
Dalal M. Aljedani
Department of Biological Sciences (Zoology), Faculty of Science, University of Jeddah, Jeddah, Saudi Arabia
LiveDNA: 966.24834
Journal of Entomology
Year: 2018 | Volume: 15 | Issue: 3 | Page No.: 114-124







ABSTRACT
Background and Objective: The honey bee has high economic importance as a successful pollinator in several agricultural crops worldwide and as a source of valued products. The objective of the current study was to and determine the effect of the difference in nutrition in the embryonic stages on the fat body and malpighian tubules of Apis mellifera jemenatica range in different ages. Materials and Methods: In the larval stage of which two ages, the third and fifth have been studied in both the queen and the worker; as queen larvae were fed an intensive diet of royal jelly throughout the larval stage evolution, while the worker larvae were fed royal gel in the first three days only, but in fifth age, it was fed a mixture of honey and bee bread. Results: Several differences were found from one developmental stage to another. Fatty bodies spread throughout the body and fill the internal spaces between the organs. Where fatty bodies increase the size of trophocyte cells with the accumulation of fatty substances and waste inside them with gaps. Either in the malpighian tubules in worker were characterized by the presence of cubic cells with clear nuclei and the composition of these cells based on the basement membrane with the presence of gaps within the cells and the inner cavity is narrow, but in the queen is characterized by the size of more than the worker. Conclusion: It seem very useful to link the study of histological structure recorded at the cellular or tissue levels with the development of age and with changes at the community levels.
PDF Abstract XML References Citation
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dalal M. Aljedani, 2018. Comparing the Histological Structure of the Fat Body and Malpighian Tubules in Different Phases of Honeybees, Apis mellifera jemenatica (Hymenoptera: Apidae). Journal of Entomology, 15: 114-124.
DOI: 10.3923/je.2018.114.124
URL: https://scialert.net/abstract/?doi=je.2018.114.124
DOI: 10.3923/je.2018.114.124
URL: https://scialert.net/abstract/?doi=je.2018.114.124
INTRODUCTION
The honey bee Apis mellifera (Linnaeus 1758) (Hymenoptera: Apidae) has high economic importance. This species has been used as a successful pollinator in several agricultural crops worldwide and as a source of valued products such as wax, honey, propolis and royal jelly1. Apis mellifera jemenatica is the main honey bee strain of the Arabian Peninsula, also spread in some parts of Africa2. Many organisms undergo distinct shifts in the history of life that are associated with changes in physiology3. Honeybee life is characterized by a profound age-related change in behavior phenotype. Usually, the hive bees remain inside the nest where they eat and feed and tend to the queen and keep the colon clean. After that, bees become slave laborers and leave the colony on daily trips to collect food4. This behavioral change is related to structural and biochemical changes in the body3.
The insect fat body is a multifunctional device that has a role in a variety of metabolic processes, including storage of proteins, fats and carbohydrates, which are precursors to metabolism in other organs5-7. This tissue is a major organ in the medium metabolism of insects, the majority of hemolymph proteins are synthesized in this tissue, which also displays the functions of fat, carbohydrate and protein storage8. In addition, the obese body of insects displays other cell specialties, such as urinary cells, hemoglobin cells, melanocytes and chromatic cells8. The fatty body is divided into a vital part, surrounds the viscera and occupies the abdominal cavity of the insects and a wall part9,10.
All in multicellular organisms there is a device that collects and disposes of waste outside the body. Production systems vary in various animals, including insects and mammals. Malpighian tubules are the main devices for insect removal. It is similar to that of the kidney invertebrate animals10,11. In addition to osmoregulation, malpighian tubules are also involved in organic material transfer, body defense system and chemical detoxification processes12. The malpighian tubules (located at the anterior end of the intestine) opens in the gate cavity directly behind the pyloric valve area13-15. Morphologically, are thin, finger-like extensions connected to the intestinal tract between the midgut and hindgut11. Urine is produced in the insects in its initial stages of malpighian tubules, by the isosmotic filtrate from the hemolymph, which carries the metabolic and toxic compounds of the gut. The final stage of the rectal output process usually occurs, where some ions and water are reabsorbed16. The excretion occurs by the secretion of ions and organic molecules from the dermal into the lumen of the tubule. Of the lumen, the contents are modified as they pass through the Malpighian and hindgut tubules until they are secreted11. Also, it is responsible for producing isosmotic filtrate of dermal, main urine, which carries excretory products of metabolism and toxic compounds in the backside11. Which are responsible for the secretion of substances in the body. In most types of insects, the output system involves a variable number of sediments. These play an important role in the detoxification process by actively functioning to dispose of substances that are not metabolized and are in excess of the organism17. In addition, to excretion and their impairment could help contribute to increased colony losses in the face of environmental stressors14. However, this needs further exploration. The Malpighian tubules play a variety of protective roles18. The malpighian tubules can also mount an immune response independent of the fat body19. Many studies have examined the structure and function of these organisms, but there is no scientific study that seriously addressed the study of the effect of nutrition in the stages of embryonic development, especially in the larval stage where the nutrition is different in the queen in the workers. This difference is followed by several changes in the external appearance or internal structure. Current study to trace differences in the histological structure of the fat body and malpighian tubules of Apis mellifera jemenatica range in different ages.
