
Clara de Castro
Sayre School, Upper School, 194 North Limestone, Lexington, 40507 KY, USA
Josh Titlow
Department of Biochemistry, University of Oxford, Oxford, United Kingdom
Zana R. Majeed
Department of Biology, University of Kentucky, 675 Rose Street, Lexington, 40506-0225 KY, USA
Cole Malloy
Department of Biology, University of Kentucky, 675 Rose Street, Lexington, 40506-0225 KY, USA
Kayla E. King
Department of Biology, University of Kentucky, 675 Rose Street, Lexington, 40506-0225 KY, USA
Robin L. Cooper
Department of Biology, University of Kentucky, 675 Rose Street, Lexington, 40506-0225 KY, USA
LiveDNA: 1.523
Journal of Entomology
Year: 2019 | Volume: 16 | Issue: 2 | Page No.: 62-73







ABSTRACT
Background and Objective: Cardiac-intrinsic mechanisms for regulating heart rate (HR) in response to stretch have been characterized in many different organisms but the molecular mechanisms for mechanosensitive regulation are not yet fully understood. Drosophila melanogaster larva, with its simple and genetically tractable circulatory system, is well-suited to investigate the functional role of stretch-activated channels (SACs) and chemical modulators in regulating heart rate. Materials and Methods: Here we establish an exposed heart preparation that delivers continuous mechanical stimulation in the presence of pharmacological manipulations. Results: Continuous mechanical stimulation substantially stabilized HR over 2 h incubations compared to stagnant incubations with the same saline. The addition of a chemical modulator cocktail further enhanced HR stability in perfused preparations. To determine the role of mechanosensitive regulation in prolonged HR stability we assayed various pharmacological SAC blockers, e.g., amiloride, gadolinium, ruthenium red, streptomycin and tarantula toxin. Conclusion: The relative efficacy of these blockers on mechanically stabilized HR further establishes Drosophila larva as an appropriate system for investigating cardiac mechanosensation and provides novel insight into the underlying SACs.
PDF Abstract XML References Citation
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Clara de Castro, Josh Titlow, Zana R. Majeed, Cole Malloy, Kayla E. King and Robin L. Cooper, 2019. Mechanical and Chemical Factors Required for Maintaining Cardiac Rhythm in Drosophila melanogaster Larva. Journal of Entomology, 16: 62-73.
DOI: 10.3923/je.2019.62.73
URL: https://scialert.net/abstract/?doi=je.2019.62.73
DOI: 10.3923/je.2019.62.73
URL: https://scialert.net/abstract/?doi=je.2019.62.73
INTRODUCTION
Despite obvious differences in heart morphology and function the mechanosensitivity of cardiac tissue appears to be fundamentally similar across vertebrate species1. It has been suggested that lower vertebrate and invertebrate circulatory systems rely more heavily on this form of chronotropic regulation because of their more primitive neurohumoral system2. Yet there are extensive examples of chemical modulators that have chronotropic effects in, for example, Drosophila melanogaster larva3-6, which has also been shown to exhibit mechanically induced changes in heart rate (HR)7. How cardiac pacemaker cells simultaneously process mechanical and chemical input remains unknown in any organism.
With its genetic and experimental tractability the Drosophila larva heart provides a powerful system for defining the molecular mechanisms involved in cardiac mechanosensation and chemical modulation. The morphology of Drosophila and mammalian hearts is different, yet the molecular mechanisms that underlie heart development share a large degree of similarity8, particularly regarding evolutionarily conserved transcription factors and signaling pathways8-10. Moreover, the hearts are functionally indexed by similar physiological measurements, e.g., cardiac output, rate and time in systole or diastole11. Therefore studies in fruit fly heart biology are likely to provide valuable insight into the mechanisms of mammalian heart function.
The larval heart consists of a dorsal vessel with the anterior aspect of the tube serving as an aorta12. Drosophila melanogaster larval heart is myogenic and is bathed by the hemolymph so any circulating modulator can have a direct effect on the entire heart physiology9,13,14. With recent development of a modified saline that maintains the exposed heart for hours15, it is now possible to perform more detailed electrophysiological and cellular studies on the mechanisms regulating ionotropic and chronotropic actions. The physiological saline commonly used for measuring synaptic responses and CNS activity in Drosophila larvae did not maintain heart rhythm over such long durations because of drift in pH15.
To progress our knowledge of HR modulation in Drosophila larva we have started to address how hormones/modulators act on the heart through various receptor subtypes3-6. There are many peptides and hormones within the hemolymph of the animal that can modulate HR, so it is a daunting task to address the role of all of them in isolation and in various combinations. We begin by examining the effect of modulator cocktails on HR. Comparing the effects of single compounds with combined effects provides novel insight into how interactions between signaling cascades modulate HR in the intact animal.
When conducting previous studies on dissected larvae we noted that simply exchanging the bath saline with fresh saline produced a slight but consistent increase in HR. This is likely a response due to stimulation of stretch-activated ion channels (SACs), which are known to be expressed in mammalian hearts16 and are present in Drosophila sensory neurons17. The TRPA channel, Painless, is also present in the larval Drosophila heart7 where it may be responsible for the pauses in contraction of the heart during larval crawling. This particular channel might also sense temperature as well as mechanical stress. The mechanical deformation on the heart and potentially SACs can occur within the intact larvae by the pull of the alary muscles around the heart and the stress on the cells with the longitudinal pull during contraction of the cardiac tube as well as locomotion7. It appears that there are SACs that can stimulate as well as depress the cardiac pacemaker potentials possibly depending on the degree of stretch on the cardiac tissue.
In this study we developed a mechanical stimulation assay to determine the role of various SACs in mechanosensitive HR modulation. Using this assay we tested a panel of pharmacological agents (amiloride, ruthenium red, gadolinium (Gd3+), streptomycin and the tarantula toxin GsMTx-4), which are known to effect some forms of SACs. We examined if these compounds might also alter the function of SAC subtypes in the larval heart18-23.
