
Rizal Subahar
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
LiveDNA: 62.35053
Adib Kamil Putra Kadarusman
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Fatmawaty
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Nurhuda Sahar
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Rawina Winita
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Lisawati Susanto
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Yulhasri
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Nadar S. Lubis
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Mulyati
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Nurhadi Eko Firmansyah
Department of Parasitology, Faculty of Medicine, University of Indonesia, Jl. Salemba Raya 6, Jakarta 10430, Indonesia
Journal of Entomology
Year: 2021 | Volume: 18 | Issue: 1 | Page No.: 19-28







ABSTRACT
Background and Objective: Mentha piperita (peppermint) contains terpenoids (essential oils) that can destroy the digestive tracts and nervous systems of insects. The objective of the study was to evaluate the effect of low concentrations of the crude leaf extract of M. piperita on C. quinquefasciatus larvae through histopathological midgut changes and decreased immunoreactivity of octopamine and tyramine. Materials and Methods: The phytochemical analysis of the extract was performed using Gas Chromatography-Mass Spectrometry (GC-MS). Culex quinquefasciatus larvae were exposed to different concentrations of the extract. Histopathological changes in the midguts of the larvae were tested by histopathological examination and the immunoreactivity of octopamine and tyramine was measured using an immunohistochemical method. Results: Terpenoids were major components of the M. piperita crude leaf extract. At 24 hrs, the LC50 and LC90 values were 2.56 and 6.64 ppm, respectively. The extract caused several histopathological midgut changes, including fragmented food boluses, deformed epithelial cells, disintegrated epithelial layers, damaged microvilli and broken peritrophic membranes. Octopamine and tyramine were detected in the midgut, but their immunoreactivity had decreased. Conclusion: Mentha piperita has the potential to eradicate the population of C. quinquefasciatus as an alternative insecticide.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Rizal Subahar, Adib Kamil Putra Kadarusman, Fatmawaty, Nurhuda Sahar, Rawina Winita, Lisawati Susanto, Yulhasri, Nadar S. Lubis, Mulyati and Nurhadi Eko Firmansyah, 2021. Toxic Effects of Mentha piperita Extract on Culex quinquefasciatus Larvae (Diptera: Culicidae). Journal of Entomology, 18: 19-28.
DOI: 10.3923/je.2021.19.28
URL: https://scialert.net/abstract/?doi=je.2021.19.28
DOI: 10.3923/je.2021.19.28
URL: https://scialert.net/abstract/?doi=je.2021.19.28
INTRODUCTION
Vector-borne Diseases (VBDs) are serious public health problems across the world, accounting for more than 17% of all infectious diseases and more than 700,000 deaths annually. The causes of VBDs include parasites, bacteria and viruses1,2. Culex quinquefasciatus is a southern house mosquito that transmits VBDs such as West Nile virus, St. Louis encephalitis, Japanese encephalitis, Rift Valley fever and lymphatic Bancroftian filariasis3,4. Thus, mosquito control programs play an important role in inhibiting the transmission of VBDs5-7.
Recently, C. quinquefasciatus mosquitoes have become resistant to insecticides in many countries8-10. Some researchers have demonstrated that insecticide-resistant C. quinquefasciatus genes exhibit point mutations. For example, an L1014F kdr mutation in the Voltage-gated Sodium Channel (VGSC) gene has been found to confer resistance to pyrethroid and DDT, while a G1195 mutation in the ace-1 gene has been found to confer resistance to temephos and malathion9. One of the factors contributing to insecticide resistance in C. quinquefasciatus is the frequent use of insecticides to control mosquito populations10. Thus, alternative insecticides obtained from plant bioactive compounds are needed to prevent insecticide resistance in mosquitoes.
Mentha piperita L. (peppermint) belongs to the Lamiaceae family and has menthol (monoterpene) as a major bioactive compound11,12. Menthol is a type of terpenoid12. A previous study demonstrated that M. piperita has toxic effects on C. quinquefasciatus larvae due to its terpenoid content13. Terpenoids comprise a type of essential oil that has toxic effects on insects via contact, ingestion and fumigation by acting on the nervous central system, acetylcholine, γ-aminobutyric acid, the octopaminergic system and the respiratory system12,14.
Most previous studies of M. piperita leaf extract have used high concentrations of extract (~80 ppm) with LC50 values ranging from 26.19-111.9 ppm against Aedes aegypti larvae for 48-72 hrs13,15,16. However, these studies did not investigate the histopathological changes in the midguts of C. quinquefasciatus larvae caused by M. piperita leaf extract. The present study investigated how low concentrations of this extract (0.05-1 ppm) can kill C. quinquefasciatus larvae. For example, the extract can damage the digestive tracts of C. quinquefasciatus larvae through histopathological changes in the midgut. Furthermore, the extract can damage components of the nervous system, particularly neurotransmitters, octopamine and tyramine.
A previous study demonstrated that terpenoids (essential oils) can modify the octopaminergic system by competing with octopamine receptors17,18. In contrast, present study detected octopamine and tyramine in the midguts of C. quinquefasciatus larvae exposed to M. piperita leaf extract within 24 hrs. The aim of the present study was to evaluate how low concentrations of the crude leaf extract of M. piperita can kill C. quinquefasciatus larvae through histopathological changes in the midgut and decreased immunoreactivity of octopamine and tyramine.
MATERIALS AND METHODS
Study area: The study was conducted in April, 2019-2021. Larvae of C. quinquefasctiatus were collected in Jakarta city, the capital city of Indonesia. The locations of the larval collection consisted of Jatinegara Sub district, East Jakarta.
