
Rahul Kumar
Department of Zoology, Vermiculture Research Station, D.S. College (Dr. B.R.A. University) Aligarh- 202001 UP, India
Shweta
Department of Zoology, Vermiculture Research Station, D.S. College (Dr. B.R.A. University) Aligarh- 202001 UP, India
Journal of Environmental Science and Technology
Year: 2011 | Volume: 4 | Issue: 6 | Page No.: 621-629







ABSTRACT
Earthworm promises to provide cheaper solutions to several social, economic and environmental problems plaguing the human society. Their body work as ‘bio-filter’ and they can ‘purify’ and also ‘disinfect’ and ‘detoxify’ the solid wastes. They are both protective and productive for environment and society. The microbiomics of gut worm (Eisenia foetida Savigny) and their association with vermistabilization was studied to determine the quantification of microbial groups in reactors. Worms were reared in three reactors viz. rice, straw, sugarcane trash, kitchen waste along with cow dung in 2:1 ratio for seventy days. The microbial load (Pseudomonas sp., Streptosporangium sp., Salmonella sp., Shigella sp. Flexibacter sp. and Escherichia sp.) in gut and cast, total nitrogen, potassium, organic carbon in reactors was determined, periodically. The microbial population in gut and reactors were determined using pour plate method The microbiomics of gut and casting revealed the removal of Salmonella (12-17x103±0.02 to 0-4x103±0.05 cfu g-1), Shigella (3-5x103±0.04 to 0.00±0.08 cfu g-1), Flexibacter (6-10x104±0.05 to 00.0±0.05 cfu g-1), Escherichia (8x102±0.02 to 0.00-1x101±0.05 cfu g-1) during vermistabilization, indicating the selective nature of feeding of earthworms which helps in the removal of pathogens as they enter in food chain of worms. However, increase in the number of Pseudomonas (13-16x104±0.05 to 18-22x105±1.0 cfu g-1), Streptosporangium (12-14x104±0.05 to 18-20x105±1.0 cfu g-1) seems to be responsible for reduction of pathogens and mineralization of other organics during the waste degradation process.
PDF Abstract XML References Citation
Received: April 25, 2011;
Accepted: May 26, 2011;
Published: July 19, 2011
How to cite this article
Rahul Kumar and Shweta, 2011. Removal of Pathogens during Vermi-Stabilization. Journal of Environmental Science and Technology, 4: 621-629.
DOI: 10.3923/jest.2011.621.629
URL: https://scialert.net/abstract/?doi=jest.2011.621.629
DOI: 10.3923/jest.2011.621.629
URL: https://scialert.net/abstract/?doi=jest.2011.621.629
INTRODUCTION
A revolution is unfolding in vermiculture studies for multiple uses in environmental protection and sustainable development. Earthworms have over 600 million years of experience as ‘environmental managers’ in the ecosystem. Vermiculture scientists all over the world knew about the role of earthworms as ‘waste managers’, as’ soil managers and fertility improvers’ and ‘plant growth promoters’ for long time (Sinha et al., 2010). But some ‘new discoveries’ about their role in removal of pathogen may revolutionized the vermiculture industry.
Soil, the major reservoir of microbes, meets the food requirement of earthworms and this has necessitated the establishment of different kinds of relationships viz., a part of food for earthworm, proliferated in the gut and reactors. The microbes and earthworm together mineralize, humifies organic matter and also facilitates chelation of some metal ions (Canellas et al., 2002; Rizl and Novokova, 2003; Maboeta and von Rensburg, 2003). Earthworms have the capacity to utilize soil microbes as their food (Parthasarathi and Ranganathan, 1999; Verma and Shweta, 2011).
