn. Soil Dyn. Plant, 3: 143-146.
Direct Link - Sekar, K.R. and N. Karmegam, 2010. Earthworm casts as an alternate carrier material for biofertilizers: Assessment of endurance and viability of Azotobacter chroococcum, Bacillus megaterium and Rhizobium leguminosarum. Sci. Hortic., 124: 286-289.
CrossRefDirect Link - Saikia, S.P. and V. Jain, 2007. Biological nitrogen fixation with non-legumes: An achievable target or a dogma. Curr. Sci., 92: 317-322.
Direct Link - Scheu, S., N. Schmitt, A.V. Tiunov, J.E. Newington and T.H. Jones, 2002. Effects of the presence and community composition of earthworms on microbial functioning. Oecologia, 133: 254-260.
Direct Link - Singh, A. and S. Sharma, 2002. Composting of a crop residue through treatment with microorganisms and subsequent vermicompost. Bioresour. Technol., 85: 107-111.
CrossRef - Sinha, R.K., S. Agarwal, K. Chauhan, V. Chandran and B.K. Soni, 2010. Vermiculture technology: Reviving the dreams of Sir Charles Darwin for scientific use of earthworms in sustainable development programs. Technol. Invest., 1: 155-172.
CrossRefDirect Link - Tiunov, A.V. and S. Scheu, 2004. Carbon availability controls the growth of detritivores (Lumbricidae) and their effect on nitrogen mineralization. Oecologia, 138: 83-90.
CrossRefPubMedDirect Link - Tiwari, S.C., B.K. Tiwari and R.R. Mishra, 1989. Microbial populations, enzyme activities and nitrogen-phosphorus-potassium enrichment in earthworm casts and in surrounding soil of a pineapple plantation. Biol. Fert. Soils, 8: 178-182.
CrossRef - Winding, A., R. Ronn and N.B. Hendriksen, 1997. Bacteria and protozoa in soil microhabitat as affected by earthworm. Biol. Fertil. Soils, 24: 133-140.
CrossRef - Yadav, J., J.P. Verma and K.N. Tiwari, 2011. Plant growth promoting activities of fungi and their effect on chickpea plant growth. Asian J. Biol. Sci., 4: 291-299.
CrossRefDirect Link - Zularisam, A.W., Z.S. Zahirah, I. Zakaria, M.M. Syukri, A. Anwar and M. Sakinah, 2010. Production of biofertilizer from vermicomposting process of municipal sewage sludge. J. Applied Sci., 10: 580-584.
CrossRefDirect Link
Narayanaswamy Dharmalingam Reply
Great vision, to uplift the science & technology.
Natchimuthu Karmegam
Thank you for your comments.