
Fri Maulina
Agricultural Polytechnic of Payakumbuh/Study Program of Agricultural Sciences, Post Graduate Program, Andalas University Pascasarjana Building, Limau Manis, 25163 Padang, Indonesia
LiveDNA: 62.21815
Novri Nelly
Department of Plant Protection, Faculty of Agriculture, Andalas University, Limau Manis, 25163 Padang, Indonesia
Hidrayani
Department of Plant Protection, Faculty of Agriculture, Andalas University, Limau Manis, 25163 Padang, Indonesia
Hasmiandy Hamid
Department of Plant Protection, Faculty of Agriculture, Andalas University, Limau Manis, 25163 Padang, Indonesia
Journal of Entomology
Year: 2018 | Volume: 15 | Issue: 1 | Page No.: 13-18







ABSTRACT
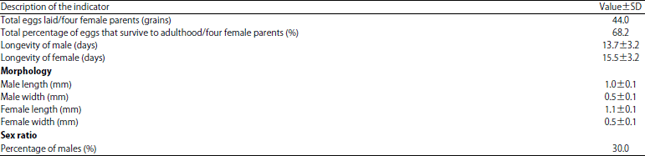
Background and Objective: Temperature and survival of a parasitoid egg before it reaches the adult stage are some of the most important biological parameters that must be understood to effectively and efficiently perform parasitoid mass rearing in a laboratory. The objectives of this study were to determine the best maintenance tube size for the longevity of the adult parasitoid, Hadronotus leptocorisae (H. leptocorisae), to determine the survival rate of the eggs laid by this parasitoid and to determine the fitness and sex ratio of the offspring. Materials and Methods: The eggs of Leptocorisa oratorius (L. oratorius) were collected from paddy fields in Sungai Sapih, Padang city and the experiments were carried out in a laboratory at 25°C and at room temperature. Data on the temperature differences were analyzed by a t-test and the various tube sizes were analyzed by a completely randomized design with STATISTIC 8.0 programs. A descriptive method was used to present fitness data. Results: The ideal maintenance temperature to ensure longevity of the parasitoid Hadronotus leptocorisae is 25°C. The tube size used at that temperature is not significantly affected. The mean survival of eggs laid by a female H. leptocorisae until adulthood was 68.2%. The mean longevity of males and females was 13.7±3.2 and 15.5±3.2 days, respectively. The mean male parasitoid body size was 1.0±0.1 mm long and 0.5±0.1 mm wide and the average female parasitoid size was 1.1±0.1 mm long and 0.5±0.1 mm wide. The percentage of parasitoid males was 30%. Conclusion: The best condition for rearing with a high survival rate is 25°C with no specific tube size.
PDF Abstract XML References Citation
Received: November 10, 2017;
Accepted: February 16, 2018;
Published: March 28, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Fri Maulina, Novri Nelly, Hidrayani and Hasmiandy Hamid, 2018. Temperature as a Key Aspect in the Survival of Hadronotus leptocorisae Offspring. Journal of Entomology, 15: 13-18.
DOI: 10.3923/je.2018.13.18
URL: https://scialert.net/abstract/?doi=je.2018.13.18
DOI: 10.3923/je.2018.13.18
URL: https://scialert.net/abstract/?doi=je.2018.13.18
INTRODUCTION
The Hadronotus leptocorisae Nixon (Gryon nixoni Masner) parasitoid have been reported to be in the eggs of the rice bug (Leptocorisa oratorius Fabricius) at Lombok Island, West Nusa Tenggara Province1 and at Padang Pariaman2 and Tanah Datar3 Regency, West Sumatra Province, Indonesia. The existence of parasitoid required biological studies to determine effective and efficient biological control agents4,5. One such biological study was the mass rearing of parasitoids in the laboratory to obtain offspring5 parasitoids that had high fitness and were ready to be introduced into the field.
