
Hallouti Ayoub
Laboratory of Microbial Biotechnology and Plant Protection, Faculty of Sciences, Ibn Zohr University, B.P 8106, Agadir, Morocco
Zahidi Abdelaziz
Faculty of Applied Science, Ait Melloul Campus, Ibn Zohr University, Agadir, Morocco
LiveDNA: 212.31502
Lamharchi El Hassane
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P 8106, Agadir, Morocco
Boubaker Hassan
Laboratory of Microbial Biotechnology and Plant Protection, Faculty of Sciences, Ibn Zohr University, B.P 8106, Agadir, Morocco
LiveDNA: 212.8065
El Mousadik Abdelhamid
Laboratory of Microbial Biotechnology and Plant Protection, Faculty of Sciences, Ibn Zohr University, B.P 8106, Agadir, Morocco
LiveDNA: 212.23672
Ait Benaoumar Abdellah
Laboratory of Biotechnology and Valorization of Natural Resources, Faculty of Sciences, Ibn Zohr University, B.P 8106, Agadir, Morocco
Journal of Entomology
Year: 2020 | Volume: 17 | Issue: 2 | Page No.: 58-67







ABSTRACT
Background and Objective: Ceratitis capitata is considered as one of the most economically destructive pest. Several alternatives have been used but efficacy remains limited. The objective consists in the search for other natural enemies such as parasitoids and evaluate parasitism rate of larval stages from infested fruits by medfly collected in argane forest. Materials and Methods: About 400 fly larvae per sample and per locality were isolated and placed in breeding boxes at optimal temperature until the emergence of adults. Number of Ceratitis adults and parasitism rate have been observed. Morphological and molecular identification using COI-F/COI-R primers of collected parasitoids were also occurred. Results: Morphological and molecular analyses confirmed the occurrence of a new haplotype of the larval parasitoid belonging to Psyttalia genus in argane forest. In addition, phylogenetic analyses revealed that these specimens were 92% (E-value = 2e-85) identical to Psyttalia lounsburyi. The presence of this parasitoid was noted in all studied localities with a variable parasitism rate which reached a value of 21.5% in the localities where agricultural activities are present. Conclusion: It can be concluded that this natural parasitoid could be native to argane forest and will be used as biological control agent against Ceratitis capitata and Bactrocera oleae in Morocco. Also in Mediterranean region which constitute a handicap to the extension of biological agriculture of fruit trees.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hallouti Ayoub, Zahidi Abdelaziz, Lamharchi El Hassane, Boubaker Hassan, El Mousadik Abdelhamid and Ait Benaoumar Abdellah, 2020. First Report of Psyttalia lounsburyi, a New Larval Parasitoid of the Mediterranean Fruit Fly Ceratitis capitata in Moroccan Argane Forest. Journal of Entomology, 17: 58-67.
DOI: 10.3923/je.2020.58.67
URL: https://scialert.net/abstract/?doi=je.2020.58.67
DOI: 10.3923/je.2020.58.67
URL: https://scialert.net/abstract/?doi=je.2020.58.67
INTRODUCTION
The Mediterranean fruit fly (Ceratitis capitata Wiedemann) also called the medfly is a fruit fly of the Tephritidae family. Despite of being native to tropical Africa and especially West Africa, this pest is continuously expand its geographic distribution globally and now occurs on five continents1. Attacking over 300 host plant species, this fruit fly is considered one of the most destructive pests of fruits production in the world especially in the Mediterranean region and Morocco2-4. The damages of the medfly are caused by the laying of the female in the fruit and the larval development inside fruits. Because larvae of this pest live inside the fruit, control is exceedingly difficult and expensive. Consequently, the medfly is responsible of extensive economic and crop losses5,6. Besides causing large damages to production, producer countries lose their international markets due to quarantine regulation against this pest in several importing countries. Accordingly, the success of fruit industries in the Mediterranean region and in Morocco depends heavily on the medfly management5,7.