Selective proliferation of microorganisms in the gut of different kinds of earthworms, due to the environment and food material available for growth of microorganisms have been reported earlier viz., fungi in Perionyx millardi (Chhotaray et al., 2011), Lampito mauritii (Ansari, 2011), Onychochaeta borincana (Barik and Gulati, 2009), bacteria in Amynthas caliginosa (Omrani et al., 2005), Lampito mauritii (Idowu et al., 2006), L. terrestris (Ansari, 2011) and actinomycetes in L. terrestris, A. longa and A. caliginosa (Chhotaray et al., 2011), L. rubellus and A. caliginosa (Ansari, 2011). Higher microbial population and the activity in the casts of earthworms compared to surrounding soils have been demonstrated by Tangjang et al. (2009). The available information on solid waste management suggests that solid wastes should be biocomposted before applying to soil in order to achieve biological transformation of the organic matter and to avoid potential risk of pathogens (Masciandaro et al., 2000; Ansari and Saywack, 2011). Biocomposting of solid wastes brings about stabilization of the organic matter and effectively reduces pathogen concentrations (Kumar and Shweta, 2011). However, absolute removal of pathogens difficult to achieve and many pathogens survive the composting process (Sinha et al., 2009b). Incorporation of earthworm in biocomposting process has been considered to be an appropriate technology for bio waste management for producing nutrient enriched vermicompost. Various investigators have, therefore, established the viability of the technology using earthworms as a treatment system for different wastes (Ansari and Sitaram, 2011; Elviror et al., 1997).
Much of the research on vermistabilization has been focused on changes in the chemical parameters. The information available on the microbiota which determine the rate of degradation is however, very little (Edwards and Bohlen, 1996a; Lavelle et al., 1998). Hence, the present investigations were under taken and cover the dynamics and quantification of microbial groups during vermistabilization process, changes in composition of microbes in the gut, casting and their association with stabilization.
MATERIALS AND METHODS
The experiment was carried out in 2009-2010.Worms (Eisenia foetida Savigny) were reared in three reactors viz., rice straw and cow dung (RC) 2:1 ratio, sugarcane trash and cow dung (SC) 2:1 ratio and vegetable waste and cow dung (VC) 2:1 ratio. The reactors were mixed well and moistened with water up to 40% moisture contents. These treatment groups were investigated in two replicates; the reactor were placed in fiber bins and kept in natural environment to maintain favorable temperature for worms, as they are highly sensitive to temperature fluctuations. Two hundred mature earthworms were introduced into the bins containing 20 kg reactors each.
The treatment groups of different composition were allowed to stabilize for 70 days. Samples from the entire units were taken out, monthly, to measure the biomass of individuals. The worms were washed with distilled water and were weighed. After weighing, they were immediately placed in bins to prevent their desiccation.
During stabilization process of the reactors, five earthworms were collected and washed with sterile water to remove the surface microbial flora from its outer skin. The earthworms were then sacrificed by freezing and their whole body was dissected. The gut region approximately (3-5 cm) was taken for analysis for the reason the higher glutamic activity favours the microbial growth as reported by Edwards (1998) and Lavelle et al. (1998). Microbial analysis was carried out immediately following Lavelle et al. (1998).
For microbial analysis of castings, worms were collected from all the experimental units and rinsed thoroughly in sterile water. The worms were then placed in petriplates containing tissue paper for 5 min. The fresh casts deposited were analyzed for different microbial count.
The castings were analyzed at interval of 30 days. The total nitrogen, total potassium and total organic carbon measured by following Shaw and Beadle (1949), Pearson (1952) and Walkey and Black (1934), respectively.
Statistical analysis: Results are the means of three replicates. One way Analysis of Variance (ANOVA) was done using the INDOSTAT programme. The objective was to determine any significant differences among the parameters analyzed in different reactors during the stabilization process.
RESULTS
General: The average body weight (g) of earthworm was monitored on 30th and 60th days of vermistabilization (Table 1). No mortality was observed throughout experiment. The biomass (weight) increased from 0.64±0.02 to 1.2±0.02 in RC and SC and 0.64±0.02 to 0.84±0.02 in VC reactor. The readily available nutrients in RC and SC might have enhanced the feeding activity of worms (Kumar and Shweta, 2011) in present study.
Chemical analysis
Total organic carbon: During the process of stabilization process worms bio-transform waste constituents into a more useful fertilizer by grinding and digesting organic wastes with the help of aerobic and anaerobic micro flora. The percentage of organic carbon was remarkably decreased from 34.42±0.01 to 32.93±0.01 in RC; 36.39±0.05 to 32.82±0.02 in SC; 36.41±0.05 to 32.39±0.04 in VC at 30th day and remains ±30% at 60th day in all reactor. The total organic carbon was metabolized into CO2 by various micro-floras present in gut of worm and in reactor, involved in the vermistabilization (Table 2).