The survival of eggs laid by a parasitoid on the host before it emerges as an adult is essential to the continuity of its use as a biological control agent. Insect survival responses (including parasitoids) are affected by temperature6,7, host susceptibility (i.e., host stage)6,8 and parasitoid virulence8. Temperature determines the superparasitism of a parasitoid9, which affects its survival. Parasitoid survival does not vary linearly with temperature, because survival depends on the type of parasitoid. High mortality usually occurs at lower and upper threshold temperatures6, so it is important to understand which temperature is suitable to the development of parasitoids. Optimum temperatures are required for parasitoids to adequately complete their life cycle. The ideal temperature to ensure the longevity of the H. leptocorisae adult parasitoid as an egg parasitoid of Leptocorisa oratorius is 25°C for a 22.6 days life cycle10. The effects of maintenance tube size on the longevity of H. leptocorisae parasitoids at room temperature and at 25°C were done in this study. In addition, the survival of the H. leptocorisae eggs, the fitness and sex ratio of H. leptocorisae offspring at 25oC were found. Therefore, this information can be used as a reference in mass rearing of H. leptocorisae in the laboratory.
MATERIALS AND METHODS
Longevity of the Hadronotus leptocorisae parasitoid at different temperatures and in different tubes: Samples (the eggs of L. oratorius) were collected from paddy fields of Sungai Sapih, Padang City (20 m above the sea level) using a transect line method (1 km). The land selection criteria were that the area had to be at least 3 ha and the condition of paddy plants had to be at a milk mature stage. The collected eggs were observed at the Insect Bioecology Laboratory, Plant Protection Department of Andalas University (220 m above the sea level). This activity was conducted from September, 2016 until January, 2017. Imago of H. leptocorisae that emerged from the egg were placed in test tubes of different sizes (diameter:height) (cm:cm) as follows: A (1:7), B (1:9.5), C (2:10), D (1.5:12.5), E (2:14), F (2:17) and G (2.5:20). A drop of honey was placed on the inside of the test tube and the tubes were placed under two conditions, at 25°C and at room temperature. Observations were conducted every day until the imago parasitoid was dead.
Survival of the Hadronotus leptocorisae parasitoid at optimum temperatures: The most suitable temperature for the development of the H. leptocorisae parasitoid (the result of step one), i.e., 25°C, was used for survival testing of parasitoid offspring. The adult female sample of the H. leptocorisae parasitoid that was used as the survival test included 4 parasitoids. The 4 female parasitoids were inserted into a separate enclosed test tube in which there was already a male parasitoid. Then, the male and female parasitoids were allowed to copulate for 24 h. The adult female was transferred into a separate test tube where they were placed on pias paper that had 10 eggs from the host for oviposition. After 24 h, the eggs were replaced with new eggs (from the insect cage). This activity was done every day until the female parasitoid was dead. Observations of the parasitoid offspring included those related to fitness (i.e., longevity (observations are made daily), morphology (observations were performed under the binocular stereo microscope), Carton SPz 50 (Aven Tools, USA) and photographed with the Olympus SZ 14 camera that was inserted a millimeter with a snipping tool) and sex ratio.
Statistical analysis: Data on the temperature differences were analyzed by a t-test and the variance of tubes was analyzed by a completely randomized design. A correlation analysis between the various parameters was conducted using linear regression. The statistical analysis were conducted in STATISTICS 8.0 (USDA, NRCS, USA). The fitness data were reported using descriptive statistics as the means±standard deviation and the sex ratio was calculated by the percentage of live males as follows:
| (1) |
Where:
| PM | = | Percentage of live males |
| M | = | Amount of emerging males |
| F | = | Amount of emerging female |
RESULTS
Longevity of the Hadronotus leptocorisae parasitoid at different temperatures and in different tubes: The longevity of the H. leptocorisae parasitoid that emerged from the eggs of the rice bug (L. oratorius) that was maintained under different temperature conditions was significantly different (p = 0.003) between room temperature and 25°C with a mean longevity of 9.7 and 19.1 days, respectively. The various tube size treatments showed that the longevities of the adult H. leptocorisae parasitoid were not different at 25°C, whereas, at room temperature, there was a difference in the longevity between treatments A and C with E, F and G (Table 1).
