
Kehinde Temitope Olatayo
Department of Zoology, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
LiveDNA: 234.21488
Journal of Entomology
Year: 2017 | Volume: 14 | Issue: 5 | Page No.: 228-233







ABSTRACT
Background and Objective: Assessments of insect diversity in human landscapes often focus on local scale (alpha) diversity with little attention given to species turnover (beta diversity) of these organisms at the landscape scale. This is particularly crucial for highly mobile insects such as flower visiting insects. This study, therefore, assessed the effects of four land use types (agricultural land, grazed grassland, mown grassland and secondary forest) on the alpha and beta diversity of flower-visiting insects in Ile-Ife, Nigeria. Materials and Methods: Three replicates of each land use type were selected. Flower-visiting insects were sampled using coloured pan traps and aerial netting along 100 m transects on each study site. Additive partitioning was used to derive alpha and beta diversity of flower-visiting insects. Results: About 40, 57, 54 and 51 species of flower-visiting insects were obtained from agricultural land, grazed grassland, mown grassland and secondary forest habitats, respectively. Beta diversity had significantly higher percentage contribution to the total diversity compared to alpha diversity. Furthermore, while land use had no significant effect on alpha diversity, there was a marginally significant effect on beta diversity. This underscores the very crucial place of beta diversity in holistic assessments of flower-visiting insect diversity. Conclusion: Species turnover of flower-visiting insects provides information on the sensitivity of these organisms to disturbance such as land use change.
PDF Abstract XML References Citation
Received: November 18, 2017;
Accepted: January 01, 2018;
Published: January 16, 2018
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Kehinde Temitope Olatayo, 2017. Alpha and Beta Diversity of Flower-visiting Insects in Different Land Use Types. Journal of Entomology, 14: 228-233.
DOI: 10.3923/je.2017.228.233
URL: https://scialert.net/abstract/?doi=je.2017.228.233
DOI: 10.3923/je.2017.228.233
URL: https://scialert.net/abstract/?doi=je.2017.228.233
INTRODUCTION
Local scale (alpha diversity) assessments does not sufficiently account for turnover in species diversity at both spatial and temporal scales1,2. Beyond alpha diversity (average within plot diversity) of species, additive partitioning of diversity has shown that there are different components that make up the diversity of flora and fauna communities1,3. These components may be spatial, γ-diversity: Total diversity across plots and β-diversity: The difference between (γ) total and (α) local diversity, which measures variation in species composition between plots1,4 or temporal: β-diversity based on temporal turnover of species richness3.
A comprehensive approach that accounts for all these components of diversity is particularly important for highly mobile groups like insects which utilize several habitats in both natural and modified landscapes5. However, much of the growing literature on the spatial and temporal patterns and consequences of taxonomic homogenization has focused on a relatively restricted set of taxa, with little known for other groups such as insects6. Increased attention is, therefore, required for insects especially functionally important insect taxa such as flower-visiting insects which play crucial role in pollination in both agricultural and natural ecosystems7. More importantly due attention is required for insects in tropical ecosystems which continue to face escalating threats for a synergy of anthropogenic disturbances8,9. The turnover of flower-visiting insects is particularly crucial in tropical ecosystems due to long flowering season of many flowering plants in the tropics which sometimes overlap between the seasons. While local scale alpha diversity of flower-visiting insects have been monitored across different land use types in the tropics5,10, little is known about other ecologically important components of the diversity of these groups such as their beta diversity across land use types. Additive partitioning (summation of within and among communities diversity) was used to determine the effect of land use on the alpha and beta diversity of flower-visiting insects in four land use types (agricultural land, grazed grassland, mown grassland and secondary forest habitats).
MATERIALS AND METHODS
Study area: The study was carried out in Ile-Ife, Southwest Nigeria with the following coordinates latitudes 6°57’05"N to 7°35'19"N and longitudes 4°20'41"E to 4°46'21"E (Fig. 1). Ile-Ife lies within the tropical rainforest vegetation of Nigeria which lies within the West African Forest biodiversity hotspot. This region like most other biodiversity hotspots is enriched with high flora and fauna diversity but faced with enormous threats and is, therefore, of high priority for conservation11. The major seasons in the study area are the rainy season characterised by heavy rains and dry season with little or no rain and dry environmental conditions.
Four land use types comprising of three replicates of each land use type were selected for the study within the study area. These include; agricultural lands which were managed by local peasant farmers. The dominant plants on these sites were Manihot esculenta, Aspilia africana, Chromolaena odorata, Sida acuta, Mangifera indica etc. The main anthropogenic disturbances on these sites were from the activities of the farmers such as tillage, weeding and use of agrochemicals. The second land use type was grazed grasslands which had the following plant species Tridax procumbens, Sida acuta, Corchorus sp., Mimosa pudica, Talinum triangulare, Aspilia africana, Indigofera sp., Chromolaena odorata, Stachytarpheta cayennensis, Ageratum conyzoides, Ipomoea sp. and Vernonia cinerea. The grasslands in these study sites were sparingly grazed by a few cattle which resulted in minimal level of disturbances to the grasses and flowering plants on the sites. The third land use type was the mown grasslands which had similar flowering plants as the grazed grassland. The plants on these sites were frequently mown and trampled by human movement. Lastly, secondary forest sites were also sampled during this study. The plants on these sites were Azadirachta indica, Hildegardia barteri, Leucaena leucocephala, Chromolaena odorata and Aspilia africana. These sites were mainly disturbed from deforestation and occasional bush burning.
Sampling of flower-visiting insects: Sampling was carried out on the agricultural lands, mown grasslands and secondary forest habitats between May, 2014 and April, 2015 and on the grazed grass lands between July, 2016 and February, 2017. Sampling was carried out on a monthly basis covering both the rainy and dry seasons. Two sampling methods were combined in the sampling. The first sampling method was observation along two 100 m×5 m transects on each of the study sites. Flower-visiting insects foraging on flowering plants along the transects were recorded and collected with aerial nets. Nine coloured pan traps of 1.5 L capacity made up of three each of colours yellow, white and blue were placed randomly on each study sites. The pan traps were filled with water to half of their capacity and a few drops of detergent added to the water to break the surface tension and enhance insect trapping. Pan traps were left active on the sites for 48 h after which they were removed and the trapped insects were stored in 75% ethanol.
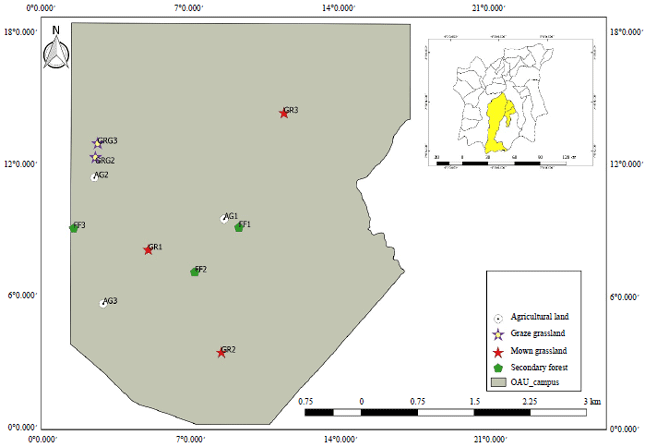 | |
| Fig. 1: | Study sites and land use types in Ile-Ife, Nigeria (Inset map of Osun State showing Ile-Ife, Nigeria). AG1: Agricultural land 1, AG2: Agricultural land 2, AG3: Agricultural land 3, GRG1: Grazed grassland 1, GRG2: Grazed grassland 2, GRG 3: Grazed grassland 3, GR1: Mown grassland 1, GR2: Mown grassland 2, GR 3: Mown grassland 3, FF1: Secondary forest 1, FF2: Secondary forest 2, FF3: Secondary forest 3 |
Reference collection specimens were deposited in the Entomology Collections of Department of Zoology, Obafemi Awolowo University, Ile-Ife and the Entomology Museum of the Agricultural Research Council, Roodeplaat, Pretoria, South Africa. Insects collected were sorted under a dissecting microscope (Model-ZEISS Steimi, 2000) and identified with the assistance of expert taxonomists and with the use of the following keys, Bees of the world12, Common Butterflies of IITA13, Butterfly of West Africa14, Reference collection in Entomology Laboratory of Zoology Department and Museum of Natural History, Obafemi Awolowo University, Ile-Ife were also used.
Statistical analysis: Additive partitioning was used to determine the alpha and beta diversity of flower-visiting insects15,16, γ = α+β. This technique shows no sensitivity to differences in sampling effort among replicates and therefore does not require rarefaction of data prior to analyses16,17. The approach of Kehinde and Samways2 and Tylianakis et al.3 was followed in defining partitions of diversity for flower-visiting insects. Alpha diversity was defined as the mean species richness of flower-visiting insects per site. Beta diversity was defined as the mean of total species richness in a land use type over the entire sampling period minus the mean species richness per site of that land use type. Generalized linear model (GLM) was used for statistical analyses of the effect of land use on the alpha and beta diversity of flower-visiting insects. All statistical analyses were performed in R (version 3.3.3, R development core team)18. Data were expressed in the form of Mean±SE. Statistical significance was determined at 5% probability level.
RESULTS
At the end of the sampling, the secondary forest, agricultural land, the grazed grassland and the mown grassland had 51, 40, 57 and 54 species of flower-visiting insects, respectively (appendix 1 species list).
| Appendix 1: | Species list of flower-visiting insects sampled |
 | |
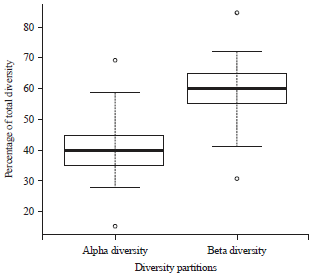 | |
| Fig. 2: | Percentage contribution of alpha and beta diversity to total diversity of flower-visiting plants |
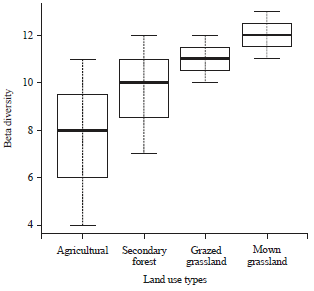 | |
| Fig. 3: | Mean (±SE) beta diversity in the different land use types sampled |
Alpha and beta partitions of diversity contributed meaningfully to the total diversity of flower-visiting insects with percentage contributions ranged from 15.4-69.2% for alpha diversity and 30.8-84.6% for beta diversity (Fig. 2). Beta diversity had a significantly higher percentage contribution to total diversity compare to alpha diversity (p = 0.004, t = -3.24, df = 22, Fig. 2).
Results of generalized linear model showed that alpha diversity was not significantly different between the land use types (p>0.05) however beta diversity showed marginal significant difference when compare among the land use types (p = 0.05, z = 1.93, df = 3). The grasslands had higher beta diversity of flower-visiting insects compared to secondary forest and agricultural land (Fig. 3).
DISCUSSION
This study reports meaningful contribution of alpha and beta diversity to the total diversity of flower-visiting insects across the land use types. Assessing the various partitions of diversity provides a holistic perspective in biodiversity surveys and assessments19. This ensures all important components of diversity at local, landscape and regional scale are accounted for in such surveys and the consequent conservation action or decision to be taken2. Beta diversity had the highest percentage contribution to total diversity showing a similar pattern with previous studies that have reported higher beta diversity compared to alpha diversity1,2,20. This implies that species turnover on the landscape scale forms a critical part of organismal biodiversity6. This is particularly so for highly mobile groups such as flower-visiting insects which disperse frequently when foraging for food and other resources. Moreover, this underscores the importance of beta diversity in assessing the spatial differentiation of species richness across the landscape which is crucial to determining important ecosystem indices such as biotic homogenization21.
Though alpha diversity of flower-visiting insects was similar among the land use types, beta diversity showed marginal significant difference, implying that even when land use effects is not detected on local scale diversity, effects might be found with the turnover in species diversity. For high dispersers such as flower-visiting insects, their foraging activity implies that their local diversity and species turnover is often subject to land use features and disturbances within their local habitat as well at the landscape level22.
Furthermore, grassland habitats especially mown grasslands had higher turnover in species of flower-visiting insects compared to the secondary forest and agricultural land. This response may be explained by the intermediate disturbance hypothesis23-25 which implies that disturbance in the grasslands is at a level that supports a diverse population of the flower-visiting insects. However, caution must be exercised to ensure that the level of disturbance does not exceed the threshold where the resources and habitat requirements of these insect groups that are available in the grasslands are affected adversely.
CONCLUSION
Species turnover of flower-visiting insects provides information on the sensitivity of these organisms to disturbance such as land use change. It is, therefore, important to include other partitions of diversity apart from the local scale (alpha) diversity in biodiversity assessments of flower-visiting insects to promote conservation decisions from holistic perspectives.
SIGNIFICANCE STATEMENT
This study discovers the important role of alpha and beta partitions of diversity in the biodiversity assessment of flower-visiting insects in different land use types. This study will help the researchers to uncover the critical area of using holistic approaches in biodiversity assessments that many researchers were not able to explore. Thus, a new theory on effects of anthropogenic activities on species turnover of flower-visiting insects and other organisms may be arrived at.
REFERENCES
- Clough, Y., A. Holzschuh, D. Gabriel, T. Purtauf and D. Kleijn et al., 2007. Alpha and beta diversity of arthropods and plants in organically and conventionally managed wheat fields. J. Applied Ecol., 44: 804-812.
CrossRefDirect Link - Kehinde, T. and M.J. Samways, 2014. Management defines species turnover of bees and flowering plants in vineyards. Agric. For. Entomol., 16: 95-101.
CrossRefDirect Link - Tylianakis, J.M., A.M. Klein and T. Tscharntke, 2005. Spatiotemporal variation in the diversity of Hymenoptera across a tropical habitat gradient. Ecology, 86: 3296-3302.
CrossRefDirect Link - Lande, R., 1996. Statistics and partitioning of species diversity and similarity among multiple communities. Oikos, 76: 5-13.
CrossRefDirect Link - Zanuncio, J.C., W.D.S. Tavares, B.V. Fernandes, C.F. Wilcken and T.V. Zanuncio, 2014. Production and use of Heteroptera predators for the biological control of Eucalyptus pests in Brazil. Ekoloji, 91: 98-104.
CrossRefDirect Link - Shaw, J.D., D. Spear, M. Greve and S.L. Chown, 2010. Taxonomic homogenization and differentiation across Southern Ocean Islands differ among insects and vascular plants. J. Biogeogr., 37: 217-228.
CrossRefDirect Link - Klein, A.M., B.E. Vaissiere, J.H. Cane, I. Steffan-Dewenter, S.A. Cunningham, C. Kremen and T. Tscharntke, 2007. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B: Biol. Sci., 274: 303-313.
CrossRefPubMedDirect Link - Tscharntke, T., A.M. Klein, A. Kruess, I. Steffan-Dewenter and C. Thies, 2005. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol. Lett., 8: 857-874.
CrossRefDirect Link - Adedoja, O. and T. Kehinde, 2017. Seasonal variation in the diversity of anthophilous insects in three land use types on Obafemi Awolowo University, Ile-Ife, Southwestern Nigeria. Environtropica, 14: 63-74.
Direct Link - Myers, N., R.A. Mittermeier, C.G. Mittermeier, G.A.B. da Fonseca and J. Kent, 2000. Biodiversity hotspots for conservation priorities. Nature, 403: 853-858.
CrossRefDirect Link - Wagner, H.H., O. Wildi and K.C. Ewald, 2000. Additive partitioning of plant species diversity in an agricultural mosaic landscape. Landscape Ecol., 15: 219-227.
CrossRefDirect Link - Crist, T.O., J.A. Veech, J.C. Gering and K.S. Summerville, 2003. Partitioning species diversity across landscapes and regions: A hierarchical analysis of α, β and γ diversity. Am. Nat., 162: 734-743.
CrossRefDirect Link - Kessler, M., S. Abrahamczyk, M. Bos, D. Buchori and D.D. Putra et al., 2009. Alpha and beta diversity of plants and animals along a tropical land-use gradient. Ecol. Applic., 19: 2142-2156.
CrossRefDirect Link - Roschewitz, I., D. Gabriel, T. Tscharntke and C. Thies, 2005. The effects of landscape complexity on arable weed species diversity in organic and conventional farming. J. Applied Ecol., 42: 873-882.
CrossRefDirect Link - Kehinde, T. and M.J. Samways, 2014. Effects of vineyard management on biotic homogenization of insect-flower interaction networks in the cape floristic region biodiversity hotspot. J. Insect Conserv., 18: 469-477.
CrossRefDirect Link - Kleijn, D. and F. van Langevelde, 2006. Interacting effects of landscape context and habitat quality on flower visiting insects in agricultural landscapes. Basic Applied Ecol., 7: 201-214.
CrossRefDirect Link - Mackey, R.L. and D.J. Currie, 2001. The diversity-disturbance relationship: Is it generally strong and peaked? Ecology, 82: 3479-3492.
CrossRefDirect Link - Bruggisser, O.T., M.H. Schmidt-Entling and S. Bacher, 2010. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv., 143: 1521-1528.
CrossRefDirect Link - Townsend, C.R., M.R. Scarsbook and S. Doledec, 1997. The intermediate disturbance hypothesis, refugia and biodiversity in streams. Limnol. Oceanogr., 42: 938-949.
CrossRefDirect Link