MATERIALS AND METHODS
The individuals used in the current study was transferred from the apiary directly to the laboratory where the study was conducted under laboratory conditions. Local bee race (Apis mellifera jemenatica)20, which is the smallest race of yellow bees, were selected for the present study. The most obvious distinguishing morphological characteristics in this race are that worker bees have relatively short wings and legs and have yellow rings on their abdomens, whereas queens have relatively large yellow abdomens. Two medium-sized hives of local pure bees were used to collect individuals from different castes at different ages for histological studies of fat body and malpighian tubules.
Individuals used in the study
| • | Larval stage: In the larval stage; of which two ages, the third and fifth have been studied in both the queen and the worker; as queen larvae were fed an intensive diet of royal jelly throughout the larval stage evolution, while the worker larvae were fed royal gel in the first three days only, but in the fifth age, it was fed a mixture of honey and bee bread21 |
| • | Pupa stage: The age (in days) of the pupa stages was determined by changes in body and eye color |
For queens, one and three-day-old pupae were selected and for workers, one and seven- day-old pupae were selected.
| • | Adult stage: | |
| • | Queen’s adult queens (virgin queens) were selected for this study | |
| • | Workers: Adult workers were examined at three age levels that varied by the type of food consumed. For newly emerged workers, the age levels were as follows: 0-12 h prior to the onset of feeding, nurse workers which consume large amounts of the pollen stored inside the hexagonal cells, particularly over days 3-6 (protein feeding) and foragers which collect and feed on nectar and pollen and they consume a larger amount of nectar than the nurse bees (carbohydrate feeding) | |
Determining ages of worker bees: One comb was selected and dated, containing one- day-old eggs. After three days, the eggs hatched and larvae continued to develop over five instars (13 days after oviposition or 6 days after the hexagonal cells are covered), then became pupae (one-day-old pupae). Worker bee age was determined by identifying newly emerged worker bees (either during exit from a cell or immediately after exit) on a brood frame. A temperature of 33°C and a relative humidity of 65% was used to obtain 24 h old workers. The newly emerging worker bees were marked with colored markers to track the different ages.
Honeybee queen breeding: Waxy cups grafting method (also known as the Doolittle), the method was used to breed and obtain queens of different ages.
Histology: For the histological studies, the samples were prepared according to the historesin technique as follows: individuals of each age were collected and placed in a preservative solution. Were fixed in 10% paraformaldehyde in 0.1 M phosphate buffer solution (pH 7.2). The material was dehydrated in an ascending series of 70, 80, 90 and 95% ethanol baths lasting 20 min each and transferred to the resin solution for 72 h at 4°C. Finally, the material was transferred to molds which were then filled with resin containing a catalyst and sealed with a metal support for microtome. After polymerization, the blocks were sliced using a microtome. The sections were mounted on slides and stained with hematoxylin and eosin. Samples were then examined and photographed using an Olympus microscope (Olympus-Bx41)17,11,22.
RESULTS AND DISCUSSION
Fat body
Larval stage: In the results of the present study in the larval instar of honey bees, the fatty bodies are characterized by two types of cells: Trophocyte of fat body (Tr) and Oenocyte of fat body (On) (Fig. 1). In the 3rd larval instar of honey bees, queen fatty bodies spread throughout the body and fill the internal spaces between the organs. Where fatty bodies increase the size of fat cells (Tr) with the accumulation of fatty substances and waste inside them with gaps in Fig. 1a. But, in the 3rd larval instar of honey bee's worker the fat cells of (Tr) large size and the presence of spaces or gaps in the internal small with the clarity of the nuclei. On cells are characterized by their small numbers, which are distributed among the (Tr) cells (Fig. 1b). In the 5fh larval instar of honey bees queen, the boundaries between the fat cells (Tr) are clearly marked, the gaps inside the are few, the accumulation of the external substances inside and the lack of clarity of the nuclei (Fig. 1c). For the worker, it is similar to honey bees queen (Fig. 1d). This similar to what was found6 the insect fat body is a diffuse organ that fills the body cavity and consists of mesodermal cells known as trophocytes or fat body cells. As well as to trophocytes, bees contain oenocytes, another cell type of ectodermal origin found scattered throughout the fat body, this agrees with Martins et al.23. Also, agrees with Ahmed et al.24, when he found that the trophocyte is the main cell type of the insect fat body. The shape, appearance and volume of the trophocytes vary and are widely depending on the development stage and nutritional state of the insect, however, a marked characteristic of this type of the cell is a cytoplasm filled with lipidic deposits except for a thin portion at the cell’s periphery, a ring surrounding the nucleus and dispersed islands between the lipid droplets. Few inclusions are present at these regions of the cytoplasm. The trophocytes increase in size and probably also in number during the postembryonic feeding period. When they become the largest cells of the insect’s body. In addition to during the larval phase, it is involved in the metabolism of lipids and carbohydrates and in the synthesis and secretion of proteins, which are released in large quantities into the hemolymph in the last larval stage to drive and carry out the metamorphosis25. Both in the pre-embryonic and adult phases, the fat body is involved in the accumulation of toxic materials, such as urate26. The fat body is more prominent in immature insects, particularly holometabolous insects in Apis mellifera larvae27.