Survival of the Hadronotus leptocorisae parasitoid at the optimum temperature: Testing four H. leptocorisae parasitoid females at the optimum temperature (25°C) showed that the eggs that were laid (4-28 eggs) were highly varied with a mean of 11±11.2 grains (Fig. 1). The percentage of eggs that had been able to survive until the adult stage was 52%. Overall, of the 44 eggs from the 4 female parental parasitoids, 78% survived until the adult stage (Table 2).
The tests that were performed showed that the proportion of surviving parasitoid female imago (lx) declined sharply 12 days after the imago was treated (Fig. 2), whereas, the eggs that were laid and survived as imago fluctuated from day 2-9. Overall, the potential age for a female imago to lay its eggs is from the 2nd to the 9th day, however, when it is above the age of 12 days, there is a decrease in its reproductive rate (lx×mx).
Fitness of Hadronotus leptocorisae offspring parasitoids: The growth/development of the eggs laid on the host (L. oratorius egg) were quite high at 25°C. More than half of the eggs (68.2%) (Table 2) survived until adult-stage parasitoids. The high survival of the eggs illustrates that the environmental conditions were suitable for mass rearing of the parasitoid in the laboratory.
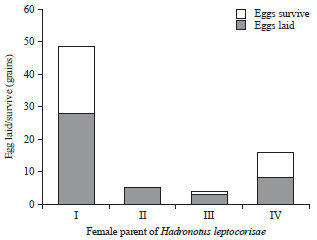 | |
| Fig. 1: | Ability of the female to lay eggs and the survival of the Hadronotus leptocorisae parasitoid at optimum temperature conditions |
| Table 1. | Longevity of Hadronotus leptocorisae at room temperature and 25°C with a variety of tubes |
 | |
The number in the right columns followed by the same notation are not significantly different by LSD test 5% level (CV = 1.97) | |
| Table 2: | Fitness of Hadronotus leptocorisae parasitoid offspring from four parasitoid female parents that survive until the adult stage at the optimum temperature (25°C) |
 | |
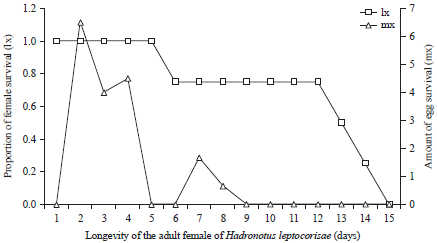 | |
| Fig. 2: | Survivorship of the female Hadronotus leptocorisae parasitoid |
The longevity of male and female parasitoid offspring under laboratory conditions at 25°C was 13.7±3.2 and 15.5±3.2 days, respectively. The body size of the female parasitoid was larger than that of the male in terms of length and width, which were 1.1±0.1 and 0.5±0.1 mm and 1.0±0.1 and 0.5±0.1 mm, respectively. The sex ratio between female and male parasitoids of H. leptocorisae was 30:70, meaning 30% of the parasitoids that emerged were male and 70% were females.
DISCUSSION
In this study, conditions at 25°C (the optimum temperature based on our previous studies) were compared with the standard condition, i.e., room temperature. Samples were placed in test tubes of various sizes to understand the effect of maintenance space on longevity. The obtained results demonstrated that 25°C provides the highest longevity. The use of different test tube sizes did not have an effect when the sample was placed in a 25°C room condition. Under these conditions, the temperature in the tube is the same as the room temperature at 25°C. The constant temperature was more suited to the metabolism of the adult parasitoid H. leptocorisae, thus resulting in longer longevity than when the samples were at room temperature. Similarly, the placement of the samples at room temperature provided the same conditions as the maintenance tube so that changing the temperature makes a difference in the length of the longevity of the H. leptocorisae adult parasitoid. There are differences in test tubes A and C with E, F and G in terms of longevity of the H. leptocorisae adult parasitoid. This result was reinforced by the high correlation between the various tube sizes and the length of adult longevity of H. leptocorisae, i.e., R = 0.89 or at 25°C, the lower correlation is R = 0.55 (Fig. 3). Adult longevity will contribute to the number of eggs that are exposed affecting the sustainability of their existence in the field.