MATERIALS AND METHODS
Fly strains and maintenance: The fly strains used were the common ‘wild-type’ laboratory strain Canton S and transgenic lines expressing Piezo dsRNA. For RNAi knockdown of Piezo, virgin females fromw; Kr/CyO; Gal4-Mef2, UAS-mCD8.mRFP (stock #26882). The RFP were crossed with UAS-Piezo-RNAi (RFP expressing larva were selected for the experiments since they have both UAS and Gal4 constructs) and males of 24B-Gal4/TM3 (non-stubble homozygous flies for the 24B-Gal4 construct were used because they are homozygous for the construct) were crossed with the virgin females of UAS-Piezo-RNAi line. The UAS-dpiezo-RNAi strain was obtained from the National Institute of Genetics (Japan, 8486R-3). The 24B-Gal4 and the Mef2-Gal4 strains were obtained from Bloomington Stock Center (# 27390; Bloomington, IN).
All larvae were maintained at room temperature ~21°C in vials partially filled with a cornmeal-agar-dextrose-yeast medium on a light:dark cycle 14:8.
Dissection and solutions: The general dissection technique used to directly assess heart rate in Drosophila larvae has been previously reported15. Dissection times were 3-6 min. Larvae were dissected in modified HL3 saline described in de Castro et al.15. The saline contained (mM): 1.0 CaCl2·2H2O, 70 NaCl, 20 MgCl2, 5 KCl, 10 NaHCO3, 5 trehalose, 115 sucrose, 25 5-N,N-bis(2-hydroxyethyl)-2-aminoethane sulfonic acid (BES). After aeration, the pH of all salines was adjusted to 7.1 with HCl (1 M) or NaOH (1 M) as needed. The saline pH was monitored at room temperature (21-23°C) using an Accumet model 10 pH meter (Fisher Scientific) and Ag/AgCl glass electrode. To examine the effect of modulators on HR, the bath saline was exchanged with a saline containing the compounds of interests. Modulator solutions (dopamine, octopamine, acetylcholine and serotonin) were prepared fresh daily from stocks of frozen aliquots. All chemicals listed above were obtained from Sigma-Aldrich (St. Louis, MO).
Pharmacological agents: Streptomycin, gadolinium trichloride-GdCl3, amiloride and ruthenium red were used at a concentration of 1 mM. This concentration was used since it was shown to be effective in blocking stretch activated ion channels (SACs)24. All chemical compounds were obtained from Sigma (St. Louis, MO, USA). A tarantula toxin (GsMTX-4) was also tested for its effects on the HR and was obtained from Alomone Labs (Jerusalem BioPark, Israel).
Heart rate measurements: Heart rate (HR) was counted by direct observation through a dissecting microscope. Experiments were performed either with or without pulsatile saline perfusion as mentioned in the text. A continuous perfusion pump (Master Flex, Vernon Hills, IL) with a flow rate of 1.6 mL min–1 was used for experiments in which perfusion was utilized. With the small recording dishes this rate of flow produces a substantial disturbance of fluid around the heart. For a visualization of the recording dish and dissection procedures see Cooper et al.25. To examine the effects of different compounds the bathing medium was exchanged. The exchange times are indicated in the figures as they varied depending on the experimental paradigm.
Raw heart rate values are presented to illustrate the variability obtained in these preparations for the various conditions. The rationale for showing raw values is if a low value occurs initially with an increase, a larger percent change is observed than for a larger initial value increasing the same number of beats per minute. Past literature representing heart rates in insects has made it difficult to compare conditions due to only presenting percent changes and the lack of presenting raw data with experimental conditions.
RESULTS
Effects of perfusion on heart rate (HR) stability in dissected larvae: A previous study15 demonstrated that dissected larva heart preparations could be maintained with a consistent HR for 10-20 min by optimizing pH in the bath saline. For longer durations the HR decreased even under optimal pH conditions. To maintain constant pH and ion composition, which could fluctuate due to evaporation, the bathing media was exchanged when HR decreased. It was noted that HR increased each time the static bathing media was exchanged, leading us to hypothesize that mechanical agitation elevates HR. To address this we first quantified HR in response to periodic mechanical agitation (i.e., exchanging the bath solution every 30 min). Without perturbation the HR began to decrease within 30 min of dissection (Fig. 1a). This decrease is not due to an alteration in pH as the pH of HL3 saline with 25 mM BES buffer is stable for several hours15. When the bath solution was exchanged after 30 min incubation, 4/6 preparations exhibited elevated HR, one preparation maintained stable HR and the last preparation did not respond (Fig. 1a). Saline changes after 60 and 90 min incubations caused similar HR elevations in 4/6 preparations, suggesting that the larval heart responds to tactile stimulation. The results are also presented as an average of the percent change (±SEM) from the initial values for each preparation (right side panels, Fig. 1a-c).
To further test our hypothesis that mechanical stimulation of the heart causes rapid HR elevation we designed an apparatus to systematically stretch the larval body wall (which attaches to the heart through alary muscles and trachea that stretch the heart during crawling). This was achieved by connecting the moveable dissection pin at the tail end of the larva to an audio speaker controlled by a DC voltage source. Vibration of the speaker was calibrated to shorten and elongate the larval body wall by ~0.5 mm at 0.5 Hz. Continuously stretching larvae with this apparatus stabilized the HR for up to 2 h (Fig. 1b, T-test p<0.05) between static bath and with use of the stretching device at 120 min.
However, stretching the larvae with this method caused the cuticle to tear and is therefore not an ideal system. Instead we found that continuous saline perfusion with a strong pulsatile flow can produce a similar disturbance that stretches the heart tube. This procedure also produced stable HRs in 6/6 preparations for 2 h without damaging the heart or skeletal muscles (Fig. 1c, T-test p<0.05) between static bath and with use of the perfusion at 120 min. Together these results show that HR is elevated by mechanical stimulation.