Mentha piperita crude leaf extraction: Mentha piperita leaves were obtained from a traditional market in East Jakarta, Indonesia. The leaves were cleaned with tap water, cut into small pieces and dried for three weeks at room temperature. Then, the pieces were blended and filtered separately to produce powder samples. Fifty grams of each filtered powder sample were added to an Erlenmeyer tube (500 mL) with 300 mL of absolute methanol and the tubes were kept at room temperature for three days. Next, the methanolic extracts were filtered using filter paper and the sediments were discarded. Finally, the supernatants were evaporated to remove methanol using a vacuum evaporator. The resulting crude extracts were used throughout the entire study.
Gas chromatography-mass spectrometry (GC-MS) analysis: To determine the chemical compounds present in the methanolic crude leaf extract of M. piperita, a phytochemical screening was conducted using gas chromatography-mass spectrometry (GC-MS; Agilent Technologies, 6890N Network GC System, made in the USA). The procedure and interpretation of the GC-MS analysis were conducted in accordance with the guidelines provided by Agilent Technologies19. The GC-MS analysis was conducted at the Forensic Laboratory Centre of Indonesian National Police Headquarters in Jakarta, Indonesia20.
Bioassay of C. quinquefasciatus larvae: A larval bioassay procedure was conducted as described previously21. The bioassay was conducted using C. quinquefasciatus fourth instar larvae collected from several locations in Jakarta, Indonesia. There were four different concentrations of each extract (0.05, 0.2, 0.5, 0.7 and 1 ppm) modified from a previous study13. The larvae were exposed to each of these concentrations separately. In the treatment group, 25 C. quinquefasciatus third and fourth instar larvae were added to each plastic cup (200 mL in volume) containing 100 mL of the extract. In the control group, 25 C. quinquefasciatus larvae were added to each plastic cup (200 mL in volume) containing tap water. There were four replicates for each treatment group and the bioassay was conducted within 24 hrs to determine the larval mortality rate.
Histopathological examination of the larval midgut: The present study used a routine histopathological technique previously described by de Lemos et al.22. In this procedure, C. quinquefasciatus larvae were exposed to the M. piperita leaf extract for 24 hrs. All the specimens were fixated with 10% formalin. The dehydration of each specimen was performed using a series of increasing alcohol concentrations (70, 80, 90, 95 and 100%). Then, the specimens were embedded in xylene 1, xylene 2 and xylene 3 solutions and a paraffin block. The resulting blocks were cut (5 μm each) using a manual microtome (Model 320, No. 17664, New York, USA) and feather microtome blades (Feather, S35, Japan). Finally, the sections were stained with hematoxylin and eosin (H and E). The stained specimens were carefully observed under a light microscope and images were taken with a digital microscopic mounted camera (Zeiss Axiocam ERC 5s, Germany).
Octopamine and tyramine immunohistochemical staining: An immunohistochemical (IHC) technique was conducted as previously described by Ramos-Vara et al.23. The IHC staining procedure was performed using diagnostic system kits (Abnova, PAB14697 and Cloud-Clone Corp., PAG048GE01). Briefly, deparaffinization was carried out using xylene 1 and xylene 2 (5 min each) and rehydration was carried out with 100, 96 and 80% alcohol followed by rinsing with distilled water. Next, endogenous peroxidase was quenched with 0.3% H2O2 in methanol followed by a tap water wash. The sections were then heated with Tris-EDTA buffer (pH 9.0) antigen retrieval using Retrieval Generation 1 (RG1, BIO GEAR, BGRG-0118) for 15 min, chilled at room temperature (15 min) and embedded in PBS solution (3 min). Afterward, nonspecific binding sites were blocked with a background blocker for 5 min. For tyramine, the sections were incubated with the primary antibody Tyramine Polyclonal Antibody (Abnova, PAB14697) 1:1000 overnight at 4°C. For octopamine, the sections were incubated with the primary antibody Polyclonal Antibody to Octopamine (Cloud-Clone Corp., PAG048Ge01) 1:50 overnight at 4°C and then washed with PBS solution. The sections were then incubated with the secondary antibody PolyVue Plus Mouse/Rabbit Enhancer (ten minutes) at room temperature and washed with PBS solution. Next, the sections were incubated with PolyVue Plus HRP Label (10 min) at room temperature and washed with PBS solution. The sections were treated with chromogen substrate and one drop of DAB mixed with 1 mL of DAB buffer, washed with distilled water, treated with hematoxylin, washed with distilled water (3 min) and then treated with one drop of bluing reagent (10 sec). Next, the sections were dehydrated with 80, 96 and 100% alcohol and treated with xylene 1 and xylene 2 for clearing. Finally, the sections were embedded in Entellan (Merck, 1.07961.0500) under glass cover slips.
Data analysis: Data were expressed as descriptive statistics and analyzed by statistical package for social sciences (SPSS) ver.20. Data on the mortality rate of the dead larvae were tested for normal distribution (Shapiro-Wilk). Data with normal distribution were analyzed by analysis of variance (ANOVA), while data with non-normal distribution were tested by the Kruskal-Wallis test24. The LC50 and LC90 values were performed by probit analysis with a 95% confidence interval; p<0.05 was considered statistically significant25.
RESULTS
Phytochemical analysis of the leaf extract: The results of the GC-MS analysis showed that the crude leaf extract contained 18 phytochemical compounds. Most of these compounds were terpenoids, followed by fatty acids, alkaloids and vitamin E. The first phytochemical compound detected by GC-MS was the alkaloid 3-(3-methyl-3-oxaziridinyl) pyridine and its area peak was 1.18%. The last phytochemical compound detected by GC-MS was cupreol (a terpenoid). Cupreol was present in the highest concentration (20.35%), while 11,13-dimethyl-12-tetradecen-1-ol acetate (a fatty acid) was present in the smallest concentration (Table 1). The terpenoids in the extract included diterpene (neophytadiene, (Z)-1,3 phytadiene, 3,7,11,15-tetramethyl-2-hexadecen-1-ol), monoterpene (camphene) and steroids (stigmastan-3,5-diene, campesterol and cupreol).