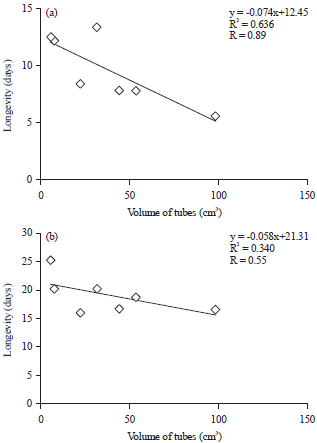 | |
| Fig. 3(a-b): | Correlation between the volumes of the rearing tube with the longevity of the Hadronotus leptocorisae parasitoid under two conditions (a) Room temperature and (b) 25°C |
The survival ability of H. leptocorisae eggs until they emerge as adults is greater than 50% for both the mean count or with four female parental parasitoids (i.e., 52 and 78%, respectively), illustrating that the parasitoid can be developed in the laboratory. The temperature of 25°C as the test temperature was based on previous research10 that the imago parasitoid H. leptocorisae survived the longest at 22.6 days, thus, it is expected to have a duration of oviposition that will be longer throughout its life. This condition was very important in the empowerment of natural competitors as a biological control agent of the pest. Their survival can be determined not only by temperature11 but also by nutrition in both artificial agroecosystems11 and the field12,13. Determining the optimum laboratory conditions should be carried out on host that a uniformity. In the present study, the high variation in the number of eggs is likely related to the different qualities of eggs on the host. Eggs were used from a host maintenance sample with relatively uniform ages and sizes, but when egg dissection occurred, the eggs were empty. No adult pest of L. oratorius was found on (if not parasitized), or the adult parasitoid H. leptocorisae was found (if eggs are parasitized) (unpublished data). This condition causes the eggs that were parasitized by the parasitoid females to not be able to develop.
The female H. leptocorisae parasitoid offspring had a longer longevity than males (13.7±3.2 and 15.5±3.2 days, respectively), which is a common phenomenon in insects. Life expectancy in females is almost always greater than in males. When compared with parasitoid sample data collected from the field, the difference in the mean age of parasitoids between males and females was relatively longer at 22.6±5.4 days10. The width and length of the H. leptocorisae female parasitoid are also greater than that of males (1.1±0.1 and 0.5±0.1 mm and 1.0±0.1 and 0.5±0.1 mm, respectively). Body size is related to the nutrients that are in the host (body fluid host). The larger the host, more food reserves are available for the parasitoids in the host (endoparasitoid). The H. leptocorisae female parasitoid has can detect the host before laying its eggs, by using an antenna or ovipositor as a detector tool. According to Anggraeni et al.14, the behavior of H. leptocorisae, laying eggs begins with an examination of the host and is followed by oviposition. The presence of more females than males (70:30) is an advantage because the females are going to spawn on the rice bug eggs. Females who continue will be a biological control agent in the field. There are differences in the sex ratio of H. leptocorisae in the field (unpublished data), i.e., there are more males than females. This result suggests that under no choice conditions, the female parasitoid tends to place the female first so that the sustainability of the offspring is maintained. According to research of the Aenasius bambawalei parasitoid,5 the temperature in a 24 h treatment had significant effects on its offspring sex ratio, i.e., the stable temperature. The offspring sex ratio is another critical index of parasitic fitness.
CONCLUSION
The highest longevity for adult Hadronotus leptocorisae parasitoids is at a temperature of 25°C. There was a correlation (0.636) between the maintenance tube size with the longevity of adult H. leptocorisae at room temperature but not at 25°C. When rearing the parasitoid H. leptocorisae at 25°C without a specific size tube, the survival ability of H. leptocorisae eggs until they emerge as adults was greater than 78%.
SIGNIFICANCE STATEMENT
This study identifies the appropriate maintenance tube size and rearing room temperature for the longevity of the adult Hadronotus leptocorisae parasitoid that can increase the oviposition time of a female parasitoid. This study will help researchers to identify mechanisms to help the adult parasitoid survive, so that oviposition time and potential for being a biological control agent will increase. Thus, a reduction will be seen in the issue of failing biological agents.
ACKNOWLEDGMENTS
Authors would like to thank the Direktorat Jenderal Pendidikan Tinggi Kementerian Pendidikan dan Kebudayaan that provided the Pendidikan Pascasarjana Dalam Negeri (BPPDN) scholarships with contract number 8222/A4.2/KP/2015 and the research arm of Penelitian Disertasi Doktor with contract number 1488a/PL25/PL/2017.
REFERENCES
- Maulina, F., N. Nelly and Hidrayani, 2014. Potensi pengembangan parasitoid telur walang sangit (Leptocorisa oratorius F.) di Kabupaten Padang Pariaman. Prosiding Seminar Nasional Kebijakan dan Pengembangan Teknologi Hilirisasi Dalam Upaya Peningkatan Nilai Tambah Produk Pertanian, December 3, 2014, Payakumbuh State Agriculture of Polytechnic, pp: 28-37.
- Maulina, F., N. Nelly, Hidrayani and H. Hamid, 2016. Species diversity and parasitization of rice bug (Leptocorisa oratorius Fabricus) egg parasitoidsin Tanah Datar Regency, West Sumatra. Pros. Sem. Nas. Masy. Biodiv. Indon., 2: 109-112.
Direct Link - Qodir, H.A., N. Maryana and Pudjianto, 2017. Biology of Scelio pembertoni Timberlake (Hymenoptera: Scelionidae) on eggs of Oxya japonica (Thunberg) (Orthoptera: Acrididae). J. Entomol. Indon., 14: 58-68.
CrossRefDirect Link - Zhang, J., J. Huang, Y. Lu and T. Xia, 2016. Effects of temperature and host stage on the parasitization rate and offspring sex ratio of Aenasius bambawalei Hayat in Phenacoccus solenopsis Tinsley. Peer J., Vol. 4.
CrossRefDirect Link - Mironidis, G.K. and M. Savopoulou-Soultani, 2009. Development, survival and growth rate of the Hyposoter didymator-Helicoverpa armigera parasitoid-host system: Effect of host instar at parasitism. Biol. Control, 49: 58-67.
CrossRefDirect Link - Regniere, J., J. Powell, B. Bentz and V. Nealis, 2012. Effects of temperature on development, survival and reproduction of insects: Experimental design, data analysis and modeling. J. Insect Physiol., 58: 634-647.
CrossRefDirect Link - Bilodeau, E., J.F. Guay, J. Turgeon and C. Cloutier, 2013. Survival to parasitoids in an insect hosting defensive symbionts: A multivariate approach to polymorphic traits affecting host use by its natural enemy. PLoS ONE, Vol. 8, No. 4.
CrossRefDirect Link - Maulina, F., N. Nelly, Hidrayani and H. Hamid, 2016. Parasitization levels and temperature tolerance of rice bug (Leptocorisa oratorius Fabricus) egg parasitoids from West Pasaman Regency, West Sumatra: Effort of mass rearing of biological agents. In Transforming Awarennes of the Important of Sustainability through Joint Action. Berjaya Hotel Colombo, Sri Lanka, October 20-22, 2016. SAFE Network, pp: SAT-50.
- Tena, A., F.L. Wackers, G.E. Heimpel, A. Urbaneja and A. Pekas, 2016. Parasitoid nutritional ecology in a community context: The importance of honeydew and implications for biological control. Curr. Opin. Insect Sci., 14: 100-104.
CrossRefDirect Link - Maulina, F. and Muflihayati, 2013. Conservation of Diadegma semiclausum Hellen. parasitoids as biological control to Plutella xylostella Linn. with adult food exploration. Int. J. Adv. Sci. Eng. Inform. Technol., 3: 6-8.
CrossRefDirect Link - Fei, M., R. Gols, F. Zhu and J.A. Harvey, 2016. Plant quantity affects development and survival of a gregarious insect herbivore and its endoparasitoid wasp. PLoS ONE, Vol. 11, No. 3.
CrossRefDirect Link - Anggraeni, T., A. Jamili and Umrah, 2010. Perilaku dan penentuan oviposisi dari parasitoid telur Hadronotus leptocorisae (Hymenoptera) pada telur hama bulir padi Leptocorisa acuta (Hemiptera). Biocelebes, 4: 76-79.
Direct Link