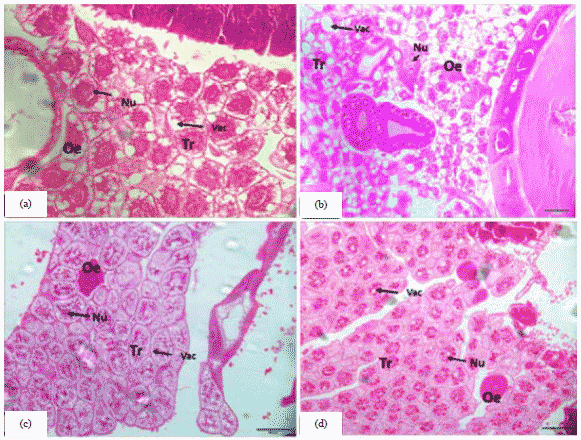 | |
| Fig. 1(a-d): | Cross section of the fat body in the honey bee larva, (a) 3rd day old honey bee queen larva (20 µm) , (b) 3rd day old honey bee worker larva (20 µm), (c) 5th day old honey bee queen larva (50 µm) and (d) 5th day old honey bee worker larva (50 µm) |
Maybe that's because the abdomen contains the bulk of the honey bee’s fat body, which is a site of nutrient synthesis and storage, including carbohydrate, protein, lipid macronutrients and important micronutrients. The insect peripheral fat body is located just beneath the cuticle and extends over most of the body28. The results of previous studies have proved a high level of protein accumulation in the form of vitellogenin as a major constituent of the honeybee fat body29. Additionally, the fatty body consists of thin layers or filaments. Generally, one or two thick cells or the small nodules are attached to the hemocele through connective tissue and trachea. Within a species, the structure of this tissue is fairly stable, but it can have significant differences between insects of different orders8. The fatty body is spread in most of the body cavity. This tissue extends into all regions of the body, including the external appendages of the insects. The fat body is more abundant in the abdominal region and is formed by thin strings or nodules that are suspended in the insect hemocoel and involve connective tissue and the trachea30.
Pupal stage: In the results of the present study, it was found that the honey bee pupa in 1st day old honey bee queen pupa the cellular boundary between the fat cells tr is dissipated with the accumulation of waste inside (Fig. 2a) and in 1st day old honey bee worker pupa the distances between the (Tr) cells are spaced, filled with internal debris and the gaps are disintegrated (Fig. 2b). Also, in the 3rd day old honey bee queen pupa, similar to what existed in the first age of the pupa. Also, it is clear that the waste is decreased and the spacing of cells is wide Fig. 2c. But, in the 7th day old honey bee worker pupa the accumulation and disintegration of fat cells in the (Tr) and the gaps inside them are obvious (Fig. 2d). The fat body contains impurities consisting of several compounds, mainly fats, proteins and glycogen. The amount and quality of the material accumulated in the cytoplasm vary according to the functional stage of the organism31. In a study by Zara and Caetano32, they show three different cell types: trophocyte, urate and the oenocyte. The trophocyte is the most abundant cell type in the fat body; the cells are rounded and show nuclei with varying sizes and shapes. Several electron-lucid spheres, lipid droplets and some eosinophil granules can be observed in their cytoplasm. The urate cells, seemingly more abundant next to the ventriculus, are distributed among the trophocytes throughout the body cavity.
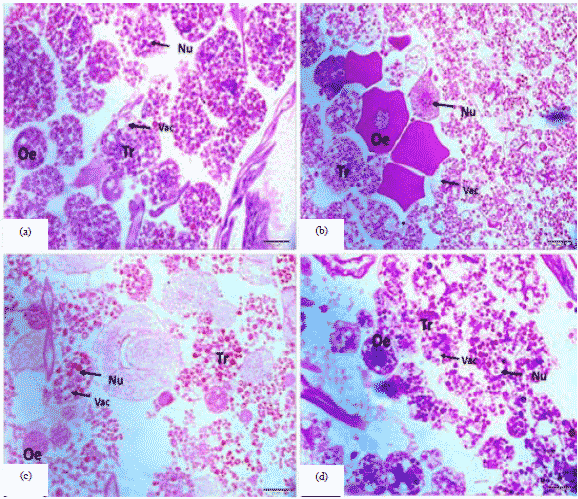 | |
| Fig. 2(a-d): | Cross section of the fat body in the honey bee pupa, (a) 1st day old honey bee queen pupa (20 µm), (b) 1st day old honey bee worker pupa (20 µm), (c) 3rd day old honey bee queen pupa (20 µm) and (d) 7th day old honey bee worker pupa (20 µm) |
These cells, recognizable based on their slightly polygonal morphology, show a central nucleus with a large amount of heterochromatin. Several small and lucid vesicles can be observed in their cytoplasm. The last cellular type found in the fat body the oenocytes, present a rounded shape although they can be slightly deformed due to pressure exerted by neighboring cells. They show a rounded and large, centrally located nucleus, with a great amount of heterochromatin. The cytoplasm is strongly stained by both hematoxylin and eosin (HE); it appears flocculated, similar to small vacuoles.
Adult stage: The results of present study showed that the adult honey bee queen, the size of the cells in the triglyceride cells increased and elongated and the gaps inside them were clear (Fig. 3a, b). In the honey bee worker, the adult stage the increase of (Tr) fat cells and internal gaps as well as accumulation of waste, which works on the cells (On) to remove toxins from the body (Fig. 3c, d). This is consistent with what is mentioned33 when they found the oenocytes can act in detoxification processes. However, there is a specific region where the fat body is associated with two or more different cell types, playing the role of only one and in bees, it may play functions homologous with the hepatocytes and nephrocytes of vertebrates34,35. The fat body of insects consists of two main types of cells: trophocyte and oenocyte. Although these cells were apparently associated in a mass of tissue, the former has a mesodermal origin and the latter, ectodermal, presenting different morphology and function9,31.
In the results of the current study found that the histological structure of fatty body varies between the worker and queen honeybees in various stages and may be due to the nature of the tasks performed by individuals in the colony. This is consistent with what many researchers have thought honeybee workers are essentially sterile female helpers that make up the majority of individuals in a colony. Workers display a marked change in physiology when they transition from in-nest tasks to foraging. Recent technological advances have made it possible to unravel the metabolic modifications associated with this transition.
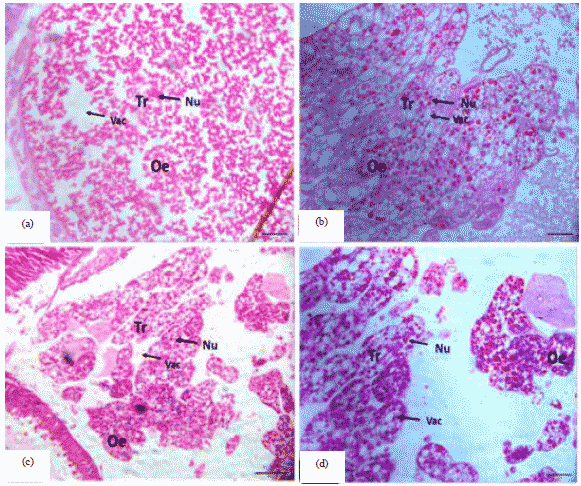 | |
| Fig. 3(a-d): | Cross section of the fat body in the adult honey bee, (a) 20 µm, (b) 20 µm adult queen, (c) 50 µm and (d) 20 µm adult worker |
The honeybee fat body is more developed in nest bees compared to forager bees4 and a decrease in abdominal lipids has been shown to precede the onset of foraging4. These observations indicate that the biochemistry of the abdomen is profoundly remodeled during the transition from in-nest tasks to foraging4. For nurse workers, the trophocytes of fat body incubated only in culture media (control) were IO-positive around the mitochondria and in heterogeneous inclusions of the cytoplasm. For nurse workers, the trophocytes of fatty body cultured only in plain media (control) were AS-positive with a powdery granulation scattered in the cytoplasm and around inclusions of low electron-density, which are probably lipids. For the virgin queen, the AP activity on control fatty body trophocytes appeared as powdery granules scattered in the cytoplasm and condensed around lipid droplets31. Lipids are the main compounds stored in the fatty body trophocytes of nurse workers and virgin queens. In the controls, lipids appeared as small dots around mitochondria in both cases, suggesting a role for these organelles in lipid metabolism as seen in several kinds of tissues, primarily the ones that store lipids and glands that secrete lipid-like substances. In a study by Seehuus et al.36, they observation than both foragers, treated bees and the reversion bees all showed a high proteolysis activity. This result probably reflects a degradation of protein storage in the forager fat body. As a preparation for foraging, workers use their fat bodies as a source of energy, fat and proteins and thereby reduce their demand for nutrients from the hive. Protein synthesis is one of the most energy consuming cellular processes, devouring an estimated 50% of total cellular energy. Limiting the protein synthesis in foragers, therefore, seems like a good energy saving solution on both the individual and colony level. A reduced translation rate would reduce not only normal protein production but also the flux of damaged proteins through various pathways.
Malpighian tubules
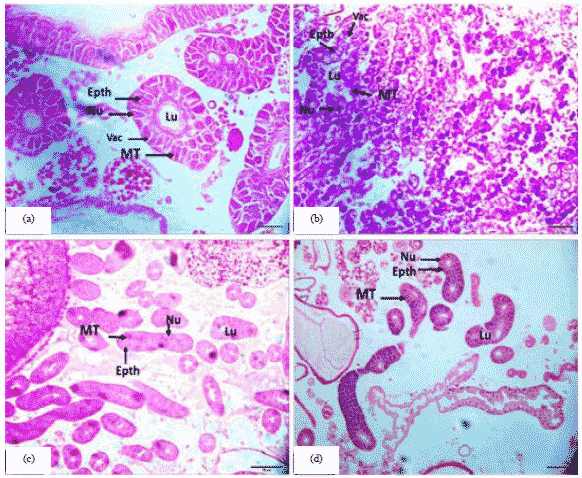
Larval stage: In the results of the present study, it was found in the 3rd day old honey bee queen larva was characterized by the existence of cubic cells in large size and a clear nucleus and the presence of gaps inside the cells and the inner cavity is narrow (Fig. 4a).
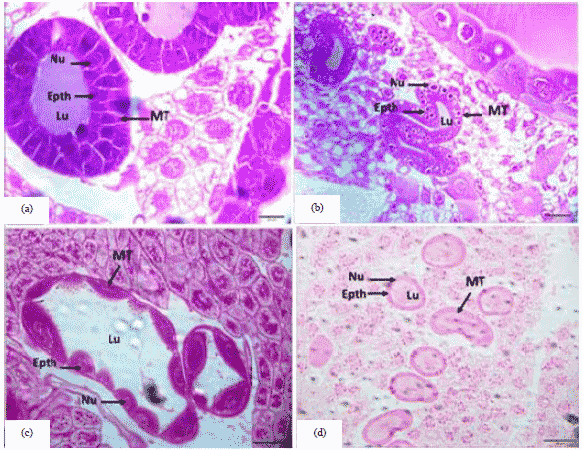 | |
| Fig. 4(a-d): | Cross section of the malpighian tubules in the honey bee larva, (a) 3rd day old honey bee queen larva (20 µm), (b) 3rd day old honey bee worker larva (20 µm), (c) 5th day old honey bee queen larva (50 µm) and (d) 5th day old honey bee worker larva (50 µm) |
The 3rd day old larva in honey bee's worker, it was characterized by the presence of cubic cells with clear nuclei and the composition of these cells based on the basement membrane with the presence of gaps within the cells and the inner cavity was narrow. But in the queen was characterized by the size of more than worker (Fig. 4b). Also, the 5th day old larva in honey bees queen epithelial cells elongation with the accumulation of waste and its accumulation in the inner cavity and secondary is larger in size but its boundaries are unclear (Fig. 4c). In the 5th day old honey bee worker larva was similar to the larva of the third age, but the cells have become the border between them is not clear and the inner cavity was wide (Fig. 4d). Therefore, current observations strongly suggested that the during the larval stage of bees, the products absorbed from the hemolymph by Malpighian tubules are accumulated in their lumen until the end of the larval feeding phase. The Malpighian tubules finally excrete their accumulated products when there is a connection between the midgut and the hindgut preceding metamorphosis11. At the end of the larval stage of bees, Malpighian tubules become inoperative and begin their degenerative process. During metamorphosis of bees, new Malpighian tubules differentiate from stem cells located at the insertion of the tubules into the digestive tract11. In a study by Scholes et al.37 and Rangel et al.38, tissue-specific differences in ploidy levels are also documented in the Hymenoptera. Using the honey bee as a model to study endoreduplication in eusocial insects. In accordance with previous findings, malpighian tubules exhibited relatively high ploidy levels, which increased with worker age, this relatively high ploidy in the Malpighian tubules highlights the essential role of endopolyploidy in insect secretory cells39.
Pupal stage: In the 1st day old honey bee queen pupa the epithelial cells are adjoining and overlapping as if they are cellular and the nanoparticles are unclear and the inner cavity is virtually devoid of any components (Fig. 5a). But, in the 1st -day-old honey bee worker pupa the epithelial cells of the malpighian tubules became adjacent and the secondary was unclear and the inner cavity was very round (Fig. 5b). In the 3rd day old honey bee queen pupa similar to the first age of the pupa as it increases as the age approaches its end (Fig. 5c). Also, in the 7th day old honey bee worker pupa the epithelial cells of the malpighian tubules have become dilapidated, the nuclei are unclear, the internal cavity is almost solid (Fig. 5d).
 | |
| Fig. 5(a-d): | Cross section of the malpighian tubules in the honey bee pupa, (a) 1st day old honey bee queen pupa (20 µm), (b) 1st day old honey bee worker pupa (20 µm), (c) 3rd day old honey bee queen pupa (50 µm) and (d) 7th day old honey bee worker pupa (20 µm) |
From the results of the present study, it can conclude that the stage of the pupa phase clearly shows the re-formation of tissues where the bees are holometabolous insects and therefore undergo a complete reorganization of organs during metamorphosis, during pupae phase. In Hymenoptera changes in the midgut begin in the prepupa with the larval epithelium degenerating, leaving only the basement membrane and the regenerative cells. At metamorphosis, the larval midgut epithelium is reabsorbed and substituted for cells from regenerative cells27. From the results of the present study, It was concluded that the stage of the pupa phase clearly shows the re-formation of tissue in the bees.
Adult stage: In the adult queen, the epithelial cells in malpighian tubules between them are unclear and the nuclei scattered. Also, the internal cavity is almost solid (Fig. 6a, b). In general, the results obtained in the present study showed in the adult worker similar to the seventh age in the pupa (Fig. 6c, d). The malpighian tubules of bees showed structural integrity and spherical-shaped nuclei; the interior of the nucleus was filled with euchromatin with some regions presenting heterochromatin around the nuclear envelope and the presence of a nucleolus was observed. Mitochondria exhibited cristae and were found throughout the cell, with a higher concentration in the apical region and within the microvilli; additionally, intact microvilli were observed40. Typical ultrastructural characteristics were observed in the Malpighian tubules for the epithelium appeared to be well preserved and showed no alterations in the basal labyrinth and the normal presence of microvilli in the apical portion. Many mitochondria with cristae could be visualized in the cells41. The results of previous studies have proved, following the metabolism of thiamethoxam, in the body of the bee, the digested products or the undigested molecule will reach the hemolymph and subsequently, the brain and the Malpighian tubules. This organ was responsible for the maintenance of homeostasis and for the detoxification process that eliminates excess substances, even those that have not been metabolized by the body42. Maybe that's because the endocycle is also active in adult tissues such as those present in the malpighian tubules all of which exhibit high levels of endoreduplication43.
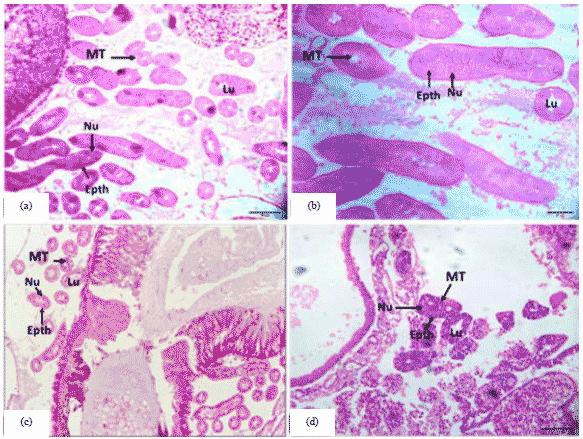 | |
| Fig. 6(a-d): | Cross section of the malpighian tubules in the honey bee, (a) 50 µm, (b) 20 µm adult queen, (c) 50 µm and (d) 50 µm adult worker |
CONCLUSION
The results showed the accuracy and miracles in the composition of such an insect and that changes in nutrition in embryonic stages have a clear effect on cellular structure, as it has been in the histological structure of the fat body and malpighian tubules of queen and worker of the local honey bees races, Apis mellifera jemenitica the various ages of the workers found differences between the honey bee queen and the worker as well as the difference in the different ages.
REFERENCES
- Lopes, M.P., K.M. Fernandes, H.V.V. Tome, W.G. Goncalves, F.R. Miranda, J.E. Serrao and G.F. Martins, 2018. Spinosad‐mediated effects on the walking ability, midgut and Malpighian tubules of Africanized honey bee workers. Pest Manage. Sci., 74: 1311-1318.
CrossRefDirect Link - Aljedani, D.M., 2017. Effects of abamectin and deltamethrin to the foragers honeybee workers of Apis Mellifera jemenatica (Hymenoptera: Apidae) under laboratory conditions. Saudi J. Biol. Sci., 24: 1007-1015.
CrossRefDirect Link - Elekonich, M.M. and S.P. Roberts, 2005. Honey bees as a model for understanding mechanisms of life history transitions. Compa. Biochem. Physiol. Part A: Mol. Integr. Physiol., 141: 362-371.
CrossRefDirect Link - Chan, Q.W., N.S. Mutti, L.J. Foster, S.D. Kocher, G.V. Amdam and F. Wolschin, 2011. The worker honeybee fat body proteome is extensively remodeled preceding a major life-history transition. PLoS One, Vol. 6, No. 9.
CrossRefDirect Link - Sousa, C.S.D., J.E. Serrao, A.M. Bonetti, I.M.R. Amaral, W.E. Kerr, A.Q. Maranhao and C. Ueira-Vieira, 2013. Insights into the Melipona scutellaris (Hymenoptera, Apidae, Meliponini) fat body transcriptome. Genet. Mol. Biol., 36: 292-297.
CrossRefDirect Link - Andrade, G.S., J.E. Serrao, J.C. Zanuncio, T.V. Zanuncio, G.L.D. Leite and R.A. Polanczyk, 2010. Immunity of an alternative host can be overcome by higher densities of its parasitoids Palmistichus elaeisis and Trichospilus diatraeae. Plos One, Vol. 5, No. 10.
CrossRefDirect Link - Ottaviani, E., D. Malagoli and C. Franceschi, 2011. The evolution of the adipose tissue: A neglected enigma. Gen. Compa. Endocrinol., 174: 1-4.
CrossRefDirect Link - Roma, G.C., O.C. Bueno and M.I. Camargo-Mathias, 2010. Morpho-physiological analysis of the insect fat body: A review. Micron, 41: 395-401.
CrossRefDirect Link - Nocelli, R., P. Cintra-Socolowski, T. Roat, E. Silva-Zacarin and O. Malaspina, 2016. Comparative physiology of Malpighian tubules: Form and function. Insect Physiol., 2016: 13-23.
CrossRefDirect Link - Wang, J., L. Kean, J. Yang, A.K. Allan, S.A. Davies, P. Herzyk and J.A. Dow, 2004. Function-informed transcriptome analysis of Drosophila renal tubule. Genome Biol., Vol. 5, No. 9.
CrossRef - Tutkun, E., 2016. Ari Akari (Varroa destructor) mucadelesinde timol’un kullanilmasi. [Using of thymol in fighting against varroa destructor]. Aricilik Arastırma Dergisi, 8: 1-5.
Direct Link - Traynor, K.S., M. Andree, E.M. Lichtenberg, Y. Chen, C. Saegerman and D.L. Cox-Foster, 2007. Colony Collapse Disorder (CCD) and bee age impact honey bee pathophysiology. PloS One, Vol. 12, No. 7.
CrossRefDirect Link - Dow, J.A. and S.A. Davies, 2006. The Malpighian tubule: Rapid insights from post-genomic biology. J. Insect Physiol., 52: 365-378.
CrossRefDirect Link - Arab, A. and F.H. Caetano, 2002. Segmental specializations in the Malpighian tubules of the fire ant Solenopsis saevissima Forel 1904 (Myrmicinae): An electron microscopical study. Arthropod Struct. Dev., 30: 281-292.
CrossRefDirect Link - De Almeida Rossi, C., T.C. Roat, D.A. Tavares, P. Cintra‐Socolowski and O. Malaspina, 2013. Effects of sublethal doses of imidacloprid in malpighian tubules of africanized Apis mellifera (Hymenoptera, Apidae). Microscopy Res. Techn., 76: 552-558.
CrossRefDirect Link - Dow, J.A.T., 2009. Insights into the Malpighian tubule from functional genomics. J. Exp. Biol., 212: 435-445.
CrossRefDirect Link - McGettigan, J., R.K.J. McLennan, K.E. Broderick, L. Kean and A.K. Allan et al., 2005. Insect renal tubules constitute a cell-autonomous immune system that protects the organism against bacterial infection. Insect Biochem. Mol. Biol., 35: 741-754.
CrossRefDirect Link - Aljedani, D.M. and R.M. Almehmadi, 2016. Effects of some insecticides on longevity of the foragers honey bee worker of local honey bee race Apis mellifera jemenatica. Electr. Phys., 8: 1843-1849.
CrossRefDirect Link - Almehmadi, R.M., A.A. Al-Ghamdi and D.M. Aljedani, 2015. The histological structure of the pyloric valve in the yemeni honey bees queen and worker (Indigenous) Apis Mellifera jemenatica (Hymenoptera: Apidae). Life Sci. J., 12: 145-153.
Direct Link - Fontanetti, C.S., M.I. Camargo-Mathias and B. Tiritan, 2004. The fat body in Rhinocricus padbergi (Diplopoda, Spirobolida). Iheringia. Serie Zool., 94: 351-355.
CrossRefDirect Link - Martins, G.F., J.M. Ramalho-Ortigao, N.F. Lobo, D.W. Severson, M.A. McDowell and P.F.P. Pimenta, 2011. Insights into the transcriptome of oenocytes from Aedes aegypti pupae. Mem. Inst. Oswaldo Cruz, 106: 308-315.
CrossRefDirect Link - Abdalla, F.C. and C.E. da Costa Domingues, 2015. Hepato-nephrocitic system: A novel model of biomarkers for analysis of the ecology of stress in environmental biomonitoring. PloS One, Vol. 10, No. 7.
CrossRefDirect Link - Lowe, T., R.J. Garwood, T.J. Simonsen, R.S. Bradley and P.J. Withers, 2013. Metamorphosis revealed: Time-lapse three-dimensional imaging inside a living chrysalis. J. Royal Soc. Interface, Vol. 10, No. 84.
CrossRefDirect Link - Nilsen, K.A., K.E. Ihle, K. Frederick, M.K. Fondrk, B. Smedal, K. Hartfelder and G.V. Amdam, 2011. Insulin-like peptide genes in honey bee fat body respond differently to manipulation of social behavioral physiology. J. Exp. Biol., 214: 1488-1497.
CrossRefDirect Link - Chapman, R.F., 1998. The Insects: Structure and Function. 1st Edn., Cambridge University Press, Cambridge.
Direct Link - Paes-De-Oliveira, V.T., B. Berger, S.B. Poiani, Z.L.P. Simoes and C. da Cruz‐Landim, 2013. Effects of treatment of the fat body trophocytes of Melipona quadrifasciata anthidioides nurse workers and virgin queens in culture by juvenile hormone III and ecdysterone (20‐HE). Microscopy Res. Techn., 76: 20-27.
CrossRefDirect Link - Zara, F.J. and F.H. Caetano, 2004. Ultramorphology and histochemistry of fat body cells from last instar larval of the Pachycondyla (= Neoponera) villosa (Fabricius) (Formicidae: Ponerinae). Braz. J. Biol., 64: 725-735.
CrossRefDirect Link - Klionsky, D.J., F.C. Abdalla, H. Abeliovich, R.T. Abraham and A. Acevedo-Arozena et al., 2012. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy, 8: 445-544.
PubMedDirect Link - Abdalla, F.C., E.C.M. Silvia-Zacarin and M.J. Costa, 2014. Biomarcadores morfologicos em abelhas nativas. Ecotox, 8: 44-48.
Direct Link - Billen, J., 1991. Ultrastructural organization of the exocrine glands in ants. Ethol. Ecol. Evol., 3: 67-73.
CrossRefDirect Link - Seehuus, S.C., S. Taylor, K. Petersen and R.M. Aamodt, 2013. Somatic maintenance resources in the honeybee worker fat body are distributed to withstand the most life-threatening challenges at each life stage. PloS One, Vol. 8, No. 8.
CrossRefDirect Link - Scholes, D.R., A.V. Suarez and K.N. Paige, 2013. Can endopolyploidy explain body size variation within and between castes in ants? Ecol. Evol., 3: 2128-2137.
CrossRefDirect Link - Rangel, J., K. Strauss, K. Seedorf, C.E. Hjelmen and J.S. Johnston, 2015. Endopolyploidy changes with age-related polyethism in the honey bee, Apis mellifera. PloS One, Vol. 10, No. 4.
CrossRefDirect Link - Edgar, B.A. and T.L. Orr-Weaver, 2001. Endoreplication cell cycles: More for less. Cell, 105: 297-306.
CrossRefDirect Link - Friol, P.S., A.F. Catae, D.A. Tavares, O. Malaspina and T.C. Roat, 2017. Can the exposure of Apis mellifera (Hymenoptera, Apiadae) larvae to a field concentration of thiamethoxam affect newly emerged bees? Chemosphere, 185: 56-66.
CrossRefDirect Link - Catae, A.F., T.C. Roat, R.A. de Oliveira, R.C.F. Nocelli and O. Malaspina, 2014. Cytotoxic effects of thiamethoxam in the midgut and malpighian tubules of Africanized Apis mellifera (Hymenoptera: Apidae). Microscopy Res. Techn., 77: 274-281.
CrossRefDirect Link - Lilly, M.A. and R.J. Duronio, 2005. New insights into cell cycle control from the Drosophila endocycle. Oncogene, 24: 2765-2775.
CrossRefDirect Link