| Table 1: | Biomass (weight) of worms during stabilization process of reactors |
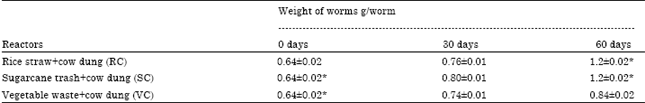 | |
| All values are mean and standard deviation of the replicates. *Significant (p<0.01) | |
| Table 2: | Chemical analysis of reactors during vermistabilization |
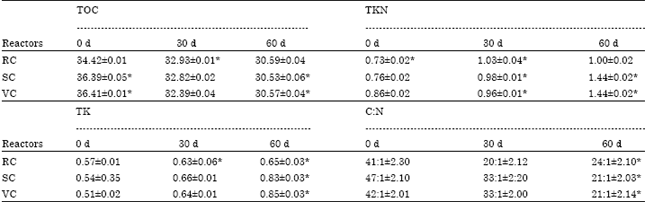 | |
| All values are mean and standard deviation of the replicates. *Significant (p<0.01), RC: Rice straw+cow dung, SC: sugarcane trash+cow dung, VC: Vegetable waste+cow dung | |
Total kjeldahl nitrogen (TKN): The total nitrogen content of the vermicompost increased with time due to rapid mineralization of organic nitrogenous compounds (Table 2). However, the nitrogen content of the final produce depends on the initial nitrogen concentration of the reactor (Kumar and Shweta, 2011). The percentage of nitrogen significantly increased in all reactors viz. 0.73±0.02 to 1.00±0.02 in RC and from 0.76-0.86±0.02 to 1.44±0.02 in SC and VC at the end of vermistabilization. Their enhanced decomposition resulted in lowering of C:N ratio (Talashilkar et al., 1999).
Total potassium: Potassium is an essential micronutrient for both animals and plants. It plays a significant role in food metabolism and maintenance of cellular organization. It activates the enzymes in protein and carbohydrate metabolism. The percentage of potassium were found maximum in VC (0.85±0.003) followed by SC (0.83±0.03) and a slight decrease in RC (0.65±0.03) treatment (Table 2) at 60th day (Table 2).
Microbiomics of gut of worms during stabilization: At the initial stage of stabilization process various pathogens viz. Salmonella, Shigella, Escherichia and Flexibacter were found to be dominant in the gut of worm but during the subsequent days their level decreased significantly (Table 3). However, Pseudomonas and Streptosporangium count increased from 12-16x104±0.05 to 18-22x105±10.0 cfu g-1. The microbial population of Salmonella decreased from 12x103±0.05 to 4x103±0.05 cfu g-1 in RC, 13x103±0.02 to 0.00±0.02 cfu g-1 in SC, 17x103±0.02 to 0.00±0.05 cfu g-1 in VC reactor at 60th day of vermistabilization.
| Table 3: | Microbiomics of gut of worm during stabilization process of reactors |
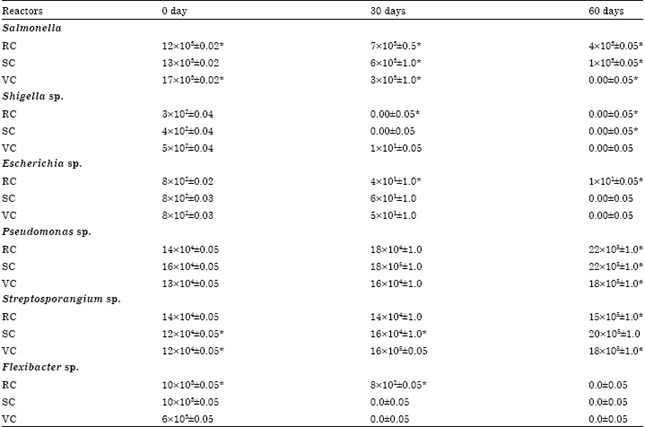 | |
| All values are mean and standard deviation of the replicates and expressed in cfu g-1. *Significant (p<0.01) | |
| Table 4: | Microbiomics in castings of worm during stabilization of reactors |
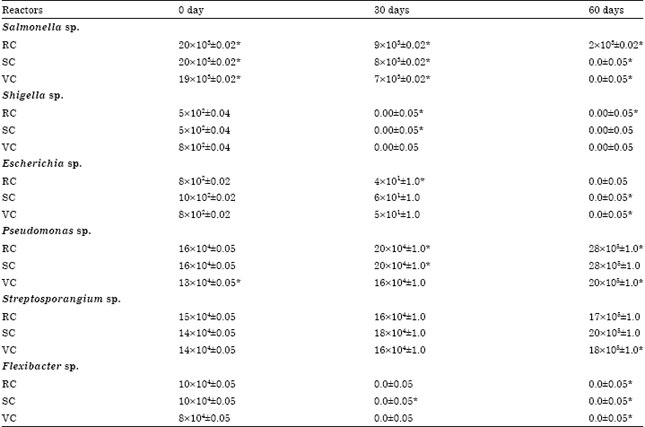 | |
| All values are mean and standard deviation of the replicates and expressed in cfu g-1. *Significant (p<0.01) | |