Larvicidal activity of the leaf extract: The present study showed that crude leaf extract concentrations ranging from 0.05-1.00 ppm are capable of killing C. quinquefasciatus larvae. Table 2 showed the larvicidal activity exhibited by the crude leaf extract. At 24 hrs, the larval mortality of C. quinquefasciatus induced by the extract ranged from 10.4-20.8%.
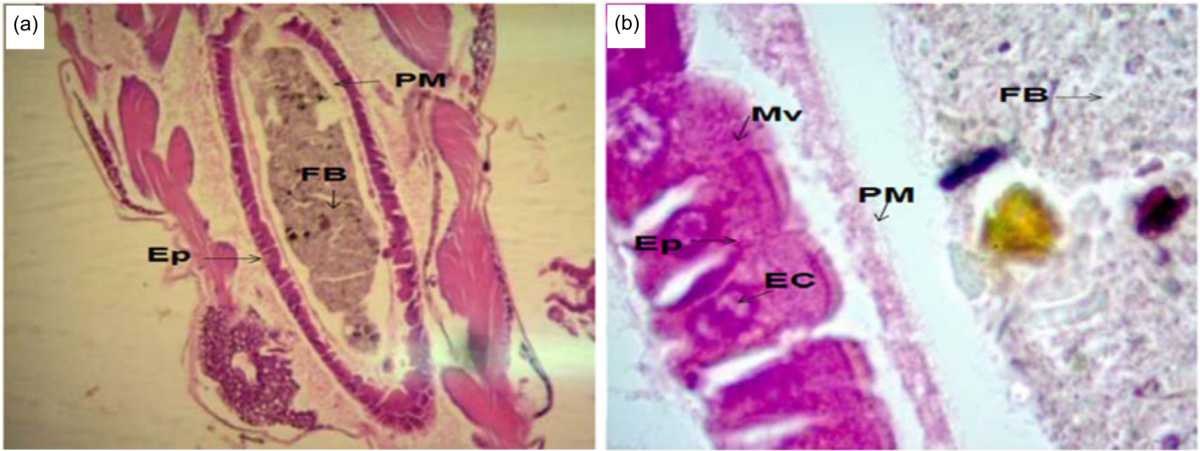 |
| Fig. 1(a-b): | Midgut of a healthy C. quinquefasciatus larva stained with H and E at (a) 10×magnification and (b) 100×magnification FB: Food bolus, PM: Peritrophic membrane, Ep: Epithelial layer, EC: Epithelial cell, Mv: Microvilli, H and E: Hematoxylin and eosin |
| Table 1: GC-MS analysis of the leaf extract | |||||
| Number | Real time | Peak | Constituents | Molecular formula | Group |
| 1 | 7.05 | 1.18 | 3-(3-Methyl-3-oxaziridinyl)pyridine | C7H8N2O | Alkaloid |
| 2 | 9.05 | 10.47 | Neophytadiene | C20H38 | Terpenoid |
| 3 | 9.35 | 2.92 | (Z)-1,3-Phytadiene | C20H38 | Terpenoid |
| 4 | 9.98 | 4.38 | Palmitic acid | C16H32O2 | Fatty acid |
| 5 | 10.85 | 6.02 | 3,7,11,15-Tetramethyl-2-hexadecen-1-ol | C20H40O | Terpenoid |
| 6 | 11.13 | 15.06 | Linolenic acid | C18H30O2 | Fatty acid |
| 7 | 12.68 | 3.49 | Camphene | C10H16 | Terpenoid |
| 8 | 12.81 | 2.03 | Dimethylaminoethyl acrylate | C7H13NO2 | Fatty acid |
| 9 | 12.89 | 1.83 | Methyl 8,11,14-heptadecatrienoate | C18H30O2 | Fatty acid |
| 10 | 13.12 | 4.28 | 2-Palmitoylglycerol | C19H38 O4 | Fatty acid |
| 11 | 14.09 | 6.86 | Methyl 8,11,14-heptadecatrienoate | C18H30O2 | Fatty acid |
| 12 | 14.25 | 6.86 | 11,13-Dimethyl-12-tetradecen-1-ol acetate | C19H38O4 | Fatty acid |
| 13 | 14.44 | 6.66 | Oleamide | C18H35NO | Fatty acid |
| 14 | 16.23 | 0.66 | Stigmastan-3,5-diene | C29H48 | Terpenoid |
| 15 | 16.35 | 4.70 | Vitamin E | C29H50O2 | Vitamin |
| 16 | 17.22 | 2.33 | Campesterol | C28H48O | Terpenoid |
| 17 | 17.46 | 6.20 | Stigmasterol | C29H48O | Terpenoid |
| 18 | 17.97 | 20.35 | Cupreol | C29H50O | Terpenoid |
| Table 2: Larvicidal activity of the leaf extract against C. quinquefasciatus larvae | ||||||
| Concentrations | Culex quinquefasciatus larvae death at 24 hrs | LC50 | LC90 | |||
| (ppm) | N | Death | % | Mean±SD | (95% CI) | (95% CI) |
| 0.05 | 125 | 13 | 10.4 | 2.6±0.9 | 2.56 ppm | 6.64 ppm |
| 0.2 | 125 | 18 | 14.4 | 3.6±1.5 | (1.85-15.831) | (3.451-38.001) |
| 0.5 | 125 | 21 | 16.8 | 4.2±0.8 | ||
| 0.7 | 125 | 23 | 18.4 | 4.6±0.5 | ||
| 1.00 | 125 | 26 | 20.8 | 5.2±0.4 | ||
| N : Number of C. quinquefasciatus larvae tested, LC: Lethal concentration, CI: Confident interval | ||||||
The LC50 and LC90 of the extract were 2.56 and 6.64 ppm, respectively. The mean mortality of the C. quinquefasciatus larvae increased with increased concentrations of the extract.