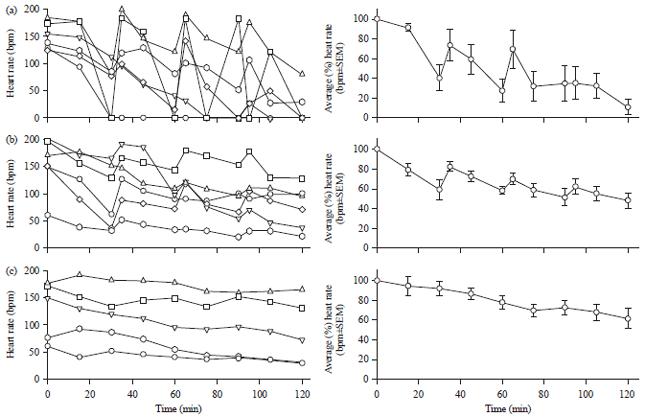 | |
| Fig. 1(a-c): | Mechanical stimulation stabilizes heart rate (HR) in dissected Drosophila larva preparations, (a) HR measurements were acquired every 15 min for 6 separate preparations over a 2 h period. New saline was added after each measurement and HR was recorded again after 2 min. HR began to decay within 30 min in every preparation and continued to decline over the 2 h regime, though intermittent saline exchanges briefly stabilized HR, (b) Mechanically stretching the preparations in 2 sec intervals stabilized HR for 6/6 preparations for the entire 2 h experiment and (c) Continuous pulsatile perfusion, which also provides a constant mechanical stimulation, stabilized HR in 5/5 preparations |
Panels on the right are the averages in the percent changes from the first measure for each preparation | |
Examining the role of stretch-activated (SACs) channels in stabilizing larval HR: To determine the molecular mechanism through which mechanical force elevates HR we investigated the role of stretch-activated channels (SACs). The SACs are a broad family of ion channel proteins that are currently being characterized by gene/protein sequencing and pharmacological profiles26. Drosophila melanogaster genome contains genes for TRP channels (Transient Receptor Potential channels), DEG/ENaCs (Degenerin/epithelial sodium channels) and Piezo (pressure sensitive channel)17,27. We obtained a transgenic RNAi line targeting dPiezo and used the Gal4-UAS binary expression system28 to express Piezo-dsRNAi in the heart. Two different Gal4 driver lines were crossed to the UAS-Piezo RNAi line and HRs were measured in dissected preparations with pulsatile perfusion. The first line (24B>dPiezo RNAi) showed normal elevated responses at the beginning of perfusion (5 min, Fig. 2a) and 4/6 preparations maintained stable HR for the 2 h observation period. In the second line (Mef-2>dPiezo RNAi), only 4/8 preparations showed an elevated response at the beginning of perfusion (5 min, Fig. 2b) and only 4/8 preparations maintained a stable HR over the 2 h observation period. The results are also presented as an average of the percent change (±SEM) from the initial values for each preparation (right side panels, Fig. 2a, b). There was no statistically significant difference between normalized HR at 120 min in Mef-2>dPiezo RNAior the 24B>dPiezo RNAi lines compared to WT. These results show that Piezo knockdown has at most a mild effect on mechanically induced HR stabilization and suggests that other SACs may mediate the stimulatory effects of mechanical stimulation on HR.
To determine which SACs are likely to function in the larval heart we employed well-characterized pharmacological effectors of SACs.
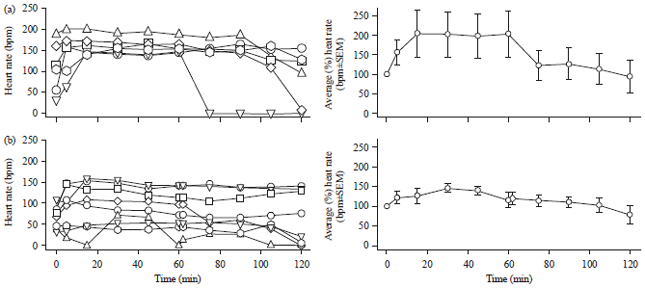 | |
| Fig. 2(a-b): | Knockdown of dPiezo has only a mild inhibitory effect on mechanically induced HR stability in dissected larva preparations, (a) Expressing dPiezo RNAi in the heart with the 24B-Gal4 driver does not interfere with rapid (1 min) elevation of HR in response to pulsatile perfusion, though 2/6 preparations did not maintain stable HR for the entire 2 h experiment and (b) Rapid and prolonged effects of pulsatile perfusion were inhibited in 4/6 preparations when dPiezo RNA was expressed in the heart using Mef2-Gal4 |
Panels on the right are the averages in the percent changes from the first measure for each preparation | |
Drugs that were tested include amiloride, gadolinium-trichloride (Gd3+), ruthenium red, streptomycin and tarantula toxin (GsMTx4). Both amiloride and ruthenium red are known to block a subset of SACs18,21. Some receptor subsets associated with the TRP family are known to be blocked by ruthenium red, while the DEG/ENaC channels are blocked by amiloride. There are subgroups of DEG/ENaC channels that are referred to as acid sensitive stretch activated channels (ASC) since they demonstrate an increased activation in the presence of a low pH environment23. Some subgroups of ASC are also blocked by amiloride21. The Drosophila Piezo channels are comprised of a distinct type of protein sequence that is conserved among species18. The mammalian form is blocked by ruthenium red while the Drosophila form is not18. The Piezo associated channels in dorsal bipolar dendritic neurons of larval Drosophila are blocked by amiloride29. The Gd3+ is a non-selective SAC inhibitor that is known to block a variety of SACs, some of which are unique in pharmacology20. Streptomycin is an antibiotic known to block SACs in sarcolemma of mammalian hearts22,30. Finally, GsMTx4 is a peptide from tarantula venom that blocks cationic SACs and is suggested to act on the extracellular side of the channel, meaning saline containing GsMTx4 would have access to these types of SACs31,32.
To examine the effects of amiloride the dissected preparations were first perfused with normal saline for 15 min to record the baseline HR. After measuring HR at 15 min the bathing solution was exchanged for a solution containing amiloride (1 mM). Within 90 min the larval hearts stopped contracting in 5/5 preparations (Fig. 3a, p<0.05 T-test). Preparations that were incubated in the control saline alone maintained stable HRs for the entire 90 min (Fig. 3b). A non-selective SAC inhibitor, Gd3+, exhibited an even stronger effect on larval HR. Hearts stopped beating after only 10 min incubation in Gd3+ (10 μM) in 5/5 preparations. After rapid exchange of the saline back to normal saline, the preparations partially recovered but did not sustain a stable HR (Fig. 3c, p<0.05 T-test).