Similarly, Escherichia count decreases from 8x102±0.02 to 1x101±0.05 cfu g-1 in RC, 8x102±0.03 to 0.00±0.05 cfu g-1 in SC and VC reactor at the 60th day. The reduction in Shigella and Flexibacter recorded at earlier (30th day of stabilization) stage. Shigella decreased from 3-5x102±0.04 to 0.00-1x101±0.05 cfu g-1 in all reactors at 30th day and completely removed at 60th day. Flexibacter sp. observed only in RC reactor (8x102±0.05 cfu g-1) at 30th day of stabilization while, in others completely removed at the same point of time. Population of Pseudomonas gradually increased from 14x104±0.05 to 22x105±1.0 cfu g-1 in RC, 16x104±0.05 to 22x105±1.0 cfu g-1 in SC, 13x104±0.05 to 18x105±1.0 cfu g-1 in VC reactor. The microbial population of Streptosporangium also significantly increased from 12x104±0.05 to 20x105±1.0 cfu g-1 in SC, 12x104±0.05 to 18x105±1.0 cfu g-1 in VC and 14x104±0.05 to 15x105±1.0 cfu g-1 RC reactor.
Microbiomics of cast during stabilization: The casting analysis did not show much difference with microbial count of gut. However, Shigella and Flexibacter were not recorded in any reactor at 30th day of vermistabilization. Salmonella sp. was completely removed out in all reactors except in RC (2x103±0.02 cfu g-1) at 60th day. Similarly, Escherichia was also not recorded in any reactor at same point of time. The significant increase in Pseudomonas (16x104±0.05 to 28x105±1.0 cfu g-1 in RC, 16x104±0.05 to 28x105±1.0 cfu g-1 in SC, 13x104±0.05 to 20x105±1.0 cfu g-1 in VC) and Streptosprangium (15x104±0.05 to 17x105±1.0 cfu g-1 in RC, 14x104±0.05 to 20x105±1.0 cfu g-1 in SC, 14x104±0.05 to 18x105±1.0 cfu g-1 in VC were recorded in castings of all reactors (Table 4).
DISCUSSION
Earthworms routinely devour on the protozoa, bacteria and fungus as food in any substrate where they inhabit (Sinha et al., 2010). They seem to realize instinctively those anaerobic bacteria and fungi are undesirable and so feed upon them preferentially, thus arresting their proliferation. More recently, Sinha et al. (2010) has found in her research that worms living in pathogen-rich materials (e.g., sewage and sludge), when dissected show no evidence of pathogens beyond 5 mm of their gut (Sinha et al., 2009a). The experimental results of microbiomics of gut and casting in present study proved the reduction/removal of Salmonella, Shigella, Escherichia and Flexibacter in all reactors. These results corroborate with the findings of Edwards and Bohlen (1996a), Bohlen and Edwards (1995) who have reported that earthworm’s food source include microorganism and can digest them selectively. Kumar and Sekaran (2005) also reported removal of Salmonella and Escherichia in 35 days in municipal solid wastes by Lampito mauritii.
Bajsa et al. (2003) reported the removal of faecal Coliform, Salmonella, enteric viruses; helminths ova from sewage and sludge appear to be much more rapid when they are processed by E. foetida. In another study Bajsa et al. (2004) reported the pathogen die-off in vermicomposting of sewage-sludge stipends with E. coli and E. faecalis at the 1.6-5.4x106 cfu g-1 and 3- 4x104 cfu g-1, respectively and achieved a safe product in 4-5 months of vermicomposting and the product remained the same quality without much reappearance of pathogens after in the remaining month of the test, confirming the observation of present study. Other studies (Singleton et al., 2003; Safawat et al., 2002) also confirmed significant human pathogen reduction in biosolids, vermicomposted by earthworms.