Histopathological damage to the midguts of C. quinquefasciatus larvae: The healthy C. quinquefasciatus larvae used as the control group showed that the normal midgut consists of a Food Bolus (FB), a Peritrophic Membrane (PM), an Epithelial Layer (Ep), microvilli and Epithelial Cells (ECs). The Ep is provided by the microvilli and ECs. The ECs are located inside the Ep and may include several degenerative cells (Fig. 1a-b).
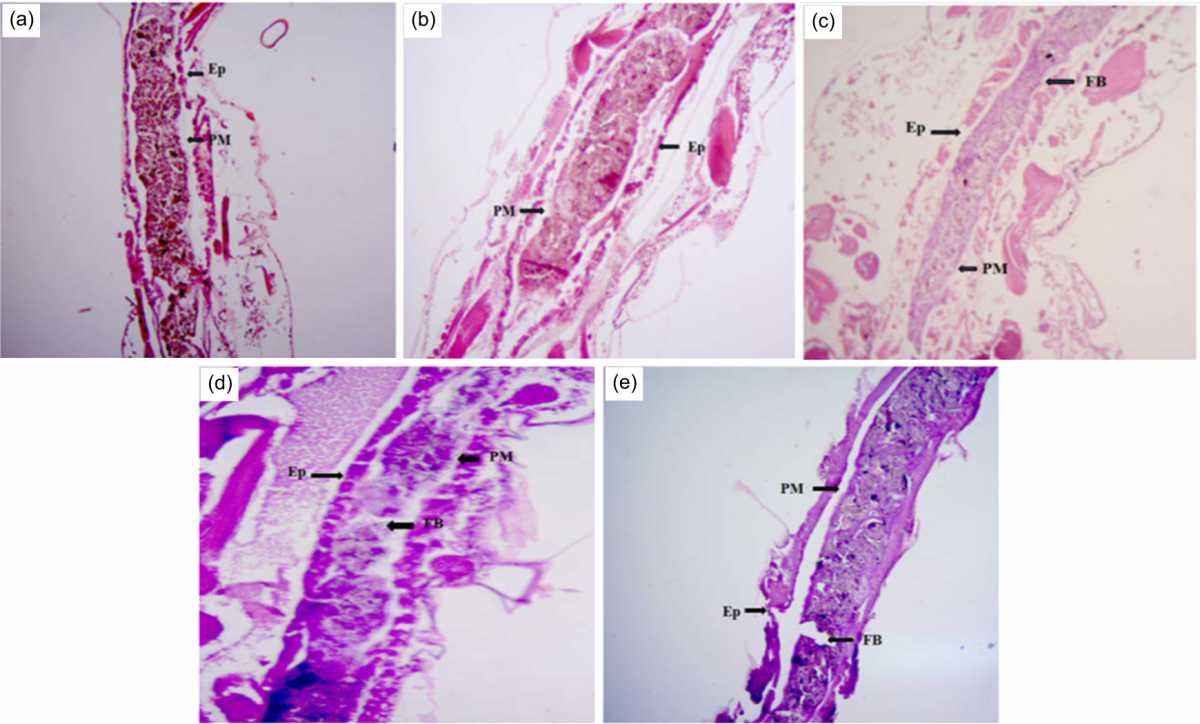 |
| Fig. 2(a-e): | Midguts of the C. quinquefasciatus larvae were histopathologically damaged by the crude leaf extract at 10×magnification. Extract concentrations (a) 0.05 ppm, (b) 0.2 ppm, (c) 0.5 ppm, (d) 0.7 ppm and (e) 1.00 ppm Arrow indicated the damaged midgut, FB: Food bolus, PM: Peritrophic membrane, Ep: Epithelial layer, EC: Epithelial cell, Mv: Microvilli |
| Table 3: Histopathological damage to the midguts of the C. quinquefasciatus larvae | ||||||
| Midgut parts of C. quinquefasciatus larvae | ||||||
Concentrations | ||||||
| Treatment | (ppm) | FB | PM | Ep | Mv | EC |
| Control | - | + | + | + | + | + |
| Leaf extract | 0.05 | + | - | - | -/+ | - |
0.2 | + | - | - | - | - | |
0.5 | - | - | - | - | - | |
0.7 | - | - | - | - | - | |
1.00 | - | - | - | - | - | |
| + (positive): No histopathological changes in the midgut, - (negative): Histopathological changes in the midgut, FB: Food bolus, PM: Peritrophic membrane, Ep: Epithelial layer, Mv: Microvilli, EC: Epithelial cell | ||||||
The present study showed that treatment with M. piperita crude leaf extract causes damage to the midguts of C. quinquefasciatus larvae. At 24 hrs, an extract concentration of 0.05 ppm damaged the Ep of the midgut (Fig. 2a). At an extract concentration of 0.2 ppm, the Ep disintegrated, the ECs ruptured and the microvilli disappeared (Fig. 2b). At extract concentrations of 0.5 ppm (Fig. 2c), 0.7 ppm (Fig. 2d) and 1 ppm (Fig. 2e), all the midgut parts (the FB, PM, Ep, microvilli and ECs) ruptured.
Table 3 summarizes the histopathological damage to the midguts of the C. quinquefasciatus larvae caused by the leaf extract. The larvae in the control group were found to have normal midguts, whereas the group treated with the extract showed damage in all parts of the midgut. Increased concentrations of the extract caused increased damage to the midgut.
Detection of octopamine and tyramine in the midgut: The present study used the healthy C. quinquefasciatus larvae as the control group to evaluate octopamine and tyramine in the midgut of the larvae. Octopamine and tyramine were successfully detected in the midguts of the control larvae. In FB of midgut, octopamine is dark brown in color under the light microscopy, 10× magnifications (Fig. 3a). In Ep of the midgut, octopamine is light brown (small dots) in color (Fig. 3b). In EC of the EP, octopamine is light brown (small dots) under light microscopy, 100 x magnifications (Fig. 3b). In contrast, tyramine is light brown (small dots) only or very light brown in color under the light microscopy, 10×magnifications (Fig. 4a). Sometimes, tyramine is very light brown in color found in FB, Ep and EC using the light microscopy, 100×magnifications (Fig. 4b).