Moroccan moderate climate coupled to intensive monoculture of fruits and wide Argania spinosa forests (wild host plant of medfly) form a favourable environment with uninterrupted supply of host fruits for the medfly to multiply and breed3,8. Under such circumstances, the development of extensive control programmes becomes a priority. In the current time the control of the medfly mostly based on using cover sprays of insecticides with the carbamate, organophosphate and synthetic pyrethroid when fruits becomes susceptible to oviposition in order to prevent direct attacks of fruits5. In addition, spot sprays of protein baits coupled to orchard sanitation and fruit wrapping aimed at reducing the medfly population. This chemical compounds when misused can induce ecological and toxicological problems5,9,10. In order to reduce the negative effects of chemical control, it is necessary to develop other alternative strategies for managing populations of this pest.
The use of biological control agents such as parasites, parasitoids and predators could be a promising and sustainable alternative to chemical control by pesticides. In addition to being safe, economical and permanent solution, natural enemies are proving effective in reducing medfly populations in many regions to economic tolerance thresholds11. Furthermore, a major biological control effort against the medfly was carried out using egg-larval parasitoids belonging to Psyttalia and Fopius genera such as Fopius arisanus, Fopius concolor and Fopius ceratitovorus and Psyttalia fletcheri (Braconidae)11-16. In this context, one of the most successful examples of using natural parasitoids to control fruit flies including C. capitata was in Hawaii by introducing over 20 parasitoids including one egg-larval parasitoid, Fopius arisanus (Sonan) and seven larval parasitoids: Psyttalia fletcheri, F. Vandenbosch, Diachasmimorpha tryoni, Diachasmimorpha longicaudata, Procecidochares incise, Opius Artun and Tetrastichus givardianus (Silvestri)12,14,17-20.
Psyttalia lounsburyi (Silvestri) is a larval parasitoid of Braconidae family used against Bactrocera oleae. It can also parasitize Ceratitis capitata larvae as reported by several studies11,21,22. This parasitoid discovered for the first time in East Africa (Kenya), is known by its endoparasitism of B. oleae23. The female of P. lounsburyi attacks the first larval stages of its host24,25. In addition, larvae located near to the surface of infested fruits are more susceptible to this insect’s attacks and therefore these larvae are easily infected than those protected by fruits tissues13,26,27. Currently, several researches are interested in this type of parasitism and its application in biological control especially in USA and France against B. oleae24,25,28. Host suitability and host range studies are demonstrated that B. oleae is the specific host of P. lounsburyi24,28, however, other studies showed the possibility of attacking other pest such as C. capitata26. In fact, this parasitoid attacks successfully larvae of C. capitata under laboratory conditions and could be easily multiplied on this fly26. In this context and with the aim of widening the range of natural enemies against ceratitis, this work has been undertaken. The objective consists in the search for other natural enemies such as parasitoids and evaluate parasitism rate of larval stages from infested fruits by medfly collected in argane forest.
MATERIALS AND METHODS
The study was carried out at the Faculty of Sciences Agadir, from March, 2018-February, 2019 in Biotechnologies and Valorization of Natural Resources and Microbial Biotechnologies and Plant Protection. Molecular analyzes (DNA extraction and sequencing) were carried out at the Technical Support Units for Scientific Research at the National Center for Scientific and Technical Research (CNRST) Rabat Morocco. This study is in continuity until today.
Insect collection and identification: During research work on entomopathogenic fungi against C. capitata isolated from argane (wild host plant of medfly) soil (thesis data), infested fruits with this fly were also collected in Souss region. Following this sampling, which occurred in May 2018, it was noted the presence in addition to fruit fly in stars several specimens (adults) of Hymenoptera.
| Table 1: | Collect localities geographical references |
 | |
In addition, several infested larvae and pupae of C. capitata were also observed. In order to study this new parasitoid, several collects of infested fruit by the host insect (C. capitata) were done between May-June, 2018 in different localities in Argania spinosa Forest (Table 1). Infested fruits thus collected were placed at room temperature in laboratory to recover pest larvae. About 400 larvae were isolated from each sample per locality and placed in boxes until the emergence of adults. During this study, the parasitism percentage and the number of the insect’s adults were calculated for each sampling site. Some parasitoid were conserved in 90% alcohol and stored at -4°C. Collected specimens were identified according to Wharton description keys11,21,22,29,30-32. Other specimens were sent to Technical Support Unit for scientific research at the National Center for Scientific and Technical Research (CNRST) in Rabat for molecular analysis.