Increase in population of Streptosporangium and Pseudomonas in present study corroborates the findings of Sidhu et al. (2001) who described the role of Streptosporangium in the suppression of pathogenic bacterial population in earthworm. Kumar and Sekaran (2005), Masciandaro et al. (2000) and Sidhu et al. (2001) also proved that the indigenous micro-flora play a significant role in the suppression of pathogenic bacterial population growth in composted sludge. This confirms the role of Streptosporangium and Pseudomonas in suppression of pathogens in present study. However, there are conflicting observations relative to the proliferation of specific bacterial group in earthworm gut micro flora; indeed passage through the gut may selectively decrease the number of certain microbes such as Shigella and Escherichia (Edwards and Bohlen, 1996b).
Vinceslas-Apka and Loquet (1997) and Lavelle et al. (1998) have reported that organic carbon is metabolized into CO2 and the various microflora present in the gut of worm and in reactor are involved in the degradation of organic carbon. The total nitrogen content of the reactors increased with time, due to rapid mineralization of organic nitrogen compounds. Present results have supported the findings of Bansal and Kapoor (2000) which have shown increase in the nitrogen content as a result of carbon loss in vermicomposting of crop residues and cattle dung. In the present study C:N ratio of reactors lowered because of enhanced decomposition, agreeing to the earlier observations of Talashilkar et al. (1999).
Nevertheless, the removal of pathogens may be due to release of antibiotics by earthworms, responsible to kill pathogenic organisms inhabit and render it virtually sterile (Sinha et al., 2009a). The worms also release the coelomic fluids that have anti-bacterial properties which may destroy all pathogen in reactors (Pierre et al., 1982). Further, there is some thing inside the worms which destroys the pathogens and makes their excreta pathogen free needed to be explored. However, absolute removal of pathogens is difficult to achieve, some may survives even after vermistabilization.
CONCLUSION
The study suggests that the worm selectively inhibits the growth of pathogens in reactors. The presence of pathogenic organisms Salmonella, Shigella, Escherichia and Flexibacter was observed at initial stage of stabilization process but after vermistabilization these were reduced to zero concentration. This amply demonstrates that these pathogens have been eliminated as they entered in food chain of worms. And for this, Pseudomonas, Streptosporangium seems to be responsible for reduction of pathogens and metabolization of reactors. However, it may not be possible to remove pathogens completely but at least worms change the ‘microbial make-up’ of reactors to make it harmless to the soil and enable its use as a nutritive organic fertilizer.
ACKNOWLEDGMENTS
The authors are thankful to the Department of Biotechnology, Ministry of Science and Technology, Government of India, New Delhi, for financial assistance.
REFERENCES
- Ansari, A.A. and K. Sitaram, 2011. An investigation into the anti-microbial and anti-fungal properties of earthworm powder obtained from Eisenia fetida. Am. J. Food Technol., 6: 329-335.
CrossRefDirect Link - Ansari, A.A. and P. Saywack, 2011. Identification and classification of earthworm species in Guyana. Int. J. Zool. Res., 7: 93-99.
CrossRefDirect Link - Ansari, A.A., 2011. Worm powered environmental biotechnology in organic waste management. Int. J. Soil Sci., 6: 25-30.
CrossRefDirect Link - Bajsa, O., J. Nair, K. Mathew and G.E. Ho, 2003. Vermiculture as a tool for domestic wastewater management. Water Sci. Technol., 48: 125-132.
PubMedDirect Link - Bansal, S. and K.K. Kapoor, 2000. Vermicomposting of crop residues and cattle dung with Eisenia foetida. Bioresour. Technol., 73: 95-98.
CrossRefDirect Link - Bohlen, P.J. and C.A. Edwards, 1995. Earthworm effects on N dynamics and soil respiration in microcosms receiving organic and inorganic nutrients. Soil Biol. Biochem., 27: 341-348.
CrossRef - Canellas, L.P., F.L. Olivares, A.L. Okorokova-Facanha and A.R. Facanha, 2002. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence and plasma membrane H+-ATPase activity in maize roots. Plant Physiol., 130: 1951-1957.