The cationic SAC inhibitor, GsMTx4, had a less severe depressive effect on HR. After 60 min exposure to pulsatile perfusion of GsMTx4 (15 μM), HR decayed to 0 BPM for 4/6 preparations (Fig. 3d, p<0.05 T-test) at 30 min compared to control (Fig. 3b). Interestingly, though hearts were unable to maintain stable contractions for long durations in GsMTx4, increasing the force of pulsatile perfusion over the hearts after contractions stopped (60 min) restarted the heart in 4/4 preparations.
Incubating dissected larvae in ruthenium red or streptomycin had no measurable effect on HR. Under constant pulsatile perfusion with ruthenium red (1 mM) 6/6 preparations maintained a stable HR over the 2 h incubation (Fig. 3e). Similarly, 6/6 preparations continuously perfused with streptomycin (1 mM) maintained steady HR for up to 2 h (Fig. 3f).
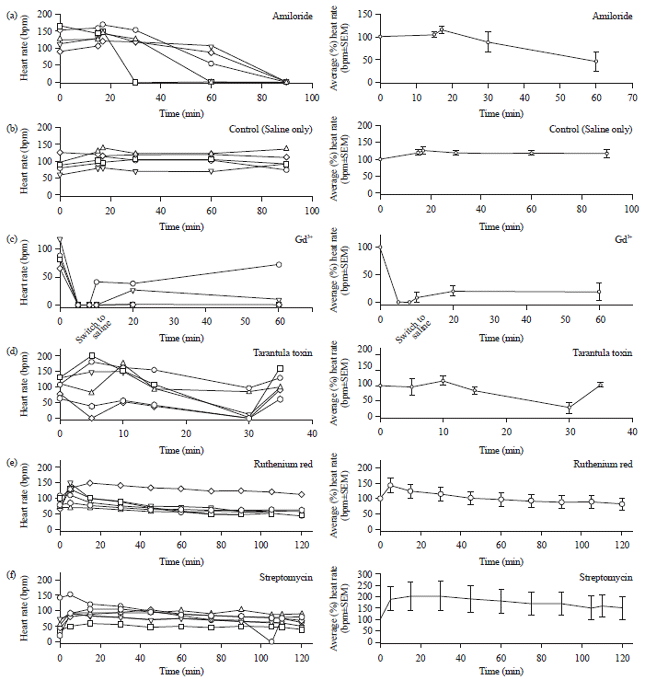 | |
| Fig. 3(a-f): | Effects of varying agents on maintaining the heart rate, (a) Effect of amiloride on larval HR, perfusing amiloride over dissected larva hearts abolishes HR within 90 min, (b) HR is stable for this duration under perfusion of saline alone, (c) Perfusing gadolinium (10 μM) over dissected larva hearts abolishes HR within 30 min, (d) Perfusing tarantula toxin (GsMTX-4) (15 μM) over dissected larva hearts abolishes HR in 4/6 preparations within 30 min, increasing the pulsatile pressure at 30 min acutely resuscitated/elevated HR in 6/6 preparations, (e) Perfusing ruthenium red (10 mM) over dissected larva hearts does not inhibit the rapid (1 min) or prolonged (120 min) response to pulsatile perfusion and (f) Perfusing streptomycin (10 mM) over dissected larva hearts does not inhibit the rapid (1 min) or prolonged (120 min) response to pulsatile perfusion |
Panels on the right are the averages in the percent changes from the first measure for each preparation | |
The results are also presented as an average of the percent change (±SEM) from the initial values for each preparation (right side panels, Fig. 3a-f). The ruthenium red or streptomycin at 120 min is not significantly different from control or with saline perfusion on its own as shown in Fig. 1c.
Chemical modulation of larva HR: The ability of the larval heart to respond to modulators is well documented. Depending on how compounds are introduced by injection, feeding or directly to hearts exposed to saline containing the compounds in the absence of hemolymph, varying results have been obtained. It is hard to determine the makeup of the active substances contained within the hemolymph within intact larvae and how they may interact. Thus, a reductionist approach of removing other variables and using a defined saline with known concentrations of substances and pharmacological agents has allowed the various receptor subtypes on the heart to be functionally characterized. Serotonin (5-HT), acetylcholine (Ach) and dopamine (DA) are known to have chronotropic effects on the larval heart. Responses to known agonist and antagonists have helped to classify the functional receptor subtypes for these modulators3-6. However, how the heart responds to cocktails of these modulators has not been addressed. Octopamine (OA) has also been reported to play a role in regulating Drosophila HR33, therefore we decided to include it in the cocktail after characterizing its effects in the dissected preparation.
To assess the actions of OA on the exposed heart a range of concentrations were examined. HR was immediately elevated in response to OA concentrations as low as 10 nM and a maximal response at 100 nM so a 100 nM was used for this assessment over time (Fig. 4, p<0.5 T-test). The effect of exchanging the saline on the exposed heart cause a transitory increase in HR and has been documented in previous studies3-6. The mechanical perturbation of exchanging the bath saline for saline in exposed preparations results in a 10-15% increase in HR that decays within 1-5 min. Thus, when assessing chronotropic effects in this assay we measure the rapid response (2 min) and the prolonged response (10 min). For a molecule to be considered a bona fide modulator with a positive chronotropic effect it must elevate HR significantly above the 5-15% sham threshold and/or maintain an elevated HR for 10 min. The HR was initially elevated by 40% in response in to 100 nM OA and the HR did not decay within 10 min (Fig. 4, no significant change after 10 min), therefore OA has a positive chronotropic effect on larva HR.
Next the effect of combining OA and 5-HT, the modulator which has the next largest effect on larva HR, was analyzed by applying equal concentrations (10 nM) of the modulators and measuring the rapid (2 min) and prolonged HR response (10 min). The graph in Fig. 5a shows the measured HR at each time point in the experiment. First, application of OA (10 nM) causes a rapid 40% elevation in HR (p<0.05, paired T-test). Applying 5-HT (10 nM) at this point caused a slight but significant elevation in HR that persisted for 10 min (p<0.05, paired T-test).
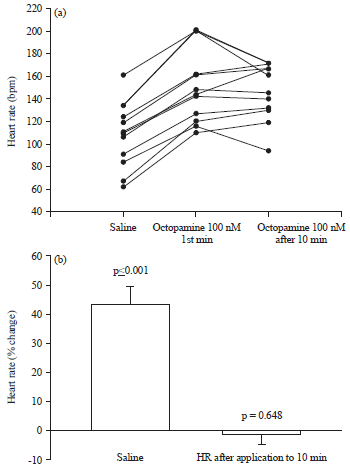 | |
| Fig. 4(a-b): | Effect of octopamine (OA) on larva HR, (a) Acute (1 min) and prolonged (10 min) responses to OA (100 nM) were measured in dissected larva heart preparations. HR rapidly increased in 11/11 preparations when OA was added and the elevated HR was sustained for 10 min and (b) Summary statistics comparing the rapid HR change after OA application to sham saline treatments (left) and comparing the HR at 1 min after application to 10 min after application (right), showing that the HR was significantly elevated immediately after OA application and did not decrease significantly after 10 min |
No significant changes occurred after 10 min of combined incubation. Summary statistics for this experiment are shown in Fig. 5b. This experiment was then repeated with increasing concentrations of both modulators. The rapid effect of OA on HR does not increase at higher concentrations but the additive effect of 5-HT does increase with concentration (Fig. 5c), indicating that separate signal transduction cascades mediate the effects of these modulators.
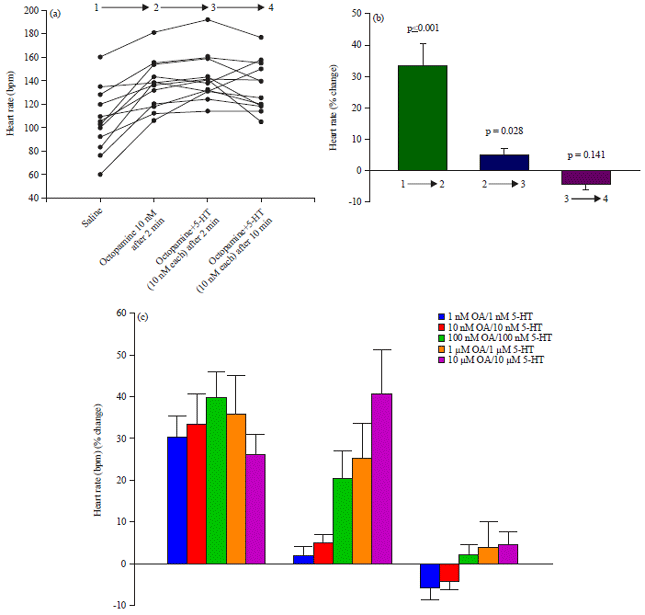 | |
| Fig. 5(a-c): | Combined effects of octopamine (OA) and serotonin (5-HT) on larval HR, (a) OA (10 nM) rapidly elevates HR 2 min after application in 12/12 preparations. Addition of 5-HT causes HR to increase further in 9/12 preparations and the elevated HR is sustained for at least 10 min, (b) Summary statistics for the initial change in HR immediately after OA application (green bar), the change in HR from OA incubation to OA+5-HT incubation (blue bar) and the change in HR from immediately after 5-HT addition to 10 min after 5-HT addition (violet bar) and (c) The same experiment was repeated over a range of OA and 5-HT concentrations. Summary statistics are the same as in (b). These results show that there are additive chronotropic effects between the two modulators |
Arrows represent the percent change calculated in top graph (a) between the time points | |
The effects of the OA and 5-HT cocktail was suggestive that they were not competing for second messenger cascades since exposure to 5-HT still resulted in large increases in HR in the presence of a saturating response to OA and the responses were maintained for 10 min with a static bathing environment. Based on these results we next tried a cocktail of various modulators known to influence larva HR (OA, 5-HT, DA and Ach all at 10 μM). Application of this cocktail caused a rapid elevation of HR that persisted for 2 h (Fig. 6, p>0.05, paired T-test as well as rank sum non-parametric). The results are also presented as an average of the percent change (±SEM) from the initial values for each preparation (see right side panel of Fig. 6).
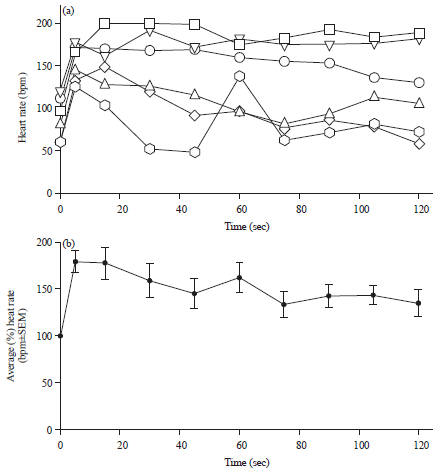 | |
| Fig. 6(a-b): | Effect of chemical modulator cocktail on larva HR, (a) A solution containing OA, 5-HT, DA and Ach (10 μM) was perfused over dissected larva hearts and (b) Average in the percent changes from the first measure for each preparation |
Note the rapid elevation in HR after applying the modulators (1 min) and the prolonged stabilization of HR in 6/6 preparations (120 min) | |
There is no difference from the trials with saline perfusion alone (Fig. 1c) to those with the cocktail added; however, the difference before adding the cocktail and 120 min after being in the cocktail is at a 93% difference (p = 0.0627, paired T-test) whereas the perfusion alone before and after 120 min is (p = 0.0184). The exposure to the cocktail resulted in a significant increase and the long term consequences of exposure to the stimulator effect may result in a greater decrease over time than if only a slight increase is caused by saline perfusion alone.