Molecular analysis: In order to confirm morphological identification, total genomic DNA was extracted from insect’s adults according to kit protocol (BIOLINE) following standard protocol for animal tissues. Concentration and purity of DNA of sample are measured by a 8000-Nanodrop-photometer. Primers used for PCR amplification in this study were COI-F (5'GTCACTTGCCGGCATTTGGATTATTT-3') as a forward and COI-R (5'TGATTTTTT GTCACCCA GAAGTGTTA-3') as a reverse.
The PCR amplification of the mitochondrial Cytochrome Oxidase I (COI) gene was performed in a total of 25 μL reaction volume with the following concentrations: PCR buffer (5x), MgCl2 (2 mM), dNTP (0.2 mM of each), primer (COI-F 10 μM and COI-R 10 μM (1 μL of each)), Taq polymerase (0.2 μL) (MyTaq DNA polymerase kit, Bioline) insect DNA (150 ng). The conditions for PCR were the following: Initial denaturation (95°C, 1 min) followed by 35 cycles of denaturation (95°C, 15 sec), primer annealing (60°C, 20 sec) and DNA extension (72°C, 15 sec). The reaction was held at 4°C.
The size of the amplified COI-region was determined using a 7 μL aliquot of the PCR product electrophoresed on 1.5% agarose gel. The ladder used was 100 bp. A volume of 5 μL of the amplified products were purified using purification Kit (ExoSAP-IT) to remove primers and nucleotides (dNTPS) not used during the PCR reaction. All sequencing reactions were prepared using a Big Dye Terminator v3.1 according to the manufacturer’s recommendations.
The obtained purified products were prepared for sequencing in the forward and reverse directions using 2 μL (3.5 pmol μL–1) of COI primers, BigDye Terminator v3.1 cycle sequencing kit’s (1 μL), Sequencing buffer (5x, 3 μL), DNA matrix (1.5 μL) and Milli-Q water (2.5 μL). The products were sequenced using an Applied Biosystem ABI 3130xl sequencer (Genetic Analyzer, 16 capillaries, Applied Biosystem).
The obtained sequencing results were submitted to a homology test using BLAST search function from the NCBI GenBank website. To confirm BLAST results, a Maximum-Likelihood phylogenetic analysis was applied to compare the specimen’s generated DNA sequences with other species form the Opiinae subfamily which are morphologically near to our insect using the Neighbor-Joining method. These species are: Diachasmimorpha longicaudata (GU097655.1), Phaenocarpa sp. (KJ412475.1), Pseudognaptodon sp. (KJ412473.1), Asobara japonica (JF430429.1), Cotesia autumnatae voucher (KC414765.1), Cotesia griffini (AF102704.1), Glyptapanteles indiensis (AF102713.1), Cotesia salebrosa voucher (KC414773.1), Habrobracon hebetor (MG744442.1), Fopius arisanus (KC581417.1), Psyttalia lounsburyi (DQ983810.1) and Psyttalia concolor TN0227 (EU761025.1). The DNA sequences of these species were collected from the NCBI Genbank website then compared to our P. lounsburyi (UIZ FSA-H1) COI gene sequence using MEGA6 software31.
Data analysis: Analysis of Variances (ANOVA) with one factor (sampling site factor) was adopted. The Least Significant Difference test (LSD α = 5%) of equality of means was used to compare differences between means for the three variables: Number of Psyttalia sp. adults, percentage of parasitism and percentage of natural mortality. All statistical analyzes were performed using the statistical version 6 software.