CrossRefDirect Link - Chhotaray, D., C.S.K. Mishra and P.K. Mohapatra, 2011. Diversity of bacteria and fungi in the gut and cast of the tropical earthworm Glyphodrillus tuberosus isolated from conventional and organic rice fields. J. Pharmacol. Toxicol., 6: 303-311.
CrossRefDirect Link - Elviror, C., L. Sampedro, J. Dominguez and S. Mato, 1997. Vermicomposting of wastewater sludge from paper pulp industry with nitrogen rich materials. Soil Biol. Biochem., 29: 759-762.
CrossRef - Kumar, A.G. and G. Sekaran, 2005. Enteric pathogen modification by anaecic earthworm Lampito mauritii. J. Applied Sci. Environ., 9: 15-17.
Direct Link - Idowu, A.B., M.O. Edema and A.O. Adeyi, 2006. Distribution of bacteria and fungi in the earthworm Libyodrillus violaceous (Annelida: Oligochaeta), a native earthworm from Nigeria. Rev. Biol. Trop., 54: 49-58.
PubMed - Maboeta, M.S. and L. von Rensburg, 2003. Vermicomposting of industrially produced woodchips and sewage sludge utilizing Eisenia fetida. Ecotoxicol. Environ. Safety, 56: 265-270.
Direct Link - Masciandaro, G., B. Ceccanti and C. Garcia, 2000. In situ vermicomposting of biological sludges and impacts on soil quality. Soil Biol. Biochem., 32: 1015-1024.
CrossRef - Omrani, G.A., M. Zamanzadeh, A. Maleki and Y. Ashori, 2005. Earthworm ecology in the Northern part of Iran: With an emphasis on compost worm Eisenia fetida. J. Applied Sci., 5: 1434-1437.
CrossRefDirect Link - Parthasarathi, K. and L.S. Ranganathan, 1999. Longevity of microbial and enzyme activity and their influence on NPK content in pressmud vermicasts. Eur. J. Soil Biol., 35: 107-113.
CrossRef - Pearson, R.W., 1952. Potassium supplying power of eight alabama soils. Soil Sci., 74: 301-310.
Direct Link - Pierre, V., R. Phillip, L. Margnerite and C. Pierrette, 1982. Anti-bacterial activity of the haemolytic system from the earthworms Elsinia foetida andrei. J. Invertebr. Pathol., 40: 21-27.
CrossRef - Rizl, V. and A. Novokova, 2003. Interactions between micro-fungi and Eisenia andrei (Oligochaeta) during cattle manure vermicomposting the 7th international symposium on earthworm ecology Cardiff Wales. Pedobiologia, 47: 895-899.
CrossRef - Sidhu, J., R.A. Gibbs, G.E. Ho and I. Unkovich, 2001. The role of indigenous microorganisms in suppression of Salmonella regrowth in composted biosolids. Water Res., 35: 913-920.
CrossRef - Singleton, D.R., P.F. Hendrix, D.C. Coleman and W.B. Whitman, 2003. Identification of uncultured bacteria tightly associated with the intestine of the earthworm Lumbricus rubellus (Lumbricidae; Oligochaeta). Soil Biol. Biochem., 35: 1547-1555.
CrossRefDirect Link - Sinha, R.K., K. Chauhan, D. Valani, V. Chandran, B.K. Soni and V. Patel, 2010. Earthworms: Charles Darwin's unheralded soldiers of mankind: Protective and productive for man and environment. J. Environ. Protect., 1: 251-260.
CrossRefDirect Link - Sinha, R.K., S. Heart, G. Bharambe and A. Brahambhatt, 2009. Vermistabilization of sewage sludge (biosolids) by earthworms converting a potential biohazard destined for landfill disposal into a pathogen free nutritive and save biofertilizer for farms. Waste Manage. Res., 28: 872-881.
CrossRefDirect Link - Tangjang, S., K. Arunachalam, A. Arunachalam and A.K. Shukla, 2009. Microbial population dynamics of soil under traditional agroforestry systems in Northeast India. Res. J. Soil Biol., 1: 1-7.
CrossRefDirect Link - Verma, D. and Shweta, 2011. Earthworm resources of western himalayan region, India. Int. J. Soil Sci., 6: 124-133.
CrossRef - Shakir, S.H.S. and R.W. Weaver, 2002. Earthworm survival in oil contaminated soil. Plant Soil, 240: 127-132.
CrossRef