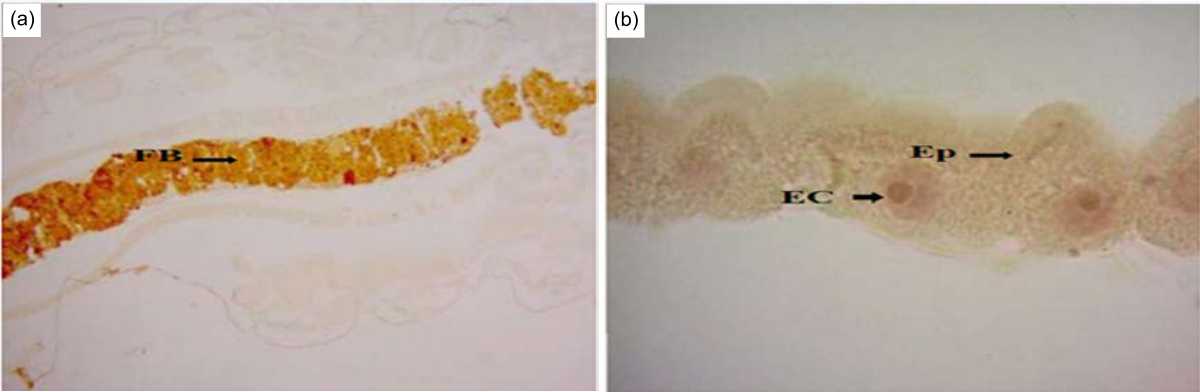 |
| Fig. 3(a-b): | Midgut of a C. quinquefasciatus larva (control group: where octopamine were detected using an IHC method) at (a) 10×magnification and (b) 100×magnification Arrow indicates the locations of positive (+) octopamine |
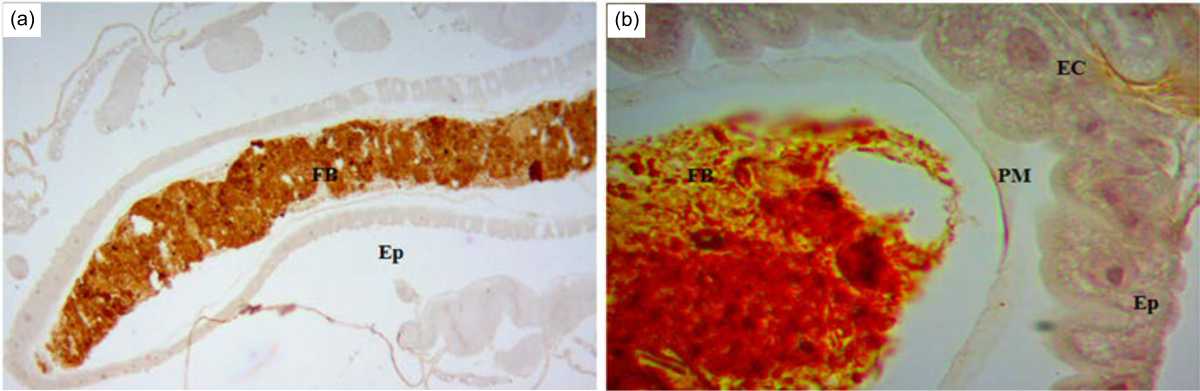 |
| Fig. 4(a-b): | Midgut of a C. quinquefasciatus larva (control group: where tyramine were detected using an IHC method) at (a) 10×magnification and (b) 100×magnification Arrow indicates the locations of positive (+) octopamine |
| Table 4: Immuno reactivity of tyramine in the midguts of C. quinquefasciatus larvae | ||||||
| Tyramine in the midgut | ||||||
Concentrations | ||||||
| Treatment | (ppm) | FB | PM | Ep | Mv | EC |
| Control | - | +++ | - | ++ | - | ++ |
| Leaf extract | 0.05 | + | - | + | - | + |
0.2 | + | - | + | - | - | |
0.5 | + | - | + | - | + | |
0.7 | + | - | - | - | - | |
1 | + | - | - | - | - | |
| + (positive): Tyramine was detected in the midgut, - (negative): Tyramine was not detected in the midgut, FB: Food bolus, PM: Peritrophic membrane, Ep: Epithelial layer, Mv: Microvilli, EC: Epithelial cell | ||||||
The study indicated that octopamine and tyramine were strong immune reactivity in the midgut of the healthy larvae.
Table 4 showed that tyramine was detected by the Polyclonal Antibody to Tyramine using the IHC method. In the control group, tyramine was found in the FB, Ep and ECs. Stronger tyramine immunoreactivity was found in the FB of the midgut than in the Ep and ECs. In contrast, the FB, Ep and ECs showed weak tyramine immunoreactivity after 24 hrs of exposure to the extract. At extract concentrations of 0.7 and 1 ppm, tyramine could not be detected.
In the control group, octopamine was detected in the FB, Ep and ECs. The FB exhibited stronger octopamine immunoreactivity in the midgut than the Ep and ECs.
| Table 5: Immuno reactivity of octopamine in the midguts of C. quinquefasciatus larvae | ||||||
| Octopamine in the midgut | ||||||
Concentrations | ||||||
| Treatment | (ppm) | FB | PM | Ep | Mv | EC |
| Control | - | ++++ | - | +++ | - | +++ |
| Leaf extract | 0.05 | ++ | - | ++ | - | + |
0.2 | ++ | - | + | - | + | |
0.5 | ++ | - | + | - | + | |
0.7 | ++ | - | - | - | - | |
1 | ++ | - | - | - | - | |
| + (positive): Octopamine was detected in the midgut, - (negative): Octopamine was not detected in the midgut, FB: Food bolus, PM: Peritrophic membrane, Ep: Epithelial layer, Mv: Microvilli, EC: Epithelial cell | ||||||
In contrast, the FB, Ep and ECs showed weak octopamine immunoreactivity after 24 hrs of exposure to the extract. At extract concentrations of 0.7 and 1 ppm, octopamine could not be detected (Table 5).