RESULTS
Morphological identification: Several morphological criteria characterize the parasitoid collected (Fig. 1). Size of individuals was between 2.3-3.4 mm (female adult) and 1.9-2.9 mm (male adult), with orange to brown color. The ovipositor is brown and 1.8±0.1 times longer than mesosoma. Wings are hyaline containing brown veins, with a large stigma about 2.9 times longer than wide, with rarising slightly in stigma midpoint; 2RS weakly sinuate; 3RSa 2.5 times longer than r; 3RSb ending slightly in wing tip (Fig. 2).
 | |
| Fig. 1(a-f): | Morphological characteristics of parasitoid, (a) Female lateral view, (b) Male lateral view, (c) Female up view (d) Female antennas (e) Male antennas and (f) Head |
Antennas with brown color are 3.0-3.1 times longer than the mesosoma and constituted by 31-37 segments in adult females and between 30-35 for adult males. The head of this insect is characterized by variable dimensions, it is 1.55-1.75 (average = 1.65±0.07) times wider than long. In addition, it was 1.25-1.35 times broader than mesoscutum, with face distinctly punctate throughout and containing an ocellar triangle with brown colour. Eyes are brown to black and they are very large and seemingly bare (no bristles), large, 1.5±0.3 times longer than wide, the temples are very receding in dorsal view. The maxillary palps are longer than the height of the head. Mesosoma 1.3±0.05 times longer than high and 1.65±0.05 times longer than broad. In the field, during the month of May, 2018 as well as in 2019, around 5 pm, a remarkable activity of the parasitoid was noted on some infested fruits by Ceratitis. This activity is observed at tree level and even on the fruit on the ground as illustrated in Fig. 3.
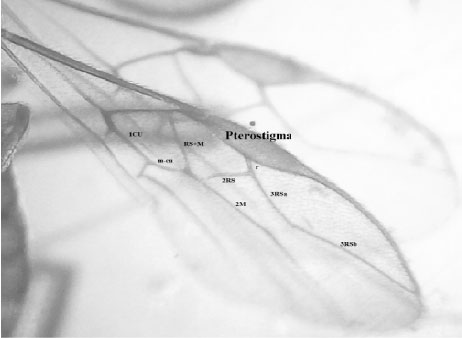 | |
| Fig. 2: | Observed Psyttalia sp. wings venation under binocular loupe (Gx40) |
 | |
| Fig. 3(a-f): | Activity of larvae parasitoid Psyttalia lounsburyi noted on some infested fruits by Ceratitis capitata in the field during 2018 -2019 |
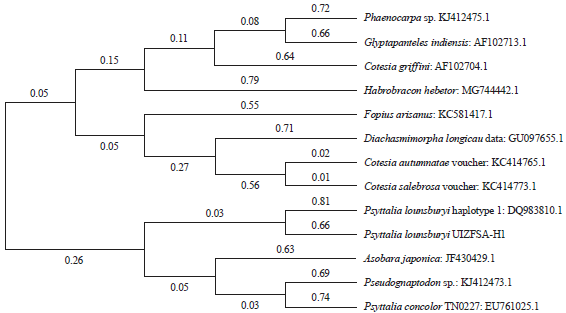 | |
| Fig. 4: | Evolutionary relationships of taxa. The evolutionary history was inferred using the Neighbor-Joining method33. The optimal tree with the sum of branch length = 9.29029587 is shown (Next to the branches). The evolutionary distances were computed using the Jukes-Cantor method and are in the units of the number of base substitutions per site. The analysis involved 13 nucleotide sequences34. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated. There were a total of 563 positions in the final dataset. Evolutionary analyses were conducted in MEGA631 |
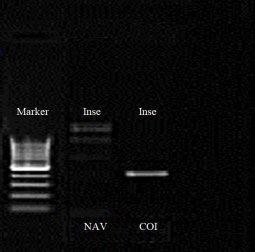 | |
| Fig. 5: | PCR amplification results of insect DNA with COI-F primer and NAV-F primer, the positive result was clearly observed with the COI-F primer |
The identification results revealed that this species belongs to the genus Psyttalia (Opius) from the Braconidae family, subfamily Opiinae know by their larval parasitism of other insects such as fruit flies.
