
Abdul Sadat
Entomology Laboratory, Department of Zoology, University of Gour Banga, Malda, 732103, West Bengal, India
LiveDNA: 91.16420
Shubham Bhattacharyya
Immunopharmacology and Molecular Cell Biology Laboratory, Department of Zoology, University of Gour Banga, 732103 Malda, West Bengal, India
Subhrajyoti Roy
Immunopharmacology and Molecular Cell Biology Laboratory, Department of Zoology, University of Gour Banga, 732103 Malda, West Bengal, India
Kaushik Chakraborty
Entomology Laboratory, Department of Zoology, University of Gour Banga, Malda, 732103, West Bengal, India
Journal of Entomology
Year: 2019 | Volume: 16 | Issue: 1 | Page No.: 23-29







ABSTRACT
Background and Objective: Corchorus olitorius, an important fiber crop in Indian economy, is infested by many pests causes it to evolve protective defense against them. In present era, Anomis sabulifera is a major threat to jute agronomy. In this study, the morphological and phyto-chemical defense strategy of jute against A. sabulifera was determined. Materials and Methods: Jute leaf phenology was studied along with standard biochemical and spectrophotometric procedures for the quantification of phyto-chemicals like saponin, alkaloid, flavonoid, tannin, riboflavin, ascorbic acid, thiamine, soluble sugar, protein, lipid, total moisture and ash content. The results were correlated with the pest incidence. Results: Phenological characters were improved throughout the jute cultivation period whereas leaf bio-chemistry showed an altered pattern. After pest menace there was depletion in leaf nutrient content although secondary metabolite concentration was found to be increased following pest attack. Conclusion: It was concluded that, C. olitorius have no morphological defense strategy against A. sabulifera whereas it has evolved itself via phyto-chemical defense mechanisms to protect from pest attack. These findings may be helpful to plan better cultural strategy and pest control programme in near future.
PDF Abstract XML References Citation
Received: November 11, 2018;
Accepted: December 28, 2018;
Published: February 22, 2019
Copyright: © 2019. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Abdul Sadat, Shubham Bhattacharyya, Subhrajyoti Roy and Kaushik Chakraborty, 2019. Defensive Response of Corchorus olitorius in Relation to Infestation of Anomis sabulifera. Journal of Entomology, 16: 23-29.
DOI: 10.3923/je.2019.23.29
URL: https://scialert.net/abstract/?doi=je.2019.23.29
DOI: 10.3923/je.2019.23.29
URL: https://scialert.net/abstract/?doi=je.2019.23.29
INTRODUCTION
The success of plants in colonizing and dominating almost every habitat on earth depends on their ability of co-evolution to persist in hostile and erratic conditions of earth1,2. This was largely due to the presence of effective resistance mechanism which includes physical, chemical and developmental features3. This diversity in plants defense mechanism was evolved through their long lasting struggle for co-existence with the herbivorous animals4. Throughout this co-existence host and pest interact in broad spectrum that influences organisms at every level including both basic biochemical events and phenological constituent5. Study of host-pest dealings revealed that success of phytophagous insects was resulted from effective neutralization of hosts defense mechanism1,3. Some of these interactions are reciprocally beneficial, such as pollination but most of the interactions involved insect dependence on plants for food and equivocally plants defend themselves from herbivorous insects6. In fact, the pray-predator association is extensive as because almost every plant species is eaten by at least one insect species7-9. Therefore, plants not only have adapted to their morphological resistance properties but a variety of secondary metabolites have also been synthesized in plants due to the selective pressure exerted by phytophagous animals10,11. Anomis sabulifera (jute semilooper) is a major leaf feeding pest of jute plant that causes severe damage to the host plant4. During severe infestation A. sabulifera feed up to 90% jute leaf that ultimately leads to 50% fibre yield loss5. To fulfill the market demand and use of more and more fertilizers and insecticides, jute pests becomes resistant to stress and become major threat for production yield12. Jute plants are devoid of physical resistance like epicuticular wax, resin or thorns. Therefore, jute leaves are easily accessible food for semilooper larvae but heterogeneous infestation pattern was recorded during jute cultivation period13,14. Utilization of plant defense mechanism is a key component for IPM programme, which looks for minimizing pest damage through a combination of cultural and biological control (natural enemies and endogenous resistance of host plant) with minimal use of chemical methods15,16. The present study was designed to evaluate the morphological and phyto-chemical defense response of jute plant, Corchorus olitorius in relation to prevalence of jute semilooper, Anomis sabulifera which will explore new facet of host plant resistance system for improvement of pest control programme under IPM model.
MATERIALS AND METHODS
Experimental design: The experiment was conducted at 3 administrative blocks of west Bengal namely Raiganj, Hemtabad and Kaliyaganj in 2016 covering the calendar month of April to August in randomized block design with 3 replications adopting standard cultivation practices. Before sowing N, P2O5 and K2O at the rate of 20 kg ha–1 were applied as basal application. The jute seeds were sown in a row spacing of 25 cm in small plots of 4×4 m with a gap of 1 m between each plot. At completely grown condition, plant to plant distance was maintained at 6-8 cm apart after thinning. Samples were collected at 4 different growth stages of jute plant at 30, 60, 90 and 120 DAS (Day after Sowing) for further examination.
Estimation of plant characteristics: Plant height, leaf area, leaf roundness, specific leaf area, leaf thickness, laminar length, laminar width, leaf ratio, leaf perimeter and leaf fresh weight were measured by standard method with necessary modification17,18.
Estimation of leaf phyto-chemistry
Plant material and sample preparation: Jute leaves were collected from 3 study sites of Uttar Dinajpur district, west Bengal, India. The leaves were first washed in tap water and then in double distilled water to remove dirt. The leaves were then dried at room temperature for 7 days, finely powdered and used for quantitative analysis of different phyto-chemicals.
Quantitative assay of phyto-chemicals: The quantitative assays of different phyto-chemical components such as total protein19, total lipid20, total sugar21, alkaloid22, flavonoid23, saponin24, riboflavin25, thiamin26, ascorbic acid27, phenol22, moisture and ash content of the collected jute leaves were determined according to standard protocols.
Observation on pest incidence: Three replicated plots (1 m2) in each of the pre-selected sites were observed for incidence of jute semilooper, Anomis Sabulifera throughout the jute growing season in the year 2016. Percentage of plant infested with A. Sabulifera larvae was estimated by observing top 8 damaged leaves of each plant18.
Statistical analysis: All the experiments were performed 3 times and data were represented as Mean±SD for further statistical analysis. Data were analyzed for correlation coefficient (r) with 5% significance level by using statistical tool kyplot version 2.0 beta 15 (32 bit)28.
RESULTS
Study on the phenological and phyto-chemical defense of jute plant against phytophagous semilooper insects were done in Uttar Dinajpur district of west Bengal during pre-kharif to kharif season of the year 2016.
Observation on morphological characteristics of jute leaf: Morphological data in Table 1 of the present study showed that nutrition contributing factors as, plant height, leaf area, laminar length and width, leaf perimeter and fresh leaf weight were increased rapidly with time. Whereas, limited changes were observed in leaf thickness (118.23-174.25, 118.56-174.66 and 118.35-174.80 μm), specific leaf area (0.033-0.033, 0.032-0.036 and 0.029-0.036), leaf roundness (40.67-40.00, 39.30-42.00 and 39.50-41.000) and leaf ratio (2.74-2.95, 2.67-3.25 and 2.71-3.13 ) at 3 study sites viz., Raiganj, Hemtabad and Kaliyaganj, respectively.
Observation on phyto-chemical characteristics of jute leaf: Quantitative study in Table 2 of jute leaf extract revealed that concentration of leaf nutrients like total sugar, total protein and total lipid rapidly increased from 30-90 DAS.
| Table 1: | Morphological characteristics of jute leaf (C. olitorius) at different growth stages during 2016 |
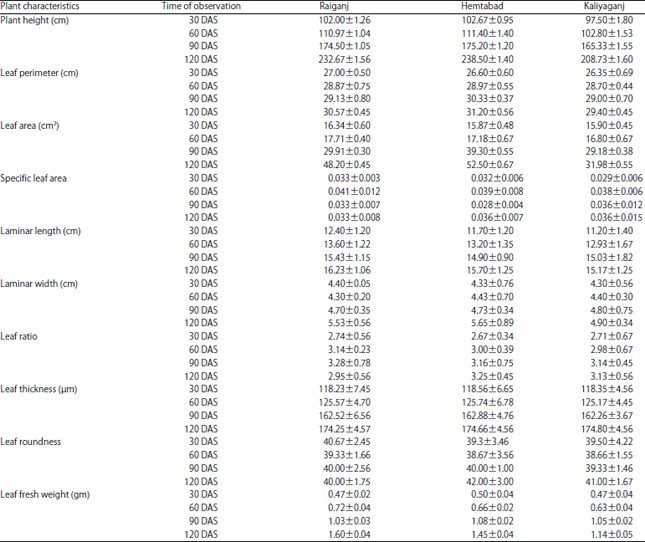 | |
| Data are presented as Mean±SD | |
| Table 2: | Phyto-chemical quantification of jute leaf (C. olitorius) at different growth stages during 2016 |
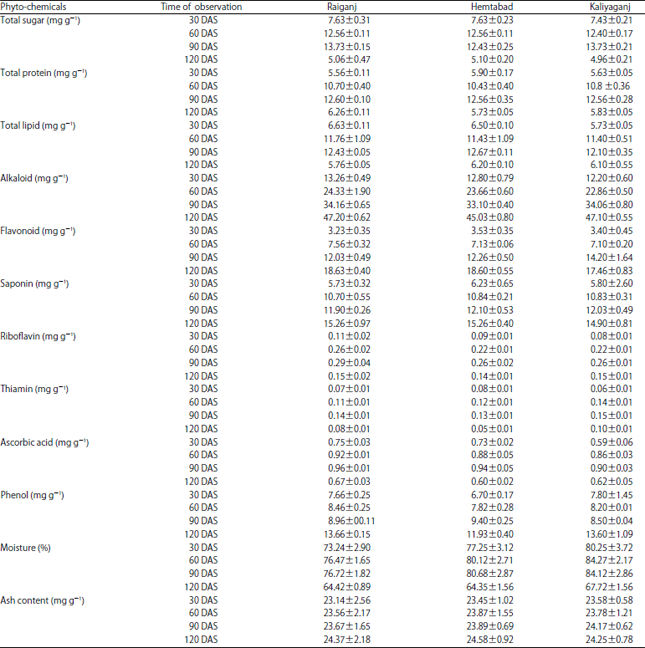 | |
| Data are presented as Mean±SD | |
But at late age period (120 DAS) of jute crop, concentration of those leaf nutrients decreased rapidly. From 30-90 DAS growth phase there was lower concentration of secondary metabolites. Whereas, highest concentration of secondary metabolites was recorded at late growth phase (120 DAS). Although, phyto-chemical estimation showed that leaf moisture and ash content was increased throughout the crop development.
Inter-relationship within leaf phenology and phyto-chemistry: In present study, matrix analysis revealed that major attributing morphological characters in Table 3 were imparted significantly (p<0.05) positive effect on each other, except specific leaf area. Table 4 showed a different pattern of inter-dependence between leaf chemical contents. Primary metabolites such as leaf carbohydrates, proteins, lipids, ascorbic acid, riboflavin and thiamin which were main nutrient contributing factors showed significantly (p<0.05) positive interaction with each other.
| Table 3: | Matrix analysis for inter-relationship of jute leaf phenology |
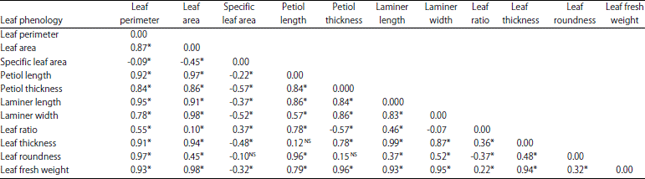 | |
| Significant at *p<0.05 | |
| Table 4: | Matrix analysis for inter-relationship study of jute phyto-chemicals |
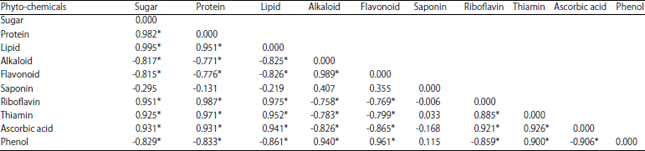 | |
| Significant at *p<0.05 | |
But interestingly, leaf nutritional characters and secondary metabolites imparted significant (p<0.05) negative effects between them. Out of the all secondary metabolites phenol exerted highest negative influence on leaf nutrients.
Correlation of pest (A. sabulifera) incidence with leaf morphology and phyto-chemistry of jute: Correlation studies Table 5 and 6 showed that relationship of incidence of A. sabulifera with the leaf morphology and phyto-chemistry of jute crop that were tested for understanding plants defense against pest herbivory in 2016. The results revealed that among various morphological characteristics, plant height, laminar length, laminar width, leaf perimeter, leaf area, leaf thickness and leaf roundness showed significant positive influence of pest menace. Whereas, specific leaf area, leaf ratio and leaf fresh weight imparted a negative impact on pest incidence.
Correlation analysis also revealed that pest menace had a significant negative impact on leaf nutrient such as sugar (p<0.05), protein (p<0.01), lipid (p<0.01), riboflavin (p<0.001), thiamine (p<0.001) and ascorbic acid (p<0.001). However, pest attack leads to increase in alkaloid (p<0.01), flavonoid (p<0.05), saponin (p<0.05), phenol (p<0.01), moisture (p<0.001) and ash content (p<0.001).
| Table 5: | Correlation of A. sabulifera with jute leaf morphology during 2016 |
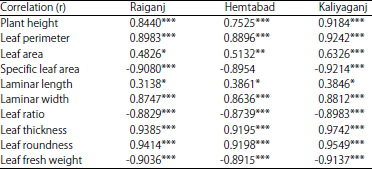 | |
| Significant at *p<0.05, **<0.01, ***<0.001, NS: Not significant | |
| Table 6: | Correlation of A. sabulifera with phytochemistry of jute leaf during 2016 |
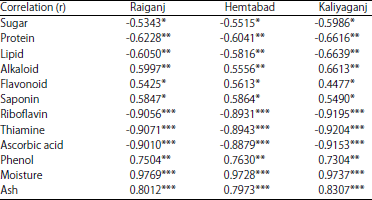 | |
| Significant at *p<0.05, **<0.01, ***<0.001, NS: Not significant | |
DISCUSSION
The crop morphology mainly the yield attributing characters such as crop height, leaf length, width, perimeter, area and thickness were found increasing towards ripeness of the crop, whereas, specific leaf area, roundness and leaf ratio did not show incremental trend towards maturity stages of plant during study period of 2016. Incidence of jute semilooper showed a positive influence on yield attributing characters of the jute crop. Increase in plant vigour after pest attacks was to compensate the damaged caused by pest herbivory29,30 as jute plants don’t have any strapping physical barrier against its pest. Increased leaf thickness made them more harden to eat by phytophagous pest which was first line of defence against herbivorous pest14. These findings were of similar with the result of the study made by Rahman and Khan18. However, plant can’t defend them only with this morphological defence line and there must be collaboration between physical and chemical14 defence machineries.
Phyto-chemicals played important role in nutritional ecology and growth of a phyto-phagous insect3. Host-plant selection of herbivorous pest was influenced by their capability to ingest and assimilate food matter to transform it into body tissues31. According to Roy32 the development and survival of insects showed significant differences with respect to their food quality. Primary metabolites such as carbohydrate, protein and lipid were key factor for general vitality, growth and reproduction of phytophagous insects33.
Further, during diapauses lipids provide energy for metabolic processes and also act as precursors of ecdysteroid34. Water content in host leaves played an important role in growth rate of plant-fed larvae18. On the other hand, secondary metabolites including phenols, flavonoids, terpenoids, alkaloids, etc., concluded the fitness of the food utilization of the phytophagous pest and thus govern plant-insect interaction14. Consumption of greater amount of secondary metabolites was found to significantly reduce adult longevity, fecundity and retardation of larval growth35. Present study revealed that young and mature leaves (30-90 DAS) of jute plant provide the best quality food for the major defoliator, A. sabulifera, because of higher nutritional factors relative to the anti-nutritional secondary metabolites that resulted into high pest incidence. Whereas, secondary metabolites was dominating over leaf nutrients in late growth phase (120 DAS), that limits pest menace. The better understanding of this defence system will allow us to reach more useful methods for the biological control of insect pests with natural products by the development of new jute varieties with enhanced chemical defences.
CONCLUSION
It was concluded that jute crop had not developed any distinct physical barrier in the form of epicuticular wax or resin or thorns. Instead of that, plant vigour attracted its pest initially. On the other hand, pest attack induce host plants chemical defence by elevating different secondary metabolites in the form of phenols, alkaloids, flavonoids and saponin which act as feeding deterrent to eliminate pest population. The stipulated information provides a well decorated knowledge about host-pest interaction and defence strategies of jute crop against A. sabulifera that will helpful to generate IPM model for sustainable production of jute crop.
SIGNIFICANCE STATEMENT
Present study revealed the influence of A. sabulifera on the defensive strategies of C. olitorius which are a major threat to jute cultivation. This study will help the researcher to explore the vital areas of host-pest interaction for jute crop which can be beneficial for construction of improved crop culture practice and pest control.
ACKNOWLEDGMENT
We acknowledge University of Gour Banga, Malda, India for providing infrastructure to run the research work.
REFERENCES
- Smith, M.A.K., 2000. Comparative response of Chromolaena odorata and Corchorus olitorius to intraspecific competition. Agric. Sci. Digest, 20: 141-145.
Direct Link - Shobana, K., K. Murugan and A.N. Kumar, 2010. Influence of host plants on feeding, growth and reproduction of Papilio polytes (The common mormon). J. Insect Physiol., 56: 1065-1070.
CrossRefDirect Link - Agrawal, A.A., 2004. Resistance and susceptibility of milkweed: Competition, root herbivory and plant genetic variation. Ecology, 85: 2118-2133.
CrossRefDirect Link - Bernays, E.A. and R.F. Chapman, 1977. Deterrent chemicals as a basis of oligophagy in Locusta migratoria (L.). Ecol. Entomol., 2: 1-18.
CrossRefDirect Link - Kessler, A. and I.T. Baldwin, 2002. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol., 53: 299-328.
CrossRefDirect Link - Chen, H., A.D. Jones and G.A. Howe, 2006. Constitutive activation of the jasmonate signaling pathway enhances the production of secondary metabolites in tomato. FEBS Lett., 580: 2540-2546.
CrossRefDirect Link - Rahman, S. and M.R. Khan, 2012. Incidence of pests in jute (Corchorus olitorius L.) Ecosystem and pest-weather relationships in West Bengal, India. Arch. Phytopathol. Plant Prot., 45: 591-607.
CrossRefDirect Link - Rahman, S. and M.R. Khan, 2010. Integrated management approach for control of the pest complex of olitorius jute, Corchorus olitorius L. J. Plant Prot. Res., 50: 340-346.
CrossRefDirect Link - Ranger, C.M., R.E. Winter, A.P. Singh, M.E. Reding, J.M. Frantz, J.C. Locke and C.R. Krause, 2011. Rare excitatory amino acid from flowers of zonal geranium responsible for paralyzing the Japanese beetle. Proc. Natl. Acad. Sci. USA., 108: 1217-1221.
CrossRefDirect Link - Savci, S., 2012. Investigation of effect of chemical fertilizers on environment. APCBEE Procedia, 1: 287-292.
CrossRefDirect Link - Agrawal, A.A., 2011. Current trends in the evolutionary ecology of plant defence. Funct. Ecol., 25: 420-432.
CrossRefDirect Link - Sadat, A. and K. Chakraborty, 2015. Insect pest constrains of jute and its control by biological agents under modern eco-friendly sustainable production system. Int. J. Multidiscip. Res. Dev., 2: 316-321.
Direct Link - Leiss, K.A., Y.H. Choi, R. Verpoorte and P.G. Klinkhamer, 2011. An overview of NMR-based metabolomics to identify secondary plant compounds involved in host plant resistance. Phytochem. Rev., 10: 205-216.
CrossRefDirect Link - Kumar, A. and P.M. Nigam, 2010. Economic and Applied Entomology. Reprint Edn., Emkay Publications, Delhi, India, ISBN-13: 9788185712550, Pages: 349.
Direct Link - Rahman, S. and M.R. Khan, 2012. Effect of plant characteristics of jute varieties on incidence of pests in West Bengal, India. Arch. Phytopathol. Plant Prot., 45: 606-619.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Jayaram, J., 2011. Laboratory Manual in Biochemistry. New Age International Pvt. Ltd. Publishers, India, ISBN-13: 978-8122430493.
Direct Link - Dubois, M., K. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1951. A colorimetric method for the determination of sugars. Nature, 168: 167-167.
CrossRefPubMedDirect Link - Obadoni, B.O. and P.O. Ochuko, 2002. Phytochemical studies and comparative efficacy of the crude extracts of some haemostatic plants in Edo and Delta States of Nigeria. Global J. Pure Appl. Sci., 8: 203-208.
CrossRefDirect Link - Bohm, B.A. and M.R. Koupai-Abyazani, 1994. Flavonoids and condensed tannins from leaves of Hawaiian Vaccinium reticulatum and V. calycinum (Ericaceae). Pac. Sci., 48: 458-463.
Direct Link - Nahapetian, A. and A. Bassiri, 1975. Changes in concentrations and interrelations of phytate, phosphorus, magnesium, calcium and zinc in wheat during maturation. J. Agric. Food Chem., 23: 1179-1182.
CrossRefPubMedDirect Link - Abe, F. and T. Yamauchi, 1992. Cardenolide triosides of oleander leaves. Phytochemistry, 31: 2459-2463.
CrossRefDirect Link - Poornima, G.N. and R.V. Ravishankar, 2009. Evaluation of phytonutrients and vitamin contents in a wild yam, Dioscorea belophylla (Prain) Haines. Afr. J. Biotechnol., 8: 971-973.
Direct Link - Sadat, A. and K. Chakraborty, 2017. Studies on incidence of jute semilooper in relation to weather of Uttar Dinajpur, India. J. Entomol., 14: 96-103.
CrossRefDirect Link - Choong, M.F., P.W. Lucas, J.S.Y. Ong, B. Pereira, H.T.W. Tan and I.M. Turner, 1992. Leaf fracture toughness and sclerophylly: Their correlations and ecological implications. New Phytol., 121: 597-610.
CrossRefDirect Link - Agrawal, A.A. and K. Konno, 2009. Latex: A model for understanding mechanisms, ecology and evolution of plant defense against herbivory. Annu. Rev. Ecol. Evol. Syst., 40: 311-331.
CrossRefDirect Link - Nation, Sr. J.L., 2015. Insect Physiology and Biochemistry. 3rd Edn., CRC Press, Boca Raton, FL., USA., ISBN-13: 9781482247589, Pages: 690.
Direct Link - Roy, N., 2014. Role of Chorchorus capsularis phytochemicals on the feeding dynamics of Diacrisia casignetum Kollar (Lepidoptera: Arctiidae). J. Entomol. Zool. Stud., 2: 227-236.
Direct Link - Turunen, S., 1990. Plant leaf lipids as fatty acid sources in two species of Lepidoptera. J. Insect Physiol., 36: 665-672.
CrossRefDirect Link - Genc, H. and J.L. Nation, 2004. Influence of dietary lipids on survival of Phyciodes phaon butterflies (Lepidoptera: Nymphalidae). J. Entomol. Sci., 39: 537-544.
CrossRefDirect Link - Mazid, M., T.A. Khan and F. Mohammad, 2011. Role of secondary metabolites in defense mechanisms of plants. Biol. Med., 3: 232-249.
Direct Link