DISCUSSION
The present study employed GC-MS to determine the chemical constituents of M. piperita leaf extracts. Most of these extracts contained terpenoids as major compounds, including diterpene (neophytadiene, (Z)-1,3 phytadene and 3,7,11,15-tetramethyl-2-hexadecen-1-ol), monoterpene (camphene) and steroids (stigmastan-3,5-diene, campesterol and cupreol). In contrast, another study found that menthol (monoterpene) is a major component of M. piperita26-28. In the present study, a vacuum evaporator was used to remove methanol absolute solution, so that there would be no menthol in the extract. Additionally, steroids (particularly cupreol) were found to be major components of the extract. Current findings were consistent with those of Tong29, who suggested that steroids are a type of terpenoid and that terpenoids, overall are a major component of M. piperita27,28.
In the present study, leaf extract concentrations ranging from 0.05-1.00 ppm exhibited larvicidal activity against C. quinquefasciatus larvae. The present study used lower concentrations of extract than those used in previous studies. The larval mortality produced by the extract ranged from 10.4-20.8% and the LC50 and LC90 of the extract were 2.56 and 6.64 ppm, respectively. These findings, along with the findings of previous studies13,15, supported the claim that M. piperita leaf extract is an efficient larvicidal against mosquito larvae.
Kumar et al.15 reported that an essential oil extracted from M. piperita leaves was an efficient larvicide and repellent against the dengue vector Ae. aegypti, with LC50 and LC90 values of 111.9 and 295.18 ppm at 24 hrs of exposure, respectively. Similarly, Kalaivani et al.13 revealed that an oil extract obtained from M. piperita was highly toxic against A. aegypti larvae, with an LC50 value of 47.54 ppm. Kalaivani et al.13 also found that higher rates of larval mortality occurred at higher concentrations (~80 ppm) of extract within 48 hrs of exposure. Another study showed that the LC50 value of the extract of M. piperita was 26.19 ppm against A. aegypti larvae16. Similarly, the present study showed that higher rates of larval mortality occurred at higher concentrations (~1 ppm) of extract. Dias and Moraes11 categorized any plant extract with an LC50<100 ppm (mg L–1) as an extract containing bioactive compounds. Therefore, the M. piperita leaf extract used in the present study can be considered to contain bioactive compounds.
The present study also showed that M. piperita leaf extract can damage the histopathological midguts of C. quinquefasciatus larvae because the terpenoids in the extract can disrupt cell membranes. This finding was consistent with those of previous studies. Terpenoids are volatile essential oils that penetrate into insects rapidly via the respiratory tract, causing breathing abnormalities that can lead to asphyxiation and, finally, the death of the insect12,28. Additionally, terpenoids have been found to induce morphological changes in the midgut30. In the present study, the leaf extract caused damage and histopathological changes in midgut parts such as the FB, PM, Ep, microvilli and ECs. According to the results of histopathological examinations using H and E staining, all tested concentrations of the extract were capable of damaging any part of the midgut, but the higher concentrations caused more damage than the lower concentrations. The histopathological changes observed in the midgut were due to the hydrophobic nature of terpenoids. Terpenoids can destroy lipid matrices and can thus destroy the lipid bilayer of the cell membrane, leading to increased fluidity and cell lysis30. Therefore, terpenoids contribute to cell membrane disruption and cause damage to the midgut in C. quinquefasciatus larvae.
The present study also supported the role of terpenoids in leaf extract-induced oxidative stress30,31. Sies32 reported that oxidative stress, an imbalance of oxidants and antioxidants, causes histopathology. Yu et al.33 demonstrated that increasing hydrogen peroxide, ion hydroxyl, superoxidate and malondialdehyde (MDA) levels leads to histopathology in the midgut structures of Bombyx mori. Furthermore, Isah et al.30 reported that sesquiterpenoids cause mitochondrial dysfunction, leading to increased Reactive Oxygen Species (ROS) production via the electron transport chain and thus causing a loss of cellular integrity and parasite death.
The present study demonstrated that the terpenoids in M. piperita leaf extract cause low immunoreactivity of the octopamine and tyramine in the midguts of C. quinquefasciatus larvae. After 24 hrs of exposure to the leaf extract octopamine and tyramine could be detected in parts of the midgut including the FB, Ep and ECs. In contrast, the immunoreactivity of the octopamine and tyramine in the midguts of the control larvae was strong, while that in the midguts of the larvae exposed to the extract was weak. Therefore, it is possible that the M. piperita leaf extract targets neural cells, particularly neurotransmitters (e.g., octopamine and tyramine).
The present findings concerning low immunoreactivity of the octopamine and tyramine in the midguts of C. quinquefasciatus larvae were consistent with the theoretical contributions of previous studies. Terpenoids act on the octopaminergic systems of insects17. Essential oils from aromatic plants have been found to cause significant increases in both cyclic AMP and calcium levels17,18. Terpenoid essential oils can modify the neuronal activity of octopamine receptors because they compete with octopamine by binding to its receptors, which belong to the G-protein receptor family18,34. Additionally, terpenoid essential oils have a nervous cell target octopamine (a neurotransmitter) through which they can cause physiological modulations in insects12. Octopamine and tyramine are known as physiological behavior markers in insects, as these neurotransmitters play important roles in feeding, flying, growth and development34,35. Therefore, it appears that the M. piperita leaf extract leads to physiological modulations and, finally, death in C. quinquefasciatus larvae by targeting the nervous system.