Species occurrence and distribution: Adults of this Psyttalia sp. were observed in all boxes of different samples collected in the studied localities. The sampling site factor influenced significantly the three parameters studied (Table 2). Indeed, variable parasitism rate was detected in C. capitata larvae obtained from all the studied sampling sites (Fig. 4). The highest parasitism (%) was observed in “Ihchach site” with 21.5% followed by “Amskroud site” with 5.5% then Admin forest with 5% of parasitism by the Psyttalia sp. In addition, these highest percentages were observed in localities near to the agricultural area. Natural mortality was also observed and could be explained by the effect of temperature, conditions inside boxes or even by the parasitism.
Molecular analysis: The PCR showed positive amplification results of the COI-F/COI-R primers with the insect’s extracted DNA (Fig. 5). The analysis of the obtained DNA sequences using BLAST search revealed that these specimens were 92% (E-value = 2e-85) identical to the Psyttalia lounsburyi (Family Braconidae, subfamily Opiinae) haplotypes 1, 2 and 3 Cytochrome Oxidase Subunit I (COI) genes with the corresponding Genbank accession DQ983810.1, DQ983811.1 and DQ983812.1, respectively. The phylogenetic tree obtained from comparison of the specimen’s generated DNA sequences with other species from the Opiinae subfamily demonstrated the relationship with specimens (Psyttalia lounsburyi UIZFSA-H1) and Psyttalia lounsburyi species (Fig. 6).
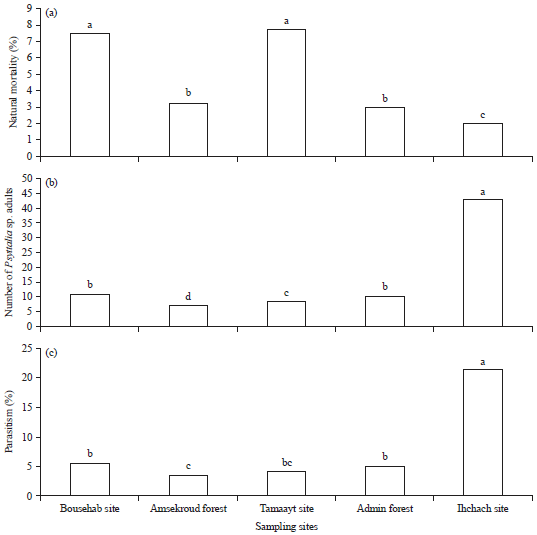 | |
| Fig. 6(a-c): | Observation of (a) Percentage of natural mortality, (b) Number of Psyttalia sp. adults and (c) Percentage of parasitism in the 5 sampling sites of the argane forest, Homogeneous groups (a-d) obtained by means comparison method |
| Least Significant Difference (LSD) at an alpha significance level of 5% | |
| Table 2: | Analysis of variance of 3 characters observed at the 5 sampling sites (Bousehab, Amskroud forest, Tamaayt, Admin forest and Ihchach) |
 | |
| **Significant at 0.05 | |
DISCUSSION
Several adults of larval parasitoid were observed among individuals emerged of the fruit fly in all boxes. These individuals are from samples collected in the localities studied in the Souss region of Morocco. This parasitoid was morphologically and molecularly identified using a universal primer pair COI-F/COI-R. Sampling of the host larvae (C. capitata) from several localities proved the attack of this pest by the parasitoid in all the studied samples with parasitism rates ranged from 3.5-21.5%. The parasitoid occurrence seems to be higher in localities near to agricultural areas where the pest is more abundant. Indeed, the parasitoid population levels are directly affected by the presence of the host insect. Which parasitic activity was also noticed in the forest mainly at the level of trees bearing argane fruits. Monitoring of population dynamics of the fruit fly and its natural enemy in the field seems to be a necessity in order to recommend methods of controlling this pest in the future.