DISCUSSION
In this study we demonstrated that pulsatile perfusion of saline over the exposed Drosophila larva heart tube is more efficacious in stabilizing heart rate (HR) over long periods of time than a static bath. Pharmacological survey of known stretch-activated channel (SAC) blockers indicated that HR stabilization by mechanical stimulation was through a subset of SACs that are sensitive to amiloride, Gd+3 and to a lesser extent tarantula toxin (GsMtx4).
We also describe a modulator cocktail (OA, DA, 5-HT, Ach) that stabilizes HR for over 2 h in dissected larva preparations. Thus there is a possibility of developing a better saline condition for long term culture of Drosophila cardiac tissue. OA has an influence on HR at relatively lower concentrations than for 5-HT or DA that is maintained for at least several minutes, indicating that the effect is greater than the mechanical stimulation of changing the bathing media. The combination of OA and 5-HT, at equal concentrations over a wide range continued to demonstrate that 5-HT further increased HR while the effect of OA was saturated, thus indicating a weak cross talk between these two modulatory intracellular signaling pathways in cardiac cells.
The receptors identified in recent studies3-6 indicate various second messenger cascades that can alter the cardiac pacemaker cycle. The serotoninergic effects work through activation of the PLC-IP3-PKC pathway initiated by 5-HT2-like receptors on the larval heart3,34,35. The dopaminergic receptors (type-1 DA and type-2 DA) are likely GPCRs that work through activation of phospholipase-C and other second messenger cascades5,33. The cholinergic actions on the larval heart elevate HR through both nAChRs and mAChRs. The A-type mAChRs are likely the predominant receptor subtype and are coupled with the Gq/11 signaling pathway4,33,36. Octopamine’s excitatory action is also likely through GPCRs6,9, possibly through the Gαq subunit, however in cold conditions (10°C) OA decreased HR37 which was speculated that the action maybe via Gαi-coupled receptors6. Thus, the cocktail of modulators used in our study are probably elevating HR through parallel activation of intracellular signaling pathways rather than converging on any individual signaling cascade. Activation of parallel pathways sustains HR at a high level for at least the 2 h of observation.
In this study, RNAi-mediated knockdown of Piezo in mesodermal-originated tissues, with two different targeted lines, did not reveal a significant difference in HR with or without perfusion. It is likely that Piezo is not expressed in the larval heart or if it is, plays only a minor role in modulating the dynamics of larval heart rate. Piezo is a SAC that is expressed in dorsal bipolar dendritic neurons. These neurons detect tactile forces on the cuticle of the larvae and were shown to be blocked by amiloride29 but not by ruthenium red18. The Piezo channels are now known to be of a unique protein sequence and are similar among species from invertebrates to mammals; however, they are now considered a novel class of ion channels involved in mechanotransduction during stretch18. The possibility of various forms of SAC being expressed in the larval heart has not been fully addressed yet. The presence of various SACs genes in Drosophila genome indicate the possibility of different tissues expressing different forms of SACs. The SAC, Painless, has been shown to be expressed in the larval heart7. However, there appears to be a question if this TRP channel is more for thermal sensing or mechanosensory7. Although, the evidence is convincing that with the RNAi knockdown of TRP the larval hearts do not pause as often when the heart is stretched in intact crawling larvae. Interestingly, in our study, supra-perfusion of the saline, which was used as mechanical agitation, elevated HR. Stimulating SACs in the hearts of mammals is also shown to stimulate contraction16. The possibility that pacemaker cells use different SACs to increase or decrease HR has not been explored in any organism.
Since Painless is a TRP channel, which allows Ca2+ influx38, one might predict this would result in the heart maintaining a contracted state and not a relaxed paused state. Loading the heart with Ca2+ by blocking the Ca2+-ATPase (PMCA) as well as the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase (SERCA) or attenuating the plasmalemmal Na+/Ca2+ exchanger (NCX) increases larval HR in Drosophila39. Also, likely increasing Ca2+ loading within the myocytes by the action of 5-HT3 or bathing the exposed heart in higher Ca2+ in the bathing media or through activation by optogenetically driven channel rhodopsin (ie., Ca2+ channels) resulted in faster not slower HRs6,15,37. This role of ionic fluxes in the cardiac excitation-contraction cycle needs to be further investigated as a subtle difference in ionic flux may alter the cardiac electrical cycle that in turn would impact the observed contractions.
It is not surprising that GdCl3 caused the heart to rapidly cease from beating as Gd3+ is known to block Ca2+ channels40 and as stated above, altering extracellular Ca2+ has an effect on HR. So it is likely that Ca2+ influx has a role within the pacemaker cycle and that blocking the influx stopped the contractions independently from blocking any potential SACs. Streptomycin blocks SACs-even the Piezo channel41,42. However, streptomycin reduces the L-type Ca2+ current in guinea-pig heart at a 2 mM but showed no effect at 50 μM43. Given that 1 mM had no significant effects in our study would indicate that the Ca2+ channels were not compromised and that the SACs that might be present were also not affected. Similarly, GsMTx-4, which blocks Piezo channels and other types of SACs, had no significant effect on the HR with supra-perfusion18-23. This result would suggest that whatever form of SACs found on the heart are not sensitive to this selective toxin. One also needs be cautious when using the GsMTx-4 since it blocks Kv11.1 and Kv11.2 potassium channels44. Because the HR was not significantly altered, it appears that GsMTx-4 has no action on the larval heart.
When intact or dissected larval hearts are exposed to cold saline the HR drastically slows down. Any stimulatory effect in speeding up the HR such as through modulation may be even more noticeable since the rate is at its lower rate. A recent study from our group found that exchanging the bathing media over the heart exposed to cold (10°C) did not have a stimulatory effect compared to exchanging the media at room temperature (21°C)6. So the SACs that are expressed in the larvae heart and function in mechanically-induced HR elevation are not activated in cold conditions. One possibility is that SACs are unable to change conformations at lower temperatures. Another possibility is that stretch-induced conformational changes are dampened by cold shock proteins acting as chaperones to stabilize protein complexes in the lipid bilayer45.
CONCLUSION AND FUTURE RECOMMENDATIONS
It can be concluded that the Drosophila larva heart possesses a SAC(s) that intrinsically elevates HR in response to tactile stimulation. With pulsatile perfusion of saline containing a cocktail of common modulators and optimal pH the exposed larva heart can now be directly monitored for multiple hours.
Comprehensive identification of the mechanosensory channels and chemical receptors expressed in larva heart would provide a clearer picture of how pacemaker activity is regulated in cardiac cells. A single cell proteomic/ transcriptomic approach would be ideal for this experiment, as cardiac cells would need to be dissociated from pericardial cells. To characterize the physiological activity of SACs in the heart one would also want to perform voltage/patch clamp analysis while stretching the cardiac cells, perhaps while blocking SACs with the pharmacological effectors described in this study. It would then be interesting to learn if stretch activation is affected by the chemical modulators used in this study.
REFERENCES
- Shiels, H.A. and E. White, 2008. The frank-starling mechanism in vertebrate cardiac myocytes. J. Exp. Biol., 211: 2005-2013.
CrossRefDirect Link - Pathak, C.L., 1973. Autoregulation of chronotropic response of the heart through pacemaker stretch. Cardiology, 58: 45-64.
CrossRefDirect Link - Majeed, Z.R., A. Stacy and R.L. Cooper, 2014. Pharmacological and genetic identification of serotonin receptor subtypes on Drosophila larval heart and aorta. J. Comp. Physiol. B., 184: 205-219.
CrossRefPubMedDirect Link - Malloy, C.A., K. Ritter, J. Robinson, C. English and R.L. Cooper, 2015. Pharmacological identification of cholinergic receptor subtypes on Drosophila melanogaster larval heart. J. Comp. Physiol. B, 186: 45-57.
CrossRefPubMedDirect Link - Titlow, J.S., J.M. Rufer, K.E. King and R.L. Cooper, 2013. Pharmacological analysis of dopamine modulation in the Drosophila melanogaster larval heart. Physiol. Rep., Vol. 1, No. 2.
CrossRefDirect Link - Zhu, Y.C., E. Yocom, J. Sifers, H. Uradu and R.L. Cooper, 2016. Modulatory effects on Drosophila larva hearts: Room temperature, acute and chronic cold stress. J. Comp. Physiol. A., 186: 829-841.
CrossRefPubMedDirect Link - Sénatore, S., V.R. Reddy, M. Sémériva, L. Perrin and N. Lalevée, 2010. Response to mechanical stress is mediated by the TRPA channel painless in the Drosophila heart. PLoS Genet., Vol. 6, No. 9.
CrossRefDirect Link - Bodmer, R. and T.V. Venkatesh, 1998. Heart development in Drosophila and vertebrates: Conservation of molecular mechanisms. Dev. Genet., 22: 181-186.
PubMedDirect Link - Johnson, E., J. Ringo, N. Bray and H. Dowse, 1998. Genetic and pharmacological identification of ion channels central to the Drosophila cardiac pacemaker. J. Neurogenet., 12: 1-24.
CrossRefDirect Link - Olson, E.N., 2006. Gene regulatory networks in the evolution and development of the heart. Science, 313: 1922-1927.
CrossRefDirect Link - Choma, M.A., M.J. Suter, B.J. Vakoc, B.E. Bouma and G.J. Tearney, 2011. Physiological homology between Drosophila melanogaster and vertebrate cardiovascular systems. Dis. Models Mech., 4: 411-420.
CrossRefDirect Link - Dowse, H., J. Ringo, J. Power, K. Kinney and L. White, 1995. A congenital heart defect in Drosophila caused by an action-potential mutation. J. Neurogenet., 10: 153-168.
CrossRefDirect Link - Gu, G.G. and S. Singh, 1995. Pharmacological analysis of heartbeat in Drosophila. J. Neurobiol., 28: 269-280.
CrossRefDirect Link - De Castro, C., J. Titlow, Z.R. Majeed and R.L. Cooper, 2014. Analysis of various physiological salines for heart rate, CNS function and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A, 200: 83-92.
CrossRefDirect Link - Baumgartner, U., W. Greffrath and R.D. Treede, 2012. Contact heat and cold, mechanical, electrical and chemical stimuli to elicit small fiber-evoked potentials: Merits and limitations for basic science and clinical use. Clin. Neurophysiol., 42: 267-280.
CrossRefDirect Link - Coste, B., J. Mathur, M. Schmidt, T.J. Earley and S. Ranade et al., 2010. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science, 330: 55-60.
CrossRefDirect Link - Coste, B., B. Xiao, J.S. Santos, R. Syeda and J. Grandl et al., 2012. Piezo proteins are pore-forming subunits of mechanically activated channels. Nature, 483: 176-181.
CrossRefDirect Link - Ducret, T., J. El Arrouchi, A. Courtois, J.F. Quignard, R. Marthan and J.P. Savineau, 2010. Stretch-activated channels in pulmonary arterial smooth muscle cells from normoxic and chronically hypoxic rats. Cell Calcium, 48: 251-259.
CrossRefPubMedDirect Link - Hajduczok, G., M.W. Chapleau, R.J. Ferlic, H.Z. Mao and F.M. Abboud, 1994. Gadolinium inhibits mechanoelectrical transduction in rabbit carotid baroreceptors. Implication of stretch-activated channels. J. Clin. Investig., 94: 2392-2396.
CrossRefDirect Link - Omerbašić, D., L.N. Schuhmacher, Y.A.B. Sierra, E.S.J. Smith and G.R. Lewin, 2015. ASICs and mammalian mechanoreceptor function. Neuropharmacology, 94: 80-86.
CrossRefDirect Link - Rhodes, S.S., A.K. Camara, M. Aldakkak, J.S. Heisner and D.F. Stowe, 2015. Stretch‐induced increase in cardiac contractility is independent of myocyte Ca2+ while block of stretch channels by streptomycin improves contractility after ischemic stunning. Physiol. Rep., Vol. 3, No. 8.
CrossRefDirect Link - Welsh, M.J., M.P. Price and J. Xie, 2002. Biochemical basis of touch perception: Mechanosensory function of degenerin/epithelial Na+ channels. J. Biol. Chem., 277: 2369-2372.
CrossRefPubMedDirect Link - Delmas, P., J. Hao and L. Rodat-Despoix, 2011. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat. Rev. Neurosci., 12: 139-153.
CrossRefPubMedDirect Link - Cooper, A.S., K.E. Rymond, M.A. Ward, E.L. Bocook and R.L. Cooper, 2009. Monitoring heart function in larval Drosophila melanogaster for physiological studies. J. Vis. Exp., Vol. 33.
CrossRefDirect Link - Boscardin, E., O. Alijevic, E. Hummler, S. Frateschi and S. Kellenberger, 2016. The function and regulation of acid‐sensing ion channels (ASICs) and the epithelial Na+ channel (ENaC): IUPHAR review. Br. J. Pharmacol., 173: 2671-2701.
CrossRefDirect Link - Geffeney, S.L. and M.B. Goodman, 2012. How we feel: Ion channel partnerships that detect mechanical inputs and give rise to touch and pain perception. Neuron, 74: 609-619.
CrossRefDirect Link - Brand, A.H. and N. Perrimon, 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development, 118: 401-415.
PubMedDirect Link - Suslak, T.J., S. Watson, K.J. Thompson, F.C., Shenton, G.S. Bewick, J.D. Armstrong and A.P. Jarman, 2015. Piezo is essential for amiloride-sensitive stretch-activated mechanotransduction in larval Drosophila dorsal bipolar dendritic sensory neurons. PloS One, Vol. 10, No. 7.
CrossRefDirect Link - Yeung, E.W., N.P. Whitehead, T.M. Suchyna, P.A. Gottlieb, F. Sachs and D.G. Allen, 2005. Effects of stretch‐activated channel blockers on [Ca2+]i and muscle damage in the mdx mouse. J. Physiol., 562: 367-380.
CrossRefDirect Link - Bowman, C.L., P.A. Gottlieb, T.M. Suchyna, Y.K. Murphy and F. Sachs, 2007. Mechanosensitive ion channels and the peptide inhibitor GsMTx-4: History, properties, mechanisms and pharmacology. Toxicon, 49: 249-270.
CrossRefDirect Link - Sukharev, S. and F. Sachs, 2012. Molecular force transduction by ion channels-diversity and unifying principles. J. Cell Sci., 125: 3075-3083.
CrossRefDirect Link - Zornik, E., K. Paisley and R. Nichols, 1999. Neural transmitters and a peptide modulate Drosophila heart rate. Peptides, 20: 45-51.
CrossRefDirect Link - Gasque, G., S. Conway, J. Huang, Y. Rao and L.B. Vosshall, 2013. Small molecule drug screening in Drosophila identifies the 5HT2A receptor as a feeding modulation target. Scient. Rep., Vol. 3.
CrossRefDirect Link - Majeed, Z.R., C.D. Nichols and R.L. Cooper, 2013. 5-HT stimulation of heart rate in Drosophila does not act through cAMP as revealed by pharmacogenetics. J. Applied Physiol., 115: 1656-1665.
CrossRefPubMedDirect Link - Ren, G.R., J. Folke, F. Hauser, S. Li and C.J. Grimmelikhuijzen, 2015. The A-and B-type muscarinic acetylcholine receptors from Drosophila melanogaster couple to different second messenger pathways. Biochem. Biophys. Res. Commun., 462: 358-364.
CrossRefDirect Link - Zhu, Y.C., H. Uradu, Z.R. Majeed and R.L. Cooper, 2016. Optogenetic stimulation of Drosophila heart rate at different temperatures and Ca2+ concentrations. Physiol. Rep., Vol. 4, No. 3.
CrossRefDirect Link - Sokabe, T., S. Tsujiuchi, T. Kadowaki and M. Tominaga, 2008. Drosophila painless is a Ca2+-requiring channel activated by noxious heat. J. Neurosci., 28: 9929-9938.
CrossRefDirect Link - Desai-Shah, M., A.R. Papoy, M. Ward and R.L. Cooper, 2010. Roles of the sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, plasma membrane Ca2+-ATPase and Na+/Ca2+ exchanger in regulation of heart rate in Larval drosophila. Open Physiol. J., 3: 16-36.
CrossRefDirect Link - Lacampagne, A., F. Gannier, J. Argibay, D. Garnier and J.Y. Le Guennec, 1994. The stretch-activated ion channel blocker gadolinium also blocks L-type calcium channels in isolated ventricular myocytes of the guinea-pig. Biochim. Biophys. Acta (BBA)-Biomembr., 1191: 205-208.
CrossRefDirect Link - Bae, C., F. Sachs and P.A. Gottlieb, 2011. The mechanosensitive ion channel Piezo1 is inhibited by the peptide GsMTx4. Biochemistry, 50: 6295-6300.
CrossRefDirect Link - Whitehead, N.P., M. Streamer, L.I. Lusambili, F. Sachs and D.G. Allen, 2006. Streptomycin reduces stretch-induced membrane permeability in muscles from mdx mice. Neuromuscular Disord., 16: 845-854.
CrossRefDirect Link - Belus, A. and E. White, 2002. Effects of streptomycin sulphate on ICaL, IKr and IKs in guinea-pig ventricular myocytes. Eur. J. Pharmacol., 445: 171-178.
CrossRefDirect Link - Redaelli, E., R.R. Cassulini, D.F. Silva, H. Clement and E. Schiavon et al., 2010. Target promiscuity and heterogeneous effects of tarantula venom peptides affecting Na+ and K+ ion channels. J. Biol. Chem., 285: 4130-4142.
CrossRefDirect Link - Kayukawa, T. and Y. Ishikawa, 2009. Chaperonin contributes to cold hardiness of the onion maggot Delia antique through repression of depolymerization of actin at low temperatures. PLoS One, Vol. 4, No. 12.
CrossRefDirect Link