The present study supported the usefulness of M. piperita leaf extract in reducing environmental pollution and preventing the development of insecticide resistance in C. quinquefasciatus and other mosquitoes. Culex quinquefasciatus, known as vector of VBDs is dangerous to human life. Many people live in the endemic areas of VBDs1,2 and C. quinquefasciatus has become resistant to insecticides in many countries10. Therefore, the findings of the present study will aid in reducing the application of synthetic insecticides. Essential oils act at multiple levels in insects, so the probability that insects will generate resistance to these compounds is low36,37. Iwuagwu et al.38 suggested that the use of plant-derived bioactive compounds is an eco-friendly way to control various vectors of disease.
CONCLUSION
Mentha piperita leaf extract was found to contain terpenoids, fatty acids, alkaloids and vitamin E. Low concentrations of the extract exhibited larvicidal activity against C. quinquefasciatus larvae. Terpenoids, which are major components of this extract, have targets of toxicity in the digestive tract (the midgut) and the nervous system (octopamine and tyramine). Therefore, M. piperita leaf extract could serve as an alternative insecticide to control the C. quinquefasciatus population.
SIGNIFICANCE STATEMENT
Low concentrations of M. piperita leaf extract showed larvicidal activity against C. quinquefasciatus larvae. Terpenoids, which are major components of this extract, have targets of toxicity in the digestive tract (the midgut) and the nervous system (octopamine and tyramine). Thus, M. piperita leaf extract could serve as an alternative insecticide to control the C. quinquefasciatus population.
ACKNOWLEDGMENTS
This study was financially supported by the University of Indonesia (Indonesia, HIBA Pengabdian Masyarakat DRPM UI, 2019). We wish to thank all of those who helped in the making of this study.
REFERENCES
- Müller, R., F. Reuss, V. Kendrovski and D. Montag, 2019. Vector-Borne Diseases. In: Biodiversity and Health in the Face of Climate Change, Marselle, M.R., J. Stadler, H. Korn, K.N. Irvine and A. Bonn (Eds)., Springer International Publishing, Switzerland, ISBN: 978-3-030-02318-8 pp: 67-90.
CrossRefDirect Link - Ramirez, B., 2017. Support for research towards understanding the population health vulnerabilities to vector-borne diseases: Increasing resilience under climate change conditions in Africa. Infect. Dis. Poverty, Vol. 6.
CrossRefDirect Link - Soh, S. and J. Aik, 2021. The abundance of Culex mosquito vectors for West Nile Virus and other flaviviruses: A time-series analysis of rainfall and temperature dependence in Singapore. Sci. Total Environ., Vol. 754.
CrossRefDirect Link - Negi, C., P. Verma, 2018. Review on Culex quinquefasciatus: Southern house mosquito. Int. J. Life Sci. Scienti. Res., 4: 1563-1566.
Direct Link - Impoinvil, D.E., S. Ahmad, A. Troyo, J. Keating and A.K. Githeko et al., 2007. Comparison of mosquito control programs in seven urban sites in Africa, the Middle East and the Americas. Health Policy, 83: 196-212.
CrossRefDirect Link - Hierlihy, C., L. Waddell, I. Young, J. Greig, T. Corrin and M. Mascarenhas, 2019. A systematic review of individual and community mitigation measures for prevention and control of chikungunya virus. PLoS ONE, Vol. 14.
CrossRefDirect Link - Antonio, C.A.T., A.N.C. Bermudez, K.L. Cochon, M.S.G.L. Reyes and C.D.H. Torres et al., 2020. Recommendations for intersectoral collaboration for the prevention and control of vector-borne diseases: Results from a modified delphi process. J. Infect. Dis., 222: S726-S731.
CrossRefDirect Link - Moyes, C.L., J. Vontas, A.J. Martins, L.C. Ng and S.Y. Koou et al., 2017. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis.
CrossRefDirect Link - Delannay, C., D. Goindin, K. Kellaou, C. Ramdini, J. Gustave and A. Vega-Rúa, 2018. Multiple insecticide resistance in Culex quinquefasciatus populations from Guadeloupe (French West Indies) and associated mechanisms. PLoS ONE, Vol. 13.
CrossRefDirect Link - Rai, P., M. Bharati, A. Subba and D. Saha, 2019. Insecticide resistance mapping in the vector of lymphatic filariasis, Culex quinquefasciatus Say from northern region of West Bengal, India. PLoS ONE, Vol. 14.
CrossRefDirect Link - Dias, C.N. and D.F.C. Moraes, 2014. Essential oils and their compounds as Aedes aegypti L. (Diptera: Culicidae) larvicides: Review. Parasitol. Res., 113: 565-592.
CrossRefDirect Link - Singh, P. and A.K. Pandey, 2018. Prospective of essential oil of the genus Mentha as biopesticide: A review. Front. Plant. Sci.
CrossRefDirect Link - Kalaivani, K., S. Senthil-Nathan and A.G. Murugesan, 2012. Biological activity of selected Lamiaceae and Zingiberaceae plant essential oils against the dengue vector Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res., 110: 1261-1268.
CrossRef - Plata-Rueda, A., G. Da Silva Rolim, C.F. Wilcken, J.C. Zanuncio, J.E. Serrão and L.C. Martínez, 2020. Acute toxicity and sublethal effects of lemongrass essential oil and their components against the granary weevil, Sitophilus granarius. Insects, Vol. 11.
CrossRefDirect Link - Kumar, S., N. Wahab and R. Warikoo, 2011. Bioefficacy of Mentha piperita essential oil against dengue fever mosquito Aedes aegypti L. Asian Pacific J. Trop. Biomed., 1: 85-88.
CrossRefDirect Link - Pathak, N., P.K. Mittal, O.P. Singh, D.V. Sagar and P. Vasudepan, 2000. Larvicidal action of essential oils from plants against the vector mosquitoes Anopheles stephensi (Liston), Culex quinquefsaciatus (Say) and Aedes aegypti (L). Int. Pest. Control, 42: 53-55.
Direct Link - Kostyukovsky, M., A. Rafaeli, C. Gileadi, N. Demchenko and E. Shaaya, 2002. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manage. Sci., 58: 1101-1106.
CrossRefPubMedDirect Link - Jankowska, M., J. Rogalska, J. Wyszkowska and M. Stankiewicz, 2018. Molecular targets for components of essential oils in the insect nervous system-A review. Molecules, Vol. 23.
CrossRefDirect Link - Shahzad, R., A.L. Khan, M. Waqas, I. Ullah and S. Bilal et al., 2019. Metabolic and proteomic alteration in phytohormone-producing endophytic Bacillus amyloliquefaciens RWL-1 during methanol utilization. Metabolomics, Vol. 15.
CrossRefDirect Link - Subahar, R., A. Achmadsyah, M. Yasmine, L. Susanto, Fatmawaty and R. Winita, 2019. Plant essential oils enhanced the percent mortality of gravid Aedes aegypti (Diptera: Culicidae) mosquitoes, dengue vector. Int. J. Entomol. Res., 4: 15-20.
Direct Link - Sharma, G., K. Kumar, A. Sharma and V. Agrawal, 2012. Bioassay of Artemisia annua leaf extracts and artemisinin against larvae of Culex quinquefasciatus and Culex tritaeniorhynchus. J. Am. Mosq. Control Assoc., 28: 317-319.
CrossRefPubMedDirect Link - Lemos, A., F. Adam, K. Moura, L. Moraes and O. Silva, 2018. Histological and histochemical characterization of the midgut of healthy aedes aegypti larvae. Annu. Res. Rev. Biol., 22: 1-15.
CrossRefDirect Link - Ramos-Vara, J.A., 2017. Principles and Methods of Immunohistochemistry. In: Drug Safety Evaluation, Gautier, J.C. (Ed.)., Springer, Humana Press, New York, ISBN: 978-1-4939-7172-5 pp: 115-128.
CrossRefDirect Link - Fan, C., D. Zhang and C.H. Zhang, 2011. On sample size of the kruskal-wallis test with application to a mouse peritoneal cavity study. Biometrics, 67: 213-224.
CrossRefDirect Link - Huang, Y.T. and T. Cai, 2016. Mediation analysis for survival data using semiparametric probit models. Biometrics, 72: 563-574.
CrossRefDirect Link - Kalemba, D. and A. Synowiec, 2019. Agrobiological interactions of essential oils of two menthol mints: Mentha piperita and Mentha arvensis. Molecules, Vol. 25.
CrossRefDirect Link - Samber, N., A. Khan, A. Varma and N. Manzoor, 2015. Synergistic anti-candidal activity and mode of action of Mentha piperita essential oil and its major components. Pharm. Biol., 53: 1496-1504.
CrossRefDirect Link - Saeidi, K. and S. Mirfakhraie, 2017. Chemical composition and insecticidal activity Mentha piperita L. essential oil against the cowpea seed beetle Callosobruchus maculatus F. (Coleoptera: Bruchidae). J. Entomol. Acarol. Res., 49: 127-134.
CrossRefDirect Link - Tong, W.Y., 2013. Biotransformation of Terpenoids and Steroids. In: Natural Products, Ramawat, K. and J.M. Mérillon (Eds.), Springer Berlin Heidelberg, New York, ISBN: 978-3-642-22144-6, pp: 2733-2759.
CrossRefDirect Link - Isah, M.B., N. Tajuddeen, M.I. Umar, Z.A. Alhafiz, A. Mohammed and M.A. Ibrahim, 2018. Terpenoids as emerging therapeutic agents: cellular targets and mechanisms of action against protozoan parasites. Stud. Nat. Prod. Chem., 59: 227-250.
CrossRefDirect Link - Singh, R., M.A. Shushni and A. Belkheir, 2015. Antibacterial and antioxidant activities of Mentha piperita L. Arabian J. Chem., 8: 322-328.
CrossRefDirect Link - Sies, H., 1997. Oxidative stress: Oxidants and antioxidants. Exp. Physiol., 82: 291-295.
CrossRefPubMedDirect Link - Yu, X., Q. Sun, B. Li, Y. Xie and X. Zhao et al., 2015. Mechanisms of larval midgut damage following exposure to phoxim and repair of phoxim-induced damage by cerium in Bombyx mori. Environ. Toxicol., 30: 452-460.
CrossRefDirect Link - Roeder, T., 2005. Tyramine and octopamine: Ruling behavior and metabolism. Annu. Rev. Entomol., 50: 447-477.
CrossRefDirect Link - Damrau, C., N. Toshima, T. Tanimura, B. Brembs and J. Colomb, 2018. Octopamine and tyramine contribute separately to the counter-regulatory response to sugar deficit in Drosophila. Front. Syst. Neurosci., Vol. 11.
CrossRefDirect Link - Maddala, V.K.S., 2019. Green pest management practices for sustainable buildings: Critical review. Sci. Prog., 102: 141-152.
CrossRefDirect Link - Saeidi, K. and B. Hassanpour, 2014. Efficiency of Mentha piperita L. and Mentha pulegium L. essential oils on nutritional indices of Plodia interpunctella Hübner (Lepidoptera: Pyralidae). J. Entomol. Acarol. Res., 46: 13-17.
CrossRefDirect Link - Iwuagwu, M.O., P.E. Etusim, N.C. Emmanuel, J.C. Igwe, V.O. Nwaugo and R.A. Onyeagba, 2020. Exploitation of plant herbs in the control of disease vectors: A review. Pharm. Biosci. J., 8: 7-21.
CrossRefDirect Link