
Sulaeha Th
Faculty of Agriculture, Hasanuddin University, 92661 Makassar, South of Sulawesi, Indonesia
LiveDNA: 62.19986
Aunu Rauf
Faculty of Agriculture, Bogor Agricultural University, 16680 Bogor, West Java, Indonesia
Purwantiningsih
Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, 16680 Bogor, West Java, Indonesia
Endang Sri Ratna
Faculty of Agriculture, Bogor Agricultural University, 16680 Bogor, West Java, Indonesia
Journal of Entomology
Year: 2017 | Volume: 14 | Issue: 5 | Page No.: 216-227







ABSTRACT
Background and Objective: Melon fly, Zeugodacus cucurbitae (Coquillett) (Diptera:Tephritidae), is a major agricultural pest in Indonesia. It is considered a specialist insect since it only attacks plants that belong to the family Cucurbitaceae. Such insects commonly use plant semiochemicals to find appropriate hosts for feeding and oviposition. The objective of this study was to identify chemical compounds from the host plants that were attractive to male and female Zeugodacus cucurbitae (Z. cucurbitae). Materials and Methods: Chemical blends were extracted from the host plants and responses of the melon fly to the scents were studied by Y-olfactometer. Results: Fractions of bitter gourd (Momordica charantia) were more attractive to male and female Z. cucurbitae compared to those of angled luffa and cucumbers. Compounds extracted from the leaves of bitter gourd after saponification were identified as a ketone (6,10,14-trimethyl-2-pentadecanon), an aldehyde (3,7,11,15-tetramethyl-2-hexadecen-1-ol) and phytosterol. Conclusion: The ketones and the aldehydes are kairomones for Z. cucurbitae.
PDF Abstract XML References Citation
Received: September 07, 2017;
Accepted: October 30, 2017;
Published: November 11, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sulaeha Th, Aunu Rauf, Purwantiningsih and Endang Sri Ratna, 2017. Identification of Kairomonal Compounds From Host Plants Attractive to Melon Fly, Zeugodacus cucurbitae (Coquillett) (Diptera:Tephritidae). Journal of Entomology, 14: 216-227.
DOI: 10.3923/je.2017.216.227
URL: https://scialert.net/abstract/?doi=je.2017.216.227
DOI: 10.3923/je.2017.216.227
URL: https://scialert.net/abstract/?doi=je.2017.216.227
INTRODUCTION
Melon fly, Zeugodacus cucurbitae (Coquillett) (Diptera:Tephritidae), is a serious pest of cucurbit plants. Yield losses caused by Z. cucurbitae attack could reach 30-100%1,2. Control of fruit flies using insecticides is generally neither effective nor efficient because the larvae of melon fly live inside the fruit tissues and the adults actively fly. Fruit fly controls that are commonly used in some countries include the sterile insect technique3 and sex pheromone traps4. The weaknesses of the former method are that it can only be conducted by certain agencies and it is relatively expensive. The latter one can only be applied to the male fruit flies and it does not attract the female insects that oviposit on host plants.
In Indonesia, controlling and monitoring the melon fly population has used an analogue of the sex pheromone compound 4-(p-acetoxyphenyl)-2-butanone in its synthetic form (cue-lure)5. Thus far, Indonesia’s Agricultural Quarantine is still importing this compound from overseas but chemical compounds similar to cue-lure have recently been found. They include raspberry ketone (RK) from orchid plants (Dendrobium superbum)6 and zingerone compounds (Z) from wild orchid [Bulbophyllum paten (Kings)]7. These compounds act similarly to sex pheromone for Z. cucurbitae, which can only attract the male melon fly8.
The mechanism of host plant selection by melon fly is guided by volatile compounds produced by plants. Therefore, it is important to identify attractant compounds for male and female Z. cucurbitae. According to Nishida8, volatile compounds produced by plants act as infochemicals for insects in the process host selection process. Bernays and Chapman9, reported that plant odors are important environmental cues for insects to locate their hosts. For example, Bactrocera invadens can detect its host plant, olive trees (Olea europaea), through the volatile compounds that they produce. These compounds are also attractants for Bactrocera oleae and stimulants for its oviposition10. Bactrocera invadens is also attracted by mango and almond fruits because of their production of 4-methyl-3-penten-2-one. This compound also occurs in the wild plant Gynandropsis gynandra and it has been used as a kairomone for B. invadens control11.