Regarding the identification of this parasitoid, morphological criteria are identical to fruit fly’s egg-larval parasitoid belonging to Psyttalia (Opius) genus, family Braconidae, subfamily Opiinae. Starting with C. capitata larvae and pupae in boxes and obtaining as results parasitoid individuals after adult emergence confirms the endoparasitism of medfly larvae. In this type of parasitism, females lay their eggs inside larvae of their host. It attacks first-instar larvae of the fruit fly14,16,22. Indeed, a major biological control effort against fruit flies was carried out using several egg larval parasitoids from of the Opiinae subfamily11,12,16,19. One of the most successful examples of using natural parasitoids to control fruit flies including C. capitata and Bactrocera oleae was in Hawaii by introducing about 20 parasitoids including one egg-larval parasitoid, Fopius arisanu (Sonan) and seven larval parasitoids: Fopius vandenbosch, Diachasmimorpha tryoni, Diachasmimorpha longicaudata, Psyttalia fletcheri, Procecidochares incise, Psyttalia artun and Tetrastichus givardianus (Silvestri)12,17-20. However, those parasitoids seem not to be specific to the medfly and could attack non-target and beneficial insects. In such situations, the choice of specific parasitoids is essential in order to succeed this biological control method and therefore control pest populations. Recently, many works were carried out to test Fopius ceratitivorus (Wharton) in fields as a new specific parasitoid to medfly13,30,35. These studies are clearly demonstrated that this parasitoid attacks successfully eggs and larvae of C. capitata with the advantage of being specific to this pest, it appears unlikely to attack non-target Tephritidae in Hawaii14,15.
The DNA sequencing using COI-F/COI-R primers revealed 92% of similarity to P. lounsburyi species COI gene sequence. This result was confirmed by the Maximum-Likelihood phylogenetic analysis. Morphological criteria of our species such as body color are different with description of P. lounsburyi species22 and P. lounsburyi (Silvestri) which are known by a dark body color with a black pigmentation on the mesosoma (thorax) and metasoma21,23. The observed species was orange with no black spots, this color is similar to Psyttalia concolor. Several researches showed that variation in color has been observed in P. lounsburyi during his introduction from Africa to Hawaii in a great biological control program21,36. Approximately 50 species of Psyttalia genus are described, all are native of South and Central Africa21,22,30. Due to the morphological similarity of Psyttalia species researchers confronted major difficulty in establishing their taxonomic identities based on morphology criteria only22,32. Thus, various names have frequently been treated as synonyms or as separate species or confused with other species26,29. The DNA sequencing data in the present study using BLAST and phylogenetic tree showed that our specimens are genetically near to P. lounsburyi but not to Psyttalia concolor or Fopius arisanus. Psyttalia concolor has been reported in Morocco in a study of the specific wealth of natural enemies of C. capitata associated to citrus orchards37 from 2001-2011. However, the study37 didn’t use DNA sequencing for identification and therefore, never mentioned the occurrence of any P. lounsburyi species in Morocco. No further studies were found in this context which confirms that the first report of this larval parasitoid in Morocco and in North Africa. Psyttalia lounsburyi, the subject of this study, is a larval parasitoid probably native of Kenya, southern and central Africa11,21. This insect has often been introduced from Africa into many agricultural areas around the world to control olive fruit fly Bactrocera oleae. The most successful biological control programs using this hymenoptera were against B. oleae in California, USA24,25 and France28. Comparing to other Psyttalia parasitoids which are not selective for their hosts, P. lounsburyi has been reported for been specific for B. oleae11,22,30 and C. capitata24,26. The parasitism rates of C. capitata observed on a small sample of 400 larvae are very promising. Mass collect of argane infested fruits with C. capitata could be a solution to have more parasitoid and therefore mass produce it under controlled conditions. This parasitoid could be a good candidate as biological agent against C. capitata and B. oleae if it’s mass produced with flooding releases following the cycle of its host in the fields in Morocco and in the Mediterranean region. Its different morphology suggests that this species could be a new haplotype of P. lounsburyi and probably native to Morocco.
CONCLUSION
To minimize the harmful impact of pests on citrus production, using parasitoid as this new haplotype of P. lounsburyi would be an effective alternative for controlling C. capitata populations. Monitoring of population dynamics of the fruit fly and its natural enemy in the field seems to be a necessity in order to recommend methods of controlling medfly pest in the future.
SIGNIFICANCE STATEMENT
This study is the first report of a new haplotype of Psyttalia lounsburyi as larvae parasitoid of medfly which is considered as one of the most economically destructive pest of wide range of fruits and vegetables. It will be a very promising alternative to reduce the use of pesticides and ensure phytosanitary protection, respectful of environment and human health. This study will also allow researchers to focus their research on optimizing the conditions of mass production in order to recommend strategies to control medfly populations by inundative or augmentive releases depending on the availability of host under field conditions.
ACKNOWLEDGMENTS
We gratefully acknowledge the Moroccan Ministry of Scientific Research, the presidency of Ibn Zohr University for financial support. Our thanks to administration, technical service of CNRST (National Center for Scientific and Technical Research) for their technical assistance and molecular analyzes. We thank the anonymous reviewers and office journal which provided helpful comments that greatly improved the manuscript.
REFERENCES
- Gasperi, G., M. Bonizzoni, L.M. Gomulski, V. Murelli, C. Torti, A.R. Malacrida and C.R. Guglielmino, 2002. Genetic differentiation, gene flow and the origin of infestations of the medfly, Ceratitis capitata. Genetica, 116: 125-135.
Direct Link - Rachid, E. and M. Ahmed, 2018. Current status and future prospects of Ceratitis capitata Wiedemann (Diptera: Tephritidae) control in Morocco. J. Entomol., 15: 47-55.
CrossRefDirect Link - Alaoui, A., A. Imoulan, Z. El Alaoui-Talibi and A. El Meziane, 2010. Genetic structure of Mediterranean fruit fly (Ceratitis capitata) populations from Moroccan endemic forest of Argania spinosa. Int. J. Agric. Biol., 12: 291-298.
Direct Link - Morales, P., M. Cermeli, F. Godoy and B. Salas, 2004. A list of Mediterranean fruit fly Ceratitis capitata Wiedemann (Diptera: Tephritidae) host plants based on the records of INIA-CENIAP museum of insects of agricultural interest. Entomotropica, 19: 51-54.
Direct Link - Bernard, J.L., 2014. Face aux ravageurs, les solutions de lutte directe. Phytoma-La Défense Des Végétaux, 675: 9-14.
Direct Link - FAO. and IAEA., 1995. Economic evaluation of damage caused by and methods of control of the Mediterranean fruit fly in the Maghreb: An analysis covering three control options, including the sterile insect technique. Report of an Expert Group Organized by the Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture.
- Dimbi, S., N.K. Maniania, S.A. Lux, S. Ekesi and J.K. Mueke, 2003. Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin, to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathology, 156: 375-382.
CrossRefPubMedDirect Link - Quesada-Moraga, E., A. Ruiz-Garcia and C. Santiago-Alvarez, 2006. Laboratory evaluation of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against puparia and adults of Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol., 99: 1955-1966.
CrossRefPubMedDirect Link - Wang, X.G. and R.H. Messing, 2003. Intra-and interspecific competition by Fopius arisanus and Diachasmimorpha tryoni (Hymenoptera: Braconidae), parasitoids of tephritid fruit flies. Biol. Control, 27: 251-259.
CrossRefDirect Link - Bokonon-Ganta, A.H., M.M. Ramadan and R.H. Messing, 2007. Reproductive biology of Fopius ceratitivorus (Hymenoptera: Braconidae), an egg-larval parasitoid of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Biol. Control, 41: 361-367.
CrossRefDirect Link - Bokonon-Ganta, A.H., M.M. Ramadan, X.G. Wanga nd R.H. Messing, 2005. Biological performance and potential of Fopius ceratitivorus (Hymenoptera: Braconidae), an egg-larval parasitoid of tephritid fruit flies newly imported to Hawaii. Biol. Control, 33: 238-247.
CrossRefDirect Link - Wang, X.G., A.H. Bokonon‐Ganta, M.M. Ramadan and R.H. Messing, 2004. Egg-larval opiine parasitoids (Hym., Braconidae) of tephritid fruit fly pests do not attack the flowerhead‐feeder Trupanea dubautiae (Dipt., Tephritidae). J. Applied Entomol., 128: 716-722.
CrossRefDirect Link - Benelli, G., G. Gennari and A. Canale, 2013. Host discrimination ability in the tephritid parasitoid Psyttalia concolor (Hymenoptera: Braconidae). J. Pest Sci., 86: 245-251.
CrossRefDirect Link - Bess, H.A., R. van den Bosch and F.H. Haramoto, 1961. Fruit fly parasites and their activities in Hawaii. Proc. Hawaiian Entomol. Soc., 17: 367-378.
Direct Link - Vargas, R.I., W.A. Walsh and T. Nishida, 1995. Colonization of newly planted coffee fields: Dominance of Mediterranean fruit fly over oriental fruit fly (Diptera: Tephritidae). J. Econ. Entomol., 88: 620-627.
CrossRefDirect Link - Wong, T.T. and M.M. Ramadan, 1987. Parasitization of the Mediterranean and oriental fruit flies (Diptera: Tephritidae) in the Kula area of Maui, Hawaii. J. Econ. Entomol., 80: 77-80.
CrossRefDirect Link - Fischer, M., 1971. Zwei von Uganda nach Hawaii importierte Opius-Arten (Hymenoptera, Braconidae). Anzeiger Schädlingskunde Pflanzenschutz, 44: 10-12.
CrossRefDirect Link - Wharton, R.A. and F.E. Gilstrap, 1983. Key to and status of opiine braconid (Hymenoptera) parasitoids used in biological control of Ceratitis and Dacus sl (Diptera: Tephritidae). Ann. Entomol. Soc. Am., 76: 721-742.
CrossRefDirect Link - Wharton, R.A. and P.M. Marsh, 1978. New world opiinae (Hymenoptera: Braconidae) parasitic on tephritidae (Diptera). J. Washington Acad. Sci., 68: 147-167.
Direct Link - Daane, K.M., K.R. Sime, X.G. Wang, H. Nadel and M.W. Johnson et al., 2008. Psyttalia lounsburyi (Hymenoptera: Braconidae), potential biological control agent for the olive fruit fly in California. Biol. Control, 44: 79-89.
CrossRefDirect Link - Wang, X.G., M.W. Johnson, V.Y. Yokoyama, C.H. Pickett and K.M. Daane, 2011. Comparative evaluation of two olive fruit fly parasitoids under varying abiotic conditions. BioControl, 56: 283-293.
CrossRefDirect Link - Billah, M.K., S. Kimani-Njogu, W.A. Overholt, R.A. Wharton, D.D. Wilson and M.A. Cobblah, 2005. The effect of host larvae on three Psyttalia species (Hymenoptera: Braconidae), parasitoids of fruit-infesting flies (Diptera: Tephritidae). Int. J. Trop. Insect Sci., 25: 168-175.
Direct Link - Giunti, G., G. Benelli, R.H. Messing and A. Canale, 2016. Early adult learning affects host preferences in the tephritid parasitoid Psyttalia concolor (Hymenoptera: Braconidae). J. Pest Sci., 89: 529-537.
CrossRefDirect Link - Kimani-Njogu, S.W., M.K. Trostle, R.A. Wharton, J.B. Woolley and A. Raspi, 2001. Biosystematics of the Psyttalia concolor species complex (Hymenoptera: Braconidae: Opiinae): The identity of populations attacking Ceratitis capitata (Diptera: Tephritidae) in coffee in Kenya. Biol. Control, 20: 167-174.
CrossRefDirect Link - Wharton, R.A., 1999. A review of the old world genus fopius wharton (Hymenoptera: Braconidae: Opiinae), with description of two new species reared from fruit-infesting Tephritidae (Diptera). J. Hymenoptera Res., 8: 48-64.
Direct Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link - Lopez, M., J. Sivinski, P. Rendon, T. Holler and K. Bloem et al., 2003. Colonization of Fopius ceratitivorus, a newly discovered African egg-pupal parasitoid (Hymenoptera: Braconidae) of Ceratitis capitata (Diptera: Tephritidae). Florida Entomol., 86: 53-60.
CrossRefDirect Link - Smaili, M.C., M. Abbassi, J.A. Boutaleb and A. Blenzar, 2013. Richesse spécifique des ennemis naturels associés aux vergers d'agrumes au Maroc: Intérêt et implication pour la lutte biologique. EPPO Bull., 43: 155-166.
CrossRefDirect Link