
Ghulam Ali Bugti
Department of Entomology, Lasbela University of Agriculture, Water and Marine Sciences, Uthal, Balochistan, Pakistan
LiveDNA: 92.29689
Wang Bin
Anhui Provincial Key Laboratory of Microbial Control, Anhui Agricultural University, People Republic of China
Shafique Ahmed Memon
Department of Entomology, Lasbela University of Agriculture, Water and Marine Sciences, Uthal, Balochistan, Pakistan
Ghulam Khaliq
Department of Horticulture, Lasbela University of Agriculture, Water and Marine Sciences, Uthal, Balochistan, Pakistan
Muhammad Abuzar Jaffar
Department of Horticulture, Lasbela University of Agriculture, Water and Marine Sciences, Uthal, Balochistan, Pakistan
Journal of Entomology
Year: 2020 | Volume: 17 | Issue: 2 | Page No.: 74-83







ABSTRACT
Entomopathogenic fungi have great potential to control insect pests, which either feed on aerial or underground plant parts. The study revealed that virulence and pathogenicity of fungi greatly affected by biotic factors such as fungal species, spore density, host plant chemistry, insect host species and host life stages and abiotic factors likewise, temperature, sunlight, rain and relative humidity. The aim of this review is to find out the factors and characteristics which affect the virulence and pathogenicity of the fungi. From the literature, it was concluded that the relative humidity seems to be more important than other factors to ensure infection and successful disease development in targeted insect population.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ghulam Ali Bugti, Wang Bin, Shafique Ahmed Memon, Ghulam Khaliq and Muhammad Abuzar Jaffar, 2020. Entomopathogenic Fungi: Factors Involved in Successful Microbial Control of Insect Pests. Journal of Entomology, 17: 74-83.
DOI: 10.3923/je.2020.74.83
URL: https://scialert.net/abstract/?doi=je.2020.74.83
DOI: 10.3923/je.2020.74.83
URL: https://scialert.net/abstract/?doi=je.2020.74.83
INTRODUCTION
Approximately one thousand insect pathogenic fungal species have been known to infect various insects species1. About 100 conidia based insecticides are registered worldwide to control many serious insect pests of agriculture2. These fungi can produce different enzymes and secondary metabolites which help insect infection development and also play and vital insecticidal role3. As insect cuticle contains up to 70% of protein, the penetration development is facilitated by fungal proteases like; trypsins, chymotrypsins, metalloproteases and subtilisins, these toxic metabolites with insecticidal effect include substances released by different entomopathogenic fungi such as; detrains produced by Metarhizium species, beauvericins, isarolides, bassianolides, beauverolides oosporein produced by Beauveria species or Isaria fumosorosea, efrapeptins produced by Tolypocladium species and hirsutellin released from Hirsutella thompsonii4-6. The existence of entomopathogenic fungi is quite wide and can be found almost all over the world (e.g., tropical rainforest, Antarctica or Arctic), so they are a worldwide spread group of fungi7,8.
In temperate regions of the world, the order hypocreales and entomophthorales are generally known infectious to many insect species. The hypocrealean species B. bassiana and M. anisopliae are known to infect many insect species related to agro-ecosystems9,10. Recent advance studies on entomopathogenic fungi clarified the importance of fungi as a conservation microbial control agent against many insect pests of field crops and forest trees. It is reported that most of B. bassiana species associated with insect pests above ground level, while M. anisopliaeis associated either on or underground11-13.
Entomopathogenic fungi occur in nature and are capable to cause epizootics and decrease the insect population and represent a larger portion of present bio-pesticide in the market worldwide14-17. Viewing importance of entomopathogenic fungi in insect control the current review article is compiled to find out the hurdles/factors which effect on the virulence and pathogenicity of entomopathogenic fungi. This review will provide useful information regarding entomopathogenic fungi and their success in various environmental conditions.
CHARACTERISTICS AFFECTING VIRULENCE OF FUNGI
Fungal taxonomy: Literature indicated that insect-pathogenic fungi are true fungi and belongs to different phylum i.e., Ascomycota, Zygomycota, Chytridiomycota and Basidiomycota Coelomomyces, among these the genus Chytridiomycota known as a good aquatic entomopathogenic species which produce zoospores asexually and significantly affect the many Dipterian insects including mosquitoes. The Zygomycota phylum is further divided into sub-divisions Zygomycotina, class Zygomycetes and order Entomophthorales18,1. These fungi are capable to infect various insect species. In case of unavailability of suitable host, most of fungi have ability to produce motionless spores, which also can live under unfavourable conditions. For example, during winter season these spores either produced sexually or asexually. The well-known common genera are Neozygites, Erynia, Zoophthora, Conidiobolus and Entomophaga19,20. While a larger portion of Entomophthorales are host-specific, e.g., Entomophthorales Neoaphidis species only reported from aphids. Whereas, Zoophthora radicans (Brefeld) Batko was documented from different insect species, however, most of the individual isolate infect particular insect species21. The Deuteromycotina is the main group of entomopathogen and known as “Imperfecti”. The reproduction in class Hyphomycetes took place asexually, however, sexual production is occurred rarely. The sexual behaviour occurs in some species of Cordyceps, but some researchers suggested that these species should be placed in the group of Ascomycota22. The fungus Deuteromycotina does not produce inactive spores and has the ability to survive in the soil23.
The well-known Entomopathogeneous fungi which belong to class Hyphomycetes are Verticillium, Metarhizium, Aschersonia, Beauveria, Paecilomyces, Nomuraea and Hirsutella. Among these the most popular species is Beauveria that cause “white muscardine” disease9,24,25. As being a good pathogenic fungus the many studies were conducted in this genus that is V. lecanii26, whereas, the genus Metarhizium known as “green muscardine” fungus, reviewed and documented by Kryukov et al.27. Entomopathogenic fungi are fungi having denticulate (tooth-like or bristle-like structure) apical extensions (rachis) containing single conidium per denticle and conidia are a-septate. Conidia of B. bassiana are approximately round ball-shaped about <3.5 pm diameter, whereas, Isaria fumosorosea conidia are grown individually or in clusters form on ringlets on branches of conidiophores or on short side branches or in apical whorls conidia are a-septate and range from colorless pigment. Conidia of Isaria fumosorosea are long, ovoid and <4 pm long28.
For example, Isaria farinos a need carbon and nitrogen for stimulating growth and developmental stage of germ tube29. The germ tube penetrates into insect body through hyphae30. The developmental stage of an “appressorium” thickened flattened most upper tip of a hyphal out-growth, which assist the fungi to bind and penetrate into host cuticle31.
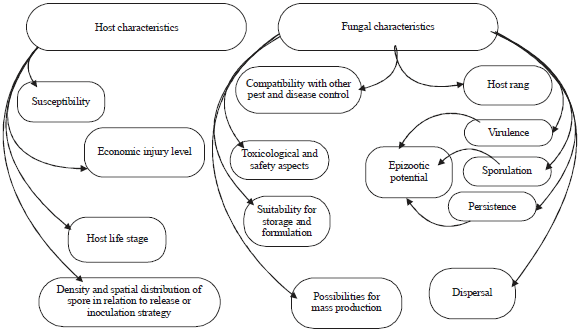 | |
| Fig. 1: | Host and fungal characteristic effect on pathogenicity and infection process of fungi |
Some of hyphomycetes fungi can cause death before the extensive intrusion of insect tissues. These types of entomopathogenic fungi cause mortality by releasing toxic substances12,32. After infection, the fungal spore emerges from insect cuticle and starts sporulation on the insect dead body. Many fungal hyphae primarily grow inside the inter-segmental regions of the host. This provides a basic pathway of inoculation for the infection in insect cuticle. Sometimes due to unfavourable conditions, the spore remain alive as mummified internally inside the host cadaver for a long time and germinate as conditions become favourable31. It releases in the environment or in the field when insect cadavers break down. This behaviour makes them more suitable for large scale insect control33.
Most of the Entomopathogenic fungi are able to cause epizootic disease in the insect population. Epizootic mean as “unusually large number of cases of disease" in a host population34 the prevalence of fungal infection in insect host is similar to that of other organisms. However epizootic disease development depends on host susceptibility, population and spreads efficacy through the contact between infected hosts to a healthy one14,35. The horizontal transmission maybe accorded form one host to another36,37 the behaviour of horizontal transmission is mostly found in hyphomycete fungi, the horizontal transmission was reported in many insects such as; aphid, grasshoppers, locusts and Diaphorina citri. A high rate of host population increases the chances of contact among individuals and cause pathogen transmission, because overpopulation increases the stress and fungal infection38,39 (Fig. 1).
CHARACTERISTICS INVOLVING IN INFECTION PROCESSES
Unlike other pathogens that required to be ingested, insect pathogenic fungi directly penetrate in the host cuticle40. The complex infectious process involved mechanical incursion, while exhibiting different extracellular enzymes, which are responsible post-infection41-43. As the insect cuticle contains up to 70% of protein, the penetration process is facilitated by extracellular fungal proteases like subtilisins, chymotrypsins, trypsins and metalloproteases, usually with multiple isoforms of each. Toxic metabolites with insecticidal effect include substances as; beauvericins, isarolides, bassianolides, beauverolides, oosporein produced by Beauveria species or Isaria fumosorosea4,5. These substances facilitate to kill many different species of insect pests and arthropods, their capacity to facilitate penetrate into insect cuticle due this they become able to breakdown immune system of insects44. The insects killed infection coming into contact spray droplets on its body, through walking on treated area or by feeding plants treated with the mycoinsecticides once the pathogenic conidia attached to the target host skin, they develop hyphae which penetrate into the insect’s cuticle and start multiplication45. Susceptible host cuticle components play a dietetic role for the development of the conidial germ tube depends on fungal species. After penetration, the fungus germinates and multiplies inside the host insect haemocoel. The infected insect host usually killed 3-14 days after penetration of germ tube.
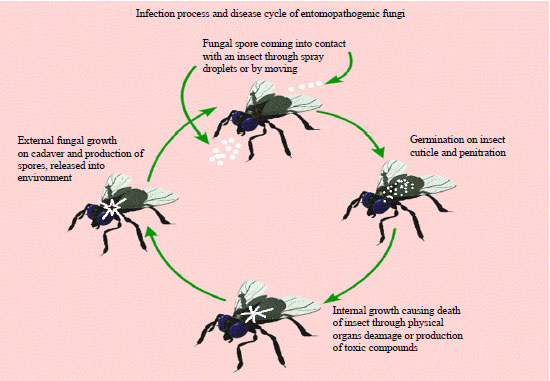 | |
| Fig. 2: | Infection and disease development cycle |
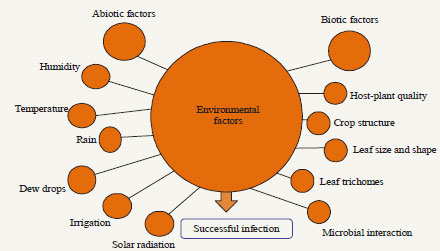 | |
| Fig. 3: | Biotic and abiotic factors involves in infection and in disease development processes |
However, sometimes it takes longer time because of fungal intrinsic potential, the number of conidia applied on targeted host species and on environmental condition as well15,46,47 (Fig. 2).
FACTORS INVOLVES IN INFECTION PROCESSES
Biotic factors: The host-plant chemistry plays an important factor which directly affects fungal infection, by suppression or enhancing the conidial germination or via the insect, influencing its susceptibility or its development rate48,49.
Plant quality, leaf structure, shape and leaf trichomes and microbial interactions are also important factors22,50. Host-specific is common in almost all entomopathogenic-fungi (Table 1) due to specific host characteristics these fungi only effects on target insect pest and no direct effect on other beneficial insects i.e., parasites and predators. Though, no significant relationship was founded in the virulence of fungi other than its original host insect51,52, however, in some cases studies showed that the repeated mass production of the same species on artificial medium loss the virulence due to changes in physiological and developmental process of fungi53. The pose influence on the pathogenicity of fungi directly either enhancing or by inhibiting conidial germination on insect cuticle48,49 (Fig. 3).
| Table 1: | Pathogenicity level of different entomopathogenic fungi against various insect pests at different environmental conditions |
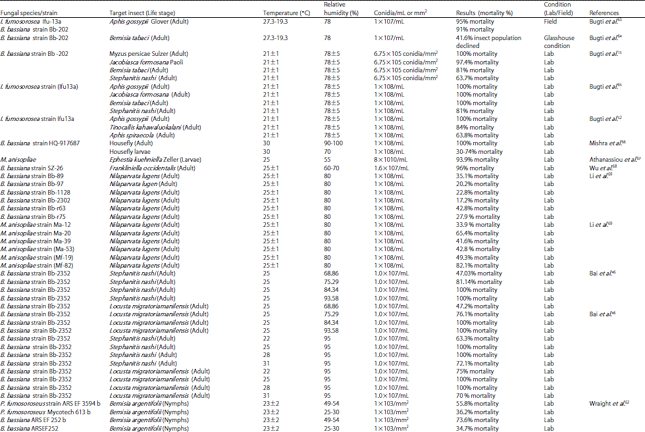 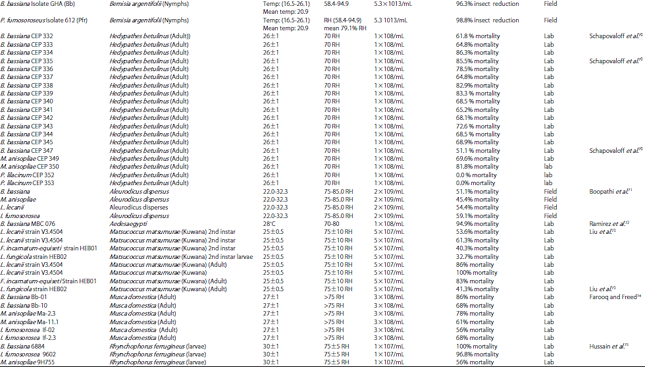 | |
Abiotic factors: Suitable environmental factors i.e., temperature and relative humidity play an important role in disease development and infection process of fungi54,55. Humidity is more important than temperature, which poses great influence on the inducing fungal disease in the host53,56. However, literature showed that 50% relative humidity also helpful infection process in some cases46,57. High humidity helps conidia to penetrate in the target host (Table 1).
It is reported58 that the relative humidity provides better results as compare than temperature in the glasshouses most of insect-pathogenic fungi grow and sporulate at optimum temperature ranged from 15-30°C46,59. The ideal range of temperature for greatly enhances the growth and rapid infection, though germination of conidia retarded the developmental process of host insect. However, many insects escape from fungal infection by molting60,61. That’s why most of the entomopathogenic fungi fail to induce fungal disease in earlier larval stage62 (Table 1).
CONCLUSION
AND RECOMMENDATIONS
It was concluded from previous literature that higher fungal conidial density provides rapid and better control. The study also showed that most of the fungi are host-specific. Moreover, host-specificity is not related to phylogenetic relationship. This was observed that initial infection is the most critical stage of causing an epizootic disease, once fungi built a successful infection in targeted insect population, it sharply spread and kills their hosts. Study further revealed that the virulence of entomopathogenic fungi significantly affected by biotic and abiotic factors such as; fungal species, host plant chemistry, insect host and its life stages and temperature, sunlight and relative humidity, respectively. Among these factors, the relative humidity seems to be more important than other factors to ensure infection and successful disease development in targeted insect population.
The farmers/horticulturists from the coastal areas with high relative humidity over than 60% with moderate temperature are strongly advised to use conidia base myco-insecticides for insect control to limit excessive application of chemical pesticides and make sure a healthy environment.
SIGNIFICANCE STATEMENT
This review article will help the researcher to understand various factors, which are involved in causing infection and successful disease development in many insect pests. This review also provides information about effective range of temperature and humidity of different Entomopathogenic fungal species to manage the insect pest infestation.
REFERENCES
- Shang, Y., P. Feng and C. Wang, 2015. Fungi that infect insects: Altering host behavior and beyond. PLoS Pathog., Vol. 11.
CrossRefDirect Link - Jaronski, S.T., 2010. Ecological factors in the inundative use of fungal entomopathogens. BioControl, 55: 159-185.
CrossRefDirect Link - Du, F.Y., X.M. Li, P. Zhang, C.S. Li and B.G. Wang, 2014. Cyclodepsipeptides and other O-containing heterocyclic metabolites from Beauveria felina EN-135, a marine-derived entomopathogenic fungus. Mar. Drugs, 12: 2816-2826.
CrossRefDirect Link - Charnley, A.K., 2003. Fungal pathogens of insects: cuticle degrading enzymes and toxins. Adv. Bot. Res., 40: 241-321.
CrossRefDirect Link - Ramzi, S. and A. Zibaee, 2014. Biochemical properties of different entomopathogenic fungi and their virulence against Chilo suppressalis (Lepidoptera: Crambidae) larvae. Biocontrol Sci. Technol., 24: 597-610.
CrossRefDirect Link - Aung, O.M., K. Soytong and K.D. Hyde, 2008. Diversity of entomopathogenic fungi in rainforests of Chiang Mai province, Thailand. Fungal Divers., 30: 15-22.
Direct Link - Goffre, D. and P.J. Folgarait, 2018. Insights into the biodiversity and causes of distribution of potential entomopathogens associated with leaf-cutting ants. Insectes Sociaux, 65: 103-115.
CrossRefDirect Link - Aak, A., M. Hage and B.A. Rukke, 2018. Insect pathogenic fungi and bed bugs: behaviour, horizontal transfer and the potential contribution to IPM solutions. J. Pest Sci., 91: 823-835.
CrossRefDirect Link - Ibarra-Cortes, K.H., A.W. Guzman-Franco, H. Gonzalez-Hernandez, L.D. Ortega-Arenas, J.A. Villanueva-Jimenez and A. Robles-Bermudez, 2018. Susceptibility of Diaphorina citri (Hemiptera: Liviidae) and its parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) to entomopathogenic fungi under laboratory conditions. Neotrop. Entomol., 47: 131-138.
CrossRefDirect Link - Dong, T.Y., B.W. Zhang, Q.F. Weng and Q.B. Hu, 2016. The production relationship of destruxins and blastospores of Metarhizium anisopliae with virulence against Plutella xylostella. J. Integr. Agric., 15: 1313-1320.
CrossRefDirect Link - Raya-Diaz, S., A.R. Sanchez-Rodriguez, J.M. Segura-Fernandez, M.D.C. del Campillo and E. Quesada-Moraga, 2017. Entomopathogenic fungi-based mechanisms for improved Fe nutrition in sorghum plants grown on calcareous substrates. Plos One, Vol. 12, No. 10.
CrossRefDirect Link - Cabanillas, H.E. and W.A. Jones, 2013. Pathogenicity of Isaria poprawskii (Ascomycota: Hypocreales: Cordycipitaceae) against the glassy-winged sharpshooter, Homalodisca vitripennis (Hemiptera: Cicadellidae), under laboratory conditions. Crop Prot., 50: 46-52.
CrossRefDirect Link - Bugti, G.A., W. Bin, H.F. Lin, C. Na and L.H. Feng, 2018. Pathogenicity of Beauveria bassiana strain 202 against sap-sucking insect pests. Plant Protect. Sci., 54: 111-117.
CrossRefDirect Link - Silva, J.D.N., G.M. Mascarin, I.C.D.S. Gomes, R.S. Tinoco, E.D. Quintela, L.D.R. Castilho and D.M.G. Freire, 2018. New cost-effective bioconversion process of palm kernel cake into bioinsecticides based on Beauveria bassiana and Isaria javanica. Applied Microbiol. Biotechnol., 102: 2595-2606.
CrossRefDirect Link - Muniz-Paredes, F., F. Miranda-Hernandez and O. Loera, 2017. Production of conidia by entomopathogenic fungi: From inoculants to final quality tests. World J. Microbiol. Biotechnol., Vol. 33, No. 3.
CrossRefDirect Link - Chen, B., Z.Y. Li and M.G. Feng, 2008. Occurrence of entomopathogenic fungi in migratory alate aphids in Yunnan Province of China. Biocontrol, 53: 317-326.
CrossRefDirect Link - Roy, H.E., D.C. Steinkraus, J. Eilenberg, A.E. Hajek and J.K. Pell, 2006. Bizarre interactions and endgames: Entomopathogenic fungi and their arthropod hosts. Annu. Rev. Entomol., 51: 331-357.
CrossRefPubMedDirect Link - Hall, D.G., M.G. Hentz, J.M. Meyer, A.B. Kriss, T.R. Gottwald and D.G. Boucias, 2012. Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorina citri (Hemiptera: Psyllidae) in a managed citrus grove. Biocontrol, 57: 663-675.
CrossRefDirect Link - Fronza, E., A. Specht, H. Heinzen and N.M. de Barros, 2017. Metarhizium (Nomuraea) rileyi as biological control agent. Biocontrol Sci. Technol., 27: 1243-1264.
CrossRefDirect Link - Zimmermann, G., 2008. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol., 18: 865-901.
CrossRefDirect Link - Kryukov, V., O. Yaroslavtseva, M. Tyurin, Y. Akhanaev and E. Elisaphenko et al., 2017. Ecological preferences of Metarhizium spp. from Russia and neighboring territories and their activity against Colorado potato beetle larvae. J. Invertebr. Pathol., 149: 1-7.
CrossRefDirect Link - Liu, F., X.L. Wu, Z.Y. He, M.C. Xiang and Y.C. He et al., 2016. Nutritional effects on the mycelial growth and enzymatic activity of Isaria farinosa, and Hepialus larvae growth. J. Applied Microbiol., 120: 1605-1615.
CrossRefDirect Link - Lu, L.M., B.P. Cheng, D.C. Du, X.R. Hu and A.T. Peng
., 2015. Morphological, molecular and virulence characterization of three Lencanicillium species infecting Asian citrus psyllids in Huangyan citrus groves. J. Invertebr. Pathol., 125: 45-55.
CrossRefDirect Link - Hatzipapas, P., K. Kalosaka, A. Dara and C. Christias, 2002. Spore germination and appressorium formation in the entomopathogenic Alternaria alternata. Mycol. Res., 106: 1349-1359.
CrossRefDirect Link - Wang, C.Z., T.N. Gao, Y. Huang and Z. Huang, 2017. Effect of Ifchit1 gene of Isaria fumosorosea on mortality, oviposition and oxidase activities of Bemisia tabaci. Biocontrol Sci. Technol., 27: 485-495.
CrossRefDirect Link - Cabanillas, H.E. and W.A. Jones, 2009. Pathogenicity of Isaria sp. (Hypocreales: Clavicipitaceae) against the sweet potato whitefly B biotype, Bemisia tabaci (Hemiptera: Aleyrodidae). Crop Prot., 28: 333-337.
CrossRefDirect Link - Tian, J., H.L. Diao, L. Liang, S. Arthurs, C. Hao, G.M. Mascarin and R.Y. Ma, 2016. Host plants influence susceptibility of whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) to the entomopathogenic fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae). Biocontrol Sci. Technol., 26: 528-538.
CrossRefDirect Link - Draganoval, S.A., D.D. Doychev, D.K. Pilarska and D.I. Takov, 2017. Bioassays of entomopathogenic fungi against Xylophagous insects in Bulgaria: Laboratory and field experiments. Acta Zool. Bulg., 69: 411-419.
Direct Link - Arthurs, S.P., M.B. Thomas and J.L. Lawton, 2001. Seasonal patterns of persistence and infectivity of Metarhizium anisopliae var. acridum in grasshopper cadavers in the Sahel. Entomol. Exp. Applic., 100: 69-76.
CrossRefDirect Link - Shapiro-Ilan, D.I., T.E. Cottrell, M.A. Jackson and B.W. Wood, 2008. Virulence of Hypocreales fungi to pecan aphids (Hemiptera: Aphididae) in the laboratory. J. Invertebr. Pathol., 99: 312-317.
CrossRefDirect Link - Conceschi, M.R., C.P. D’Alessandro, R. de Andrade Moral, C.G.B. Demetrio and I. Delalibera, Jr., 2016. Transmission potential of the entomopathogenic fungi Isaria fumosorosea and Beauveria bassiana from sporulated cadavers of Diaphorina citri and Toxoptera citricida to uninfected D. citri adults. BioControl, 61: 567-577.
CrossRefDirect Link - Hajek, A.E. and R.J. St. Leger, 1994. Interactions between fungal pathogens and insect hosts. Annu. Rev. Entomol., 39: 293-322.
CrossRefDirect Link - St Leger, R.J., L. Joshi and D.W. Roberts, 1997. Adaptation of proteases and carbohydrases of saprophytic, phytopathogenic and entomopathogenic fungi to the requirements of their ecological niches. Microbiology, 143: 1983-1992.
CrossRefDirect Link - Ortiz-Urquiza, A. and N.O. Keyhani, 2013. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects, 4: 357-374.
CrossRefDirect Link - Gökçe, A. and M.K. Er, 2014. Pathogenicity of Paecilomyces spp. to the glasshouse whitefly, Trialeurodes vaporariorum, with some observations on the fungal infection process. Turk. J. Agric. For., 29: 331-340.
Direct Link - Gabarty, A., H.M. Salem, M.A. Fouda, A.A. Abas and A.A. Ibrahim, 2014. Pathogencity induced by the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in Agrotis ipsilon (Hufn.). J. Radiat. Res. Applied Sci., 7: 95-100.
CrossRefDirect Link - Long, D.W., F.A. Drummond, E. Groden and D.W. Donahue, 2000. Modelling Beauveria bassiana horizontal transmission. Agric. For. Entomol., 2: 19-32.
CrossRefDirect Link - Bai, Y., Y. Cui, N. Cao, Y. Liu, G.A. Bugti and B. Wang, 2016. Effects of humidity and temperature on the pathogenecity of Beauveria bassiana against Stephanitis nashi and Locusta migratoria manilensis. Chin. J. Biol. Control, 32: 735-742.
CrossRefDirect Link - Wilson, W.M., J.E. Ibarra, A. Oropeza, M.A. Hernandez, R.A. Toledo-Hernandez and J. Toledo, 2017. Infection of Anastrepha ludens (Diptera: Tephritidae) adults during emergence from soil treated with Beauveria bassiana under various texture, humidity and temperature conditions. Florida Entomol., 100: 503-508.
CrossRefDirect Link - Blakeman, J.P., 1973. The chemical environment of leaf surfaces with special reference to spore germination of pathogenic fungi. Pestic. Sci., 4: 575-588.
CrossRefDirect Link - Poprawski, T.J., S.M. Greenberg and M.A. Ciomperlik, 2000. Effect of host plant on Beauveria bassiana and Paecilomyces fumosoroseus induced mortality of Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Environ. Entomol., 29: 1048-1053.
CrossRefDirect Link - Gan, H., A.C.L. Churchill and K. Wickings, 2017. Invisible but consequential: Root endophytic fungi have variable effects on belowground plant-insect interactions. Ecosphere, Vol. 8, No. 3.
CrossRefDirect Link - Mor, H., G. Gindin, I.S. Ben-Ze’ev, B. Raccah, N.U. Geschtovt, N. Ajtkhozhina and I. Barash, 1996. Diversity among isolates of Verticillium lecanii as expressed by DNA polymorphism and virulence towards Bemisia tabaci. Phytoparasitica, 24: 111-118.
CrossRefDirect Link - Bugti, G.A., C. Na, W. Bin and L.H. Feng, 2017. Pathogenicity of Isaria fumosorosea strain Ifu 13a against different aphid species. J. Kansas Entomol. Soc., 90: 365-372.
CrossRefDirect Link - Santos, A.H., M.H.H. Tai, L.F.N. Rocha, H.H.G. Silva and C. Luz, 2009. Dependence of Metarhizium anisopliae on high humidity for ovicidal activity on Aedes aegypti. Biol. Control, 50: 37-42.
CrossRefDirect Link - Ment, D., G. Gindin, I. Glazer, S. Perl, D. Elad and M. Samish, 2010. The effect of temperature and relative humidity on the formation of Metarhizium anisopliae chlamydospores in tick eggs. Fungal Biol., 114: 49-56.
CrossRefDirect Link - Cui, J.H., Z.J. Tan and H.T. Chen, 2012. Pathogenicity of Beauveria bassiana to Spodoptera exigua larvae at different temperature and humidity. Acta Agric. Jiangxi, 24: 41-43.
Direct Link - Lin, H., M. Fan, Z. Li and C. Hu, 1998. Pathogenic effect of Beauveria bassiana infected on Dendrolimus punctatus under different temperature and humidity. Chin. J. Applied Ecol., 9: 195-200.
Direct Link - Bateman, R.P., M. Carey, D. Moore and C. Prior, 1993. The enhanced infectivity of Metarhizium flavoviride in oil formulations to desert locusts at low humidities. Ann. Applied Biol., 122: 145-152.
CrossRefDirect Link - Lord, J.C., 2005. Low humidity, moderate temperature and desiccant dust favor efficacy of Beauveria bassiana (Hyphomycetes: Moniliales) for the lesser grain borer, Rhyzopertha dominica (Coleoptera: Bruchidae). Biol. Control, 34: 180-186.
CrossRefDirect Link - Tang, L.C. and R.F. Hou, 2001. Effects of environmental factors on virulence of the entomopathogenic fungus, Nomuraea rileyi, against the corn earworm, Helicoverpa armigera (Lep., Noctuidae). J. Applied Entomol., 125: 243-248.
CrossRef - Luz, C., J. Fargues and C. Romana, 2003. Influence of starvation and blood meal-induced moult on the susceptibility of nymphs of Rhodnius prolixus Stål (Hem., Triatominae) to Beauveria bassiana (Bals.) Vuill. infection. J. Applied Entomol., 127: 153-156.
CrossRefDirect Link - Wraight, S.P., R.I. Carruthers, S.T. Jaronski, C.A. Bradely, J.C. Garza and S. Galaini-Whraight, 2000. Evaluation of the entomopathogenic fungi Beauveria bassiana and Paecilomyces fumosoroseus for microbial control of the silverleaf whitefly, Bemisia argentifolii. Biol. Control, 17: 203-217.
CrossRefDirect Link - Bugti, G.A., N. Cao, L.H. Feng and W. Bin, 2020. Field bioassay assessment of entomopathogenic fungi on Aphis gossypii (Glover). J. Entomol., 17: 20-26.
CrossRefDirect Link - Bugti, G.A., C. Na, L.H. Feng and W. Bin, 2019. Control efficiency of entomopathogenic fungal species against Bemisia tabaci (Gennadius) in laboratory and glasshouse condition. Proceedings of the 3rd Iranian International Congress of Entomology, (IICE'19), The University of Tabriz, Iran, pp: 273-289.
- Bugti, G.A., C. Na, W. Bin and L.H. Feng, 2018. Control of plant sap-sucking insects using entomopathogenic fungi Isaria fumosorosea strain (Ifu13a). Plant Prot. Sci., 54: 258-264.
CrossRefDirect Link - Mishra, S., P. Kumar and A. Malik, 2015. Effect of temperature and humidity on pathogenicity of native Beauveria bassiana isolate against Musca domestica L. J. Parasitic Dis., 39: 697-704.
CrossRefDirect Link - Athanassiou, C.G., N.G. Kavallieratos, C.I. Rumbos and D.C. Kontodimas, 2017. Influence of temperature and relative humidity on the insecticidal efficacy of Metarhizium anisopliae against larvae of Ephestia kuehniella (Lepidoptera: Pyralidae) on wheat. J. Insect Sci., Vol. 17, No. 1.
CrossRefDirect Link - Wu, S., Y. Gao, Y. Zhang, E. Wang, X. Xu and Z. Lei, 2014. An entomopathogenic strain of Beauveria bassiana against Frankliniella occidentalis with no detrimental effect on the predatory mite Neoseiulus barkeri: Evidence from laboratory bioassay and scanning electron microscopic observation. PLoS ONE, Vol. 9.
CrossRefDirect Link - Li, M., H. Lin, S. Li, P. Chen, L. Jin and J. Yang, 2012. Virulence of entomopathogenic fungi to adults and eggs of Nilaparvata lugens Stal (Homopera: Delphacidae). Afr. J. Agric. Res., 7: 2183-2190.
Direct Link - Schapovaloff, M.E., L.F.A. Alves, A.L. Fanti, R.A. Alzogaray and C.C. López Lastra, 2014. Susceptibility of adults of the cerambycid beetle Hedypathes betulinus to the entomopathogenic fungi Beauveria bassiana, Metarhizium anisopliae and Purpureocillium lilacinum. J. Insect Sci., Vol. 14, No. 1.
CrossRefDirect Link - Boopathi, T., P. Karuppuchamy, S.B. Singh, M. Kalyanasundaram, S. Mohankumar and M. Ravi, 2015. Microbial control of the invasive spiraling whitefly on cassava with entomopathogenic fungi. Braz. J. Microbiol., 46: 1077-1085.
CrossRefDirect Link - Ramirez, J.L., C.A. Dunlap, E.J. Muturi, A.B.F. Barletta and A.P. Rooney, 2018. Entomopathogenic fungal infection leads to temporospatial modulation of the mosquito immune system. PLOS Negl. Trop. Dis., Vol. 12, No. 4.
CrossRefDirect Link - Liu, W., Y. Xie, J. Dong, J. Xue, Y. Zhang, Y. Lu and J. Wu, 2014. Pathogenicity of three entomopathogenic fungi to Matsucoccus matsumurae. PloS One, Vol. 9, No. 7.
CrossRefDirect Link - Farooq, M. and S. Freed, 2016. Infectivity of housefly, Musca domestica (Diptera: Muscidae) to different entomopathogenic fungi. Braz. J. Microbiol., 47: 807-816.
CrossRefDirect Link - Hussain, A., M. Rizwan-ul-Haq, H. Al-Ayedh and A.M. AlJabr, 2016. Susceptibility and immune defence mechanisms of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) against entomopathogenic fungal infections. Int. J. Mol. Sci., Vol. 17.
CrossRefDirect Link