Fieldwork results have shown that fruit flies ingesting cucurbit plants in Bogor are predominately Z. cucurbitae. This finding highlighted the attractiveness of Cucurbitaceae plants to Z. cucurbitae. According to Sapkota et al.12, Z. cucurbitae has a very strong host association with cucurbit plants such as melon, kabocha, angled luffa [Luffa acutangula (L.) Roxb.] and cucumber (Cucumis sativus L.). The focus on certain hosts is influenced by the existence of specific chemical compounds in the plant13 and the specificity can be exploited for insect control. For example, Hawaiian farmers have used cucumber slices in traps for female melon flies14. Furthermore, volatile compounds contained in the host plants strongly influence fruit fly behaviors and activities, including the host finding15,16, foraging17, mating and oviposition8,10,18,19.
Identification of the kairomonal compounds derived from host plants that attract both male and female Z. cucurbitae has never been reported. Currently, much research focuses on identifying plant derivatives that function as sex pheromones for melon flies but they are only attractive for male melon flies, not females. Therefore, this study focuses on identifying attractant compounds for male and female Z. cucurbitae. These compounds will have a crucial role for determining kairomonal compounds that are attractive for Z. cucurbitae control. The identification was aimed at finding and identifying compounds from cucurbit plants that attract Z. cucurbitae.
MATERIALS AND METHODS
Time and locations: Samples used in this identification were leaves and fruits of three plant species collected from several locations in Bogor, West Java, Indonesia. The leaves and fruits of bitter gourd (Momordica charantia) were collected from Jabon Mekar village, Parung District (6°26 21.7"S, 106°42 57.4"E), while the leaves and fruits of angled luffa were collected from Situ Udik village, Cibungbulang District (6°26 21.7"S, 106°42 57.4"E). The leaves and fruits of cucumber were collected from Sukawening village, Darmaga District (6°36 24.0"S, 106°44 22.8"E), Bogor Regency, Indonesia. Olfactometer bioassays and the identification of chemical compounds were done between September, 2015 and July, 2016 at the Laboratory of Insect Ecology, Department of Plant Protection and the Laboratory of Organic Chemistry, Department of Chemistry, Bogor Agricultural University (IPB). The gas chromatography-mass spectrometry (GC-MS) analysis was done at the Laboratory of Forensics in the National Police Headquarters.
Rearing of Z. cucurbitae: Melon flies were collected from the fields by collecting infested fruits of angled luffa, bitter gourd and cucumber. The infested fruits were placed in plastic containers (12 cm diameter and 20 cm high) that were filled with fine, sterile sawdust. After 7 days, the samples were observed to determine if the insect larvae had transformed into pupae. Pupae were then collected in small cages that were placed in a wood cage (50×40×30 cm). Emerging adults were fed with a mixture of hydrolyzed protein and fine sugar (1:4). Once adults reached 10 days of age, an egging device was placed in the cages. Eggs were subsequently harvested and then spread on the artificial diet in box plastic 20×10×5 cm and developed in the Laboratory of Insect Ecology in IPB. Insects from the third laboratory raised generation were used in experiments.
Preparations of sample: Samples of leaves collected from angled luffa, bitter gourd and cucumber were cleaned using tissue wipes and then dried in the oven at 40°C temperature for 7 days. The samples were mechanically cut with blade until their pieces were 60 mesh in size. The minced sample was then macerated in methanol solution (purities were >98%) at 1:10 ratio. During maceration, the solution was stirred several times. The filtering process was done in every 3 days. Samples were macerated in methanol until the filtering process was done three times. The resulting filtrate was evaporated by using a rotary evaporator until all solvent was evaporated, leaving a paste. The resulting paste, referred to as the crude methanol extract, was then weighed. All chemicals used in the present study were obtained from Merck®, including the standards available and used for the chromatographic determinations and behavioral bioassays.
Isolation of compounds: The process of compound fractionation was done by dissolving the crude methanol extracts in n-hexane for analysis purities were >99% at a 1:3 ratio. This process produced filtrate and a residue. The filtrate was evaporated with a rotary evaporator, eventually forming a paste once the solvent evaporated. This paste is referred to as the n-hexane fraction. The residue underwent a process of maceration in acetone solvent for analysis purities were >99%, which produced another residue and filtrate. The resulting filtrate from this process is the acetone fraction.
The identification of terpenoid compounds was done through hydrolysis and saponification of the n-hexane fraction of bitter gourd. Ten grams of this fraction was dissolved in 25 mL of ethanol and 9.5 g of KOH. After the sample was entirely dissolved, it was refluxed for 1 h and subsequently cooled for approximately 1 h. It was then saponified by adding 100 mL of diethyl ether and 50 mL of H2O to eliminate the fats in the sample. Next, the solution was agitated until the separation of the saline solution (saponified fraction) and the unsaponified fraction was observed. The process of separation was done continuously until separation ceased to occur. The identification for compound contained in the sample was done through GC-MS analysis.
GC-MS analysis was performed on an Agilent 6890N GC interfaced with a Hewlett-Packard 5973 mass selective detector. The GC was equipped with an HP-5MS column (30 m×250 μm×0.25 μm ID 0.25 μm film thickness), which was temperature programmed from 10-300°C min‾1 following a 1 min delay. The injector temperature was 100°C and helium was the carrier gas (1.1 mL min‾1). Compounds cucurbits were identified on the basis of their retention times and mass spectra (mass spectral database chemstation E.02.02.1431) and in comparison with authentic standards.
Y-tube olfactometer: The examination of Z. cucurbitae attractiveness used Y-olfactometer tubes as described by Finidori-Logli et al.20, with modifications. The air source was provided by a 115-V, 1.5-A air pump. The air flowed through a charcoal filter tube (5 cm diameter, 21 cm height) connected to a hose leading into the flow meter. The flow rate into the odor source tube was 12-15 m sec‾1. This tube was connected to the olfactometer Y tube, which had a 32 cm long branch, 10.5 cm-long base and 75°C branching angle. The circuit of the Y-olfactometer tube was placed in a cage (50 cm height), the bottom surface of the cage was white and the cage was covered by a dark cloth. The upper part of the cage was illuminated by a 75 W white fluorescent lamp.
Olfactometer bioassay: Responses of Z. cucurbitae to volatiles compounds emitted from extracts of cucurbit leaves and fruits were observed. This test monitored the behavior of flies in terms of their attraction to volatile compounds from the crude methanol extract and n-hexane fraction as well as extracts from the leaves and fruits. Attraction to volatile compounds of the acetone and n-hexane (candidate) fractions and to isolated terpenoid compounds was assessed. This examination was done in an olfactometer in which a 0.1 mL drop of extract was placed by syringe on a 4×3 cm, No. 42 Whatman filter paper and allowed to dry for approximately 1 min. The paper with the sample was inserted into a chamber separate from other examined samples and pure methanol was used as the control sample. Experimental insects were released into the olfactometer and then into the Y-tube (one individual per observation). The experiment was replicated 25 times.
Experimental insects were fasted for 1 h prior to the experiment. The examination was divided into three age categories, including the newly emerged male and female, imago aged 1-2 days (had not been copulated) and copulated female imago (pre-oviposition period) and aged 30 days (post-oviposition period). Prior to each set of bioassays, adults were allowed to acclimatize for 1 h in the new cage. Adults had no exposure to host plant chemicals before the bioassays. Experimental insects were given 1 h to choose their direction. These insects were considered to have made a choice once they headed into one arm of the Y-tube (i.e., either the control or the experimental sample). If an insect remained at the base (stem) of the Y-tube, their response was labeled "undetermined", following Addesso and McAuslane21. Individual flies were used only once. For each experimental insect, the odor chamber was also replaced with a new sterilized Y-tube. Sterilization was done by cleaning the chamber tube with sterile water, rinsing it with methanol and acetone and allowing it to air-dry. The Y-tube was activated for 1 h before the olfactometer was used. The parameters for behavioral observation were the duration of Z. cucurbitae visits on each extract and the response of both male and female insects to volatile compounds from the crude methanol extract, n-hexane fraction and the isolated terpenoid compounds.
Statistical analysis: All statistical models included the effect of the odor source. Data from bioassays carried out in the Y-tube olfactometer were analyzed using the independent-sample t-test. Chi-square (χ2) analysis was used to compare the observed attraction for the percentage of flies in the Y-tube arm with the expected value (α = 0.05).
RESULTS
Response of Z. cucurbitae to methanol crude extract of cucurbit plants leaves and fruits: On average, the time spent by Z. cucurbitae on the methanol solvent (control) was higher than on the methanol extracts (treatment). The results of the independent sample t-test showed that both newly emerged and mated Z. cucurbitae were significantly more active when exposed to the control as compared to the plant odor. Attractiveness on males and females Z. cucurbitae showed a significant response to control compared to the odor source of angled luffa, bitter gourd and cucumber for leaves and fruit (t = 2.472, -2.411, -3.738, 4.481, t = 2.244, p<0.05) but response on control and leaves angle luffa were showed not significant response by Z. cucurbitae (t = 1.875, p = 0.062). The average time used for volatile organic compounds the crude extract of methanol is less than 20 min, while the time taken for methanol solvent is more than 20 min (Table 1).
Response of Z. cucurbitae to n-hexane fraction of cucurbit plants leaves and fruits: The results from the experiments with the n-hexane fraction derived from the leaves and fruits of cucurbit plants showed significant differences among plant species in terms of volatile compounds that acted as attractants for Z. cucurbitae ((sig.) = 0.000<0.05). Observation of 300 melon flies showed that angled luffa and bitter gourd leaves were more attractive to Z. cucurbitae. On average, flies visiting the odor chamber containing the n-hexane fractions of leaves from bitter gourd and angled luffa remained for 28 min, 10 sec (t = 4.613, p = .00) and 27 min, 23 sec (t = 1.971, p = 0.05), respectively, while time spent on the fruit extract samples was less than 21 min. The shortest visit time was observed for the cucumber fraction, which was only 10 min, 49 sec (t = 3.603, p = 0.00) (Table 2).
| Table 1: | Length of time spent on each given odor source by Z. cucurbitae on crude methanol extracts of cucurbit plant leaves and fruits |
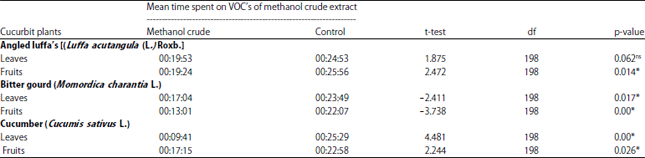 | |
| *Mean time spent in each arm of the Y-tube olfactometer was significant in t-tests (p<0.05), ns: Not significant, VOC: Volatile organic compounds | |
| Table 2: | Length of time on each given odor source by Z. cucurbitae on the n-hexane fraction of cucurbit plant leaves and fruits |
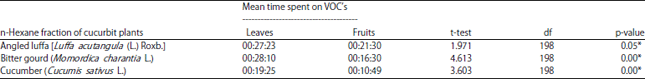 | |
| *Mean time spent in each arm of the Y-tube olfactometer was significant in t-tests (p<0.05), VOC: Volatile organic compounds | |
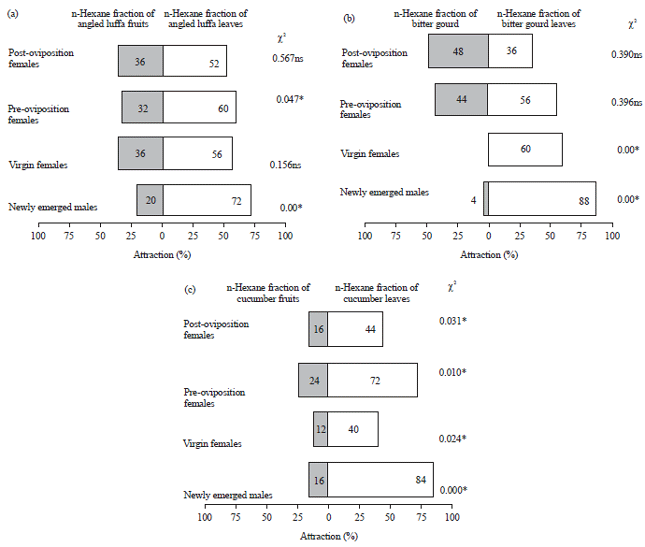 | |
| Fig. 1(a-c): | Responses of males and females Z. cucurbitae in an olfactometer to the n-hexane fraction of three kinds of cucurbit plants, (a) Angled luffa, (b) Bitter gourd and (c) Cucumber *p<0.05, ns: Not significant |
The highest percentage (88%) of melon flies attracted to fractions extracted from cucurbit plants was found on the leaves of bitter gourd (χ2 = 0.00, p<0.05). Overall, the highest percentages of attraction (mostly over 50%) were found for leaves of all plant species for all ages. The exceptions were post-oviposition females on bitter gourd (36%) and virgin females and post-oviposition females on cucumber (40%, χ2 = 0.024 and 44%, χ2 = 0.031, respectively). The percentage of attraction to the leaves and fruits of cucumber showed significantly different responses for all fruit flies (male and female) with χ2 = 0.00, 0.024, 0.01 and 0.031 (p<0.05) for newly emerged males and females, virgin females and pre-oviposition and post-oviposition females (Fig. 1).
The attraction response of Z. cucurbitae flies at various ages (newly emerged males, virgin females, pre-oviposition females) showed that visits to the leaves were longer (over 30 min) than to the fruits (under 25 min). The average number of visits was one to three times. The n-hexane fraction of bitter gourd leaves appeared to be the most attractive, with virgin females spending an average 36 min, 23 sec on it. The average duration of male fruit fly visits on the extracts from angled luffa leaves was 31 min, 24 sec, which was higher than the time spent on bitter gourd (23 min, 7 sec). However, the percentage of males interested in the bitter gourd leaves reached 88%, which was higher than the interest in angled luffa leaves, which was 72% (Table 3 and Fig. 1).
GC-MS analysis on the n-hexane fraction of bitter gourd leaves revealed the presence of Z-13-octadecenal, farnesyl acetone, hexadecanoic acid and methyl-1inolenate. The compounds in the n-hexane fraction included 6 alcohols, 4 aldehydes, 3 ketones and 15 esters.
| Table 3: | Length of time on each given odor source by Z. cucurbitae on the n-hexane fraction of the leaves and fruits of cucurbit plants |
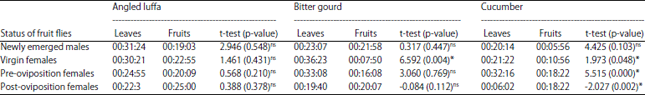 | |
| *Mean time spent in each arm of the Y-tube olfactometer was significant in t-test (p<0.05), ns: Not significant | |
Several of these compounds were found in higher concentrations in the leaves compared with the fruits (Table 4).
Response of Z. cucurbitae to n-hexane fraction and acetone fraction of bitter gourd’s leaves: The duration Z. cucurbitae visits differed significantly between n-hexane and acetone fractions of bitter gourd leaves (p = 0.000 and 0.027, respectively). Our observations showed that the average time on the n-hexane fraction was higher than on the acetone fraction in all age categories, except for the post-oviposition females (Table 5). This result indicated that bitter gourd leaf extracts using polar and nonpolar solvents could be affected by the specific volatile compounds from the plant. The volatile compounds extracted with the nonpolar solvent n-hexane yielded the best results.
Response of Z. cucurbitae to the n-hexane fraction and terpenoid compound isolated from bitter gourd leaves: Experimental results from the n-hexane fraction and the isolated terpenoid compounds from bitter gourd leaves showed significantly different responses between the age categories of Z. cucurbitae (p = 0.020). These results proved that the insects’ age affected the duration of their visits. The longest visit duration for newly emerged Z. cucurbitae (male and female) occurred with the n-hexane fraction. For post-copulation fruit flies, both in the pre-oviposition and post-oviposition periods, the longest visit duration was associated with isolated terpenoids
The high percentage and long visit duration of Z. cucurbitae males and females in the virgin category (newly emerged) on the n-hexane fraction was likely due to the chemical contents of the extract. The independent sample t-test showed both newly emerged Z. cucurbitae (male and female) were significantly more active when exposed to the n-hexane fraction of bitter gourd leaves (t = 6.168, p = 0.000; t = 3.423, p = 0.001). While the longest visit duration of post-copulation fruit flies, in both the pre-oviposition and post-oviposition periods, was found in the compound resulting from the isolation of terpenoid. Mean time spent on the isolated terpenoid was 30 min, 36 sec and 24 min, 15 sec for the pre-oviposition and post-oviposition periods, respectively, while time spent on the n-hexane fraction was less than 12 min for both. The independent sample t-test showed melon flies in both pre-oviposition and post-oviposition periods were significantly more active when exposed to isolated terpenoid than the n-hexane fraction (t = 6.168, p = 0.000; t = 3.423, p = 0.001) (Table 6).
The highest percentage (60%) and longest visit duration (30 min, 19 sec) of post-copulation Z. cucurbitae (pre-oviposition and post-oviposition) occurred for terpenoid compounds (Fig. 2). These compounds serve as stimulants for fruit flies in pre-oviposition and post-oviposition phases seek host plants on which to lay eggs, which was observed with the behavioral response of fruit flies responding to the scent of the compounds.
Behavioral responses shown for both odor sources (n-hexane fraction and terpenoid compounds) indicated that Z. cucurbitae of all age categories were more active in the chamber with terpenoid compounds. The flies fluttered their wings very rapidly and continuously. They also rubbed their antennae with anterior limbs and with posterior legs on their abdomens very rapidly and repeatedly. It was also observed that the ovipositor of post-copulation females (pre-oviposition and post-oviposition) was extended.
Mixed compounds may provoke different behavioral responses compared to a single compound. Therefore, behavioral activities will depend on the type of components in the mixture that can be detected by the olfactory system. It was concluded that the isolated terpenoids are short distance stimulants for Z. cucurbitae to locate their host plants. For that reason, the level of perception of odor quality will depend on the combination of volatile compounds that can be detected by insects. These responses will vary among insects based on their ages and species.
Compounds identified after saponification and terpenoid isolation were 6,10,14-trimethyl 2-pentadecanone (ketone) and 3,7,11,15-tetramethyl-2-hexadecen-1-ol (aldehyde), common phytol compounds and another compounds from the sterol group (terpenoid) including δ-7,25-stigmasta-dien-ol and o-xylotocopherol (Table 7).
| Table 4: | GC-MS analysis of chemical compounds from bitter gourd leaves in the n-hexane fraction |
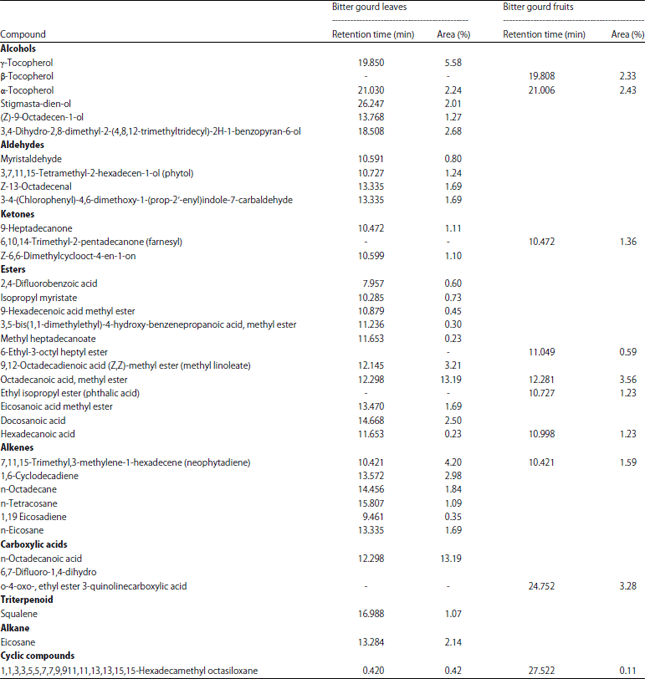 | |
| Table 5: | Length of time on each given odor source by Z. cucurbitae on n-hexane and acetone fractions of bitter gourd leaves by fruit fly status |
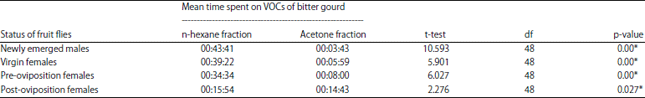 | |
| *Mean time spent in each arm of the Y-tube olfactometer was significant in t-tests (p<0.05), VOC: Volatile organic compounds | |
| Table 6: | Length of time on each given odor source by Z. cucurbitae on n-hexane fraction and terpenoid isolation of bitter gourd’s leaves in status of fruit flies |
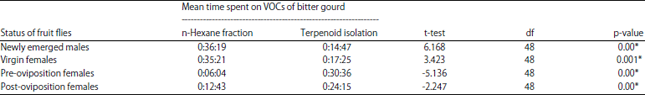 | |
| *Mean time spent in each arm of the Y-tube olfactometer was significant in t-tests (p<0.05), VOC: Volatile organic compounds | |
| Table 7: | Chemical composition of terpenoid compounds from bitter gourd leaves |
 | |
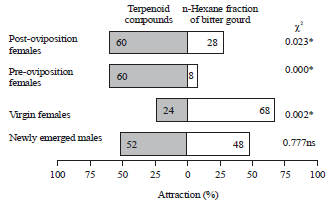 | |
| Fig. 2: | Responses of males and females Z. cucurbitae in an olfactometer to the n-hexane fraction and isolated terpenoid compounds from bitter gourd leaves *p<0.05, ns: Not significant |
The concentration of the aldehyde compound 3,7,11,15-tetramethyl-2-hexadecen-1-ol was 30 times higher than the ketone compound 6,10,14-trimethyl 2-pentadecanone (farnesyl acetone) in 1 μL of the experimental sample (Fig. 3).
DISCUSSION
Attraction to whole methanol crude extract of cucurbit plants leaves and fruits: Mean time spent by Z. cucurbitae on volatile organic compounds of the methanol crude extract of volatile compounds was less than the time spent on the solvent. There was no difference found in the responses of the three species of cucurbits due to the use of solvent used in extracting volatile compounds. The solvent used was methanol, which is a polar compound and contains an alcohol functional group. The use of polar solvent can limit extraction of nonpolar volatile compounds from plants. Harborne22, reported that volatile compounds generally belong to the terpenoid group. In order to extract these kinds of compounds from plant tissues, solvents with a low level of polarity are needed. One such solvent is n-hexane. Therefore, the subsequent identification of volatile compounds was done by using n-hexane as the fractionating solvent.
Attraction to n-hexane fraction of cucurbit plants leaves and fruits: The attraction of Z. cucurbitae to the odor chamber that contained n-hexane fractions of bitter gourd leaves and angled luffa led to longer visit times compared with the fruit extract samples. These results are in contrast to those of Siderhurst and Jang23, who used gas chromatography electroantenogram (GC-EAG) detection. They reported that a slice of cucumber contained volatile compounds that were more attractive to males and females of Z. cucurbitae than volatile compounds from alcohol and aldehyde groups.
The phytochemical contents in the leaves of cucurbit plants were alkaloid, saponin, flavonoid and steroid/terpenoid. These primary and secondary metabolite compounds affected the fruit flies visits. Our experiments showed that male and female flies (aged 1-2 days and pre-oviposition) visited leaves more frequently than fruits. These findings were most likely caused by higher concentration of phytochemicals such as aldehydes and steroids in the leaves. In nature, newly emerged flies would prioritize identifying nectars and plant exudates that would fulfill their nutritional needs rather than finding host fruits for their offspring24,25. According to Allwood26, newly emerged insects identify their nutritional sources based on proteins and nitrogen containing compounds, such as amino acids, alkaloids and aldehydes, that are needed for development and reaching sexual maturity27. A different pattern of attraction was observed in insects that were already sexually mature. They appeared to be more sensitive to the easily evaporated organic compounds [volatile organic compounds (VOC’s)] released by host plants. Several studies using the GC-EAG method have shown that certain volatile compounds attract fruit flies. These compounds included (+/-)-1-octen-3-ol, (Z)-6-nonenal, Z-13-octadecenal, farnesyl acetone, pentadecanone, linalool, (E)-2-hexanal, acetic acid, hexadecanoic acid and methyl-1inolenate.
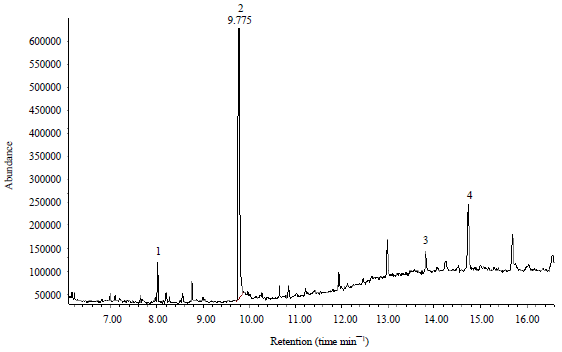 | |
| Fig. 3: | GC-MS analysis bitter gourd leaves in the isolation of volatile compound, 1: 6,10,14-Trimethyl 2-pentadecanone, 2: 3,7,11,15-Tetramethyl-, [R-[R*,R*-(E)]] 2-hexadecen-1-ol, 3: 8-methyltoco, 4: O-xylotocopherol |
They are known to attract various species of fruit flies, including Anastrepha ludens28, Dacus dorsalis29, Ceratitis capitata30,31, Rhagoletis pamonella32, B. invadens33 and B. oleae34. The compounds are categorized as green leaf volatiles and they consist of numerous saturated or mono unsaturated aldehydes, alcohols and acetate compounds that occur in all plants but in varying proportions depending on species35.
GC-MS analysis on the n-hexane fraction of bitter gourd leaves revealed the presence of Z-13-octadecenal, farnesyl acetone, hexadecanoic acid and methyl-1inolenate. The compounds in the n-hexane fraction included 6 alcohols, 4 aldehydes, 3 ketones and 15 esters. Several of these compounds were found in higher concentrations in the leaves compared with the fruits. According to Light and Jang29, aldehydes have a stronger role than alcohols as olfactory cues for D. dorsalis fruit flies. In addition, experiments complemented with EAG showed that ketones attracted the fruit fly Bactrocera tryoni (Foggatt)36.
Attraction to terpenoid compound and plant volatile blends in n-hexane fraction by Z. cucurbitae on bitter gourd’s leaves: Experimental results from the n-hexane fraction of bitter gourd leaves and isolated terpenoid compounds from bitter gourd leaves showed that there was a significantly different attractive response between the age categories of melon flies. Attraction of Z. cucurbitae to VOCs was highest for complex chemical compounds compared with a terpenoid compounds for the newly emerged males and females. For age status of Z. cucurbitae, flies in the post-copulation period were found to have the greatest attraction to terpenoid compounds. This study indicated that the age category of insects will affect the capture response to volatile compounds produced by plants. It also found synergy among alcohols, aldehydes, ketones and esters and the compounds may have formed a mixture providing olfactory cues for the newly emerged Z. cucurbitae. Newly emerged fruit flies are more active in response to complex volatile compounds, presumably for finding host plants for feeding. Blends of plant volatile compounds were also studied by Aglamalaria et al.37, who found that a six component blend of acetates elicited stronger attraction than simpler blends for Ethiopian fruit flies Dacus ciliates on Cucumis melo. A separate study on behavioral responses shower stronger attraction to host plant volatile blends than simpler blends in Mexican fruit fly, Anastrepha ludens (Diptera:Tephritidae), on Sargentia greggii plant38.
The percentage and visit duration of post-copulation Z. cucurbitae (pre-oviposition and post-oviposition) were the highest in terpenoid chamber of the Y-tube olfactometer. Adult fruit flies that have copulated are interested in certain compounds that orient them to locations for laying eggs as well as volatile signals that lead them to plants that meet their nutritional needs. According to Bruce and Pickett39, the physiological condition of the insects will be able to influence the signal retrieval response to volatile compounds.
Further identification of terpenoids revealed that two compounds were 6,10,14-trimethyl 2-pentadecanone (ketone) and 3,7,11,15-tetramethyl-2-hexadecen-1-ol (aldehyde). The presence of ketones and aldehydes in plants could act as olfactory cues for fruit flies (both in long and short distance)29. Aldehydes are constituents of the general green-leaf odor that emanates from most plants. It was shown to have a strong effect on host foraging behavior of Mediterranean fruit fly C. capitata40. However, aldehydes can deter female Sesamia nonagrioides (Lepidoptera: Noctuidae)41 and Liriomyza trifolii (Diptera: Agromyzidae)42 from ovipositing. The ketone is also known as a sex pheromone produced by male Tirathaba mundella Walker (Lepidoptera:Pyralidae) to attract females. Following extraction of the males’ hair-pencils, wings, or entire body in n-hexane, GC-EAD analysis of hair-pencil extracts revealed four compounds that consistently elicited responses from female antennae. One of these compounds was 6,10,14-trimethyl 2-pentadecanone43. Therefore, behavioral activities will depend on the type of components in a mixture of compounds that can be detected by the olfactory system. It was concluded that the terpenoid compounds resulting from the isolation are short-distance stimulants for Z. cucurbitae to locate their host. These compounds are specific odors for Z. cucurbitae in cucurbits that act as infochemicals in insect and plant interaction.
Plant volatile compounds provide a crucial code for insects to find their host plants when foraging and ovipositing and also act as deterrents to unsuitable host plants17. The olfactory system of fruit flies can sense volatile chemicals at minuscule amounts, which permits them to detect a compound although the odor concentration is low. Single as well as mixed compounds produced by plants can prompt different responses from individual insects38. Therefore, behavioral activities will depend on the type of components mixed in the compound which can be detected by the olfactory nervous system.
Use of plant volatiles for host location by insects very much depends on their excellent ability to process olfactory signals. The insect olfactory system has extremely high sensitivity and specificity for certain volatiles44. For that reason, the level of perception on odor quality will depend on the combination of volatile compounds which can be detected by insects. These responses will vary among insects based on their ages and species.
CONCLUSION
In conclusion, melon flies (Z. cucurbitae) did not differ in their attractive response to crude methanol extracts of the leaves and fruits of bitter gourd, cucumber and angled luffa. Fruit flies were the most attracted to the n-hexane fraction of bitter gourd leaves. The leaves were more attractive than the fruits. Virgin female fruit flies hat had not laid eggs were more attracted to the n-hexane fraction of bitter gourd leaves compared with flies entering the oviposition stage.
The GC-MC analysis of bitter gourd plants identified alcohols, aldehydes, ketones and esters that were found in higher amounts in the leaves compared with the fruits. The terpenoid compounds 3,7,11,15-tetramethyl-2-hexadecen-1-ol and 6,10,14-trimethyl 2-pentadecanone that resulted from the isolation are a short distance stimulant used by Z. cucurbitae to locate their host. This compounds were specific odor for Z. cucurbitae in cucurbits plant as infochemical in insect and plant interaction.
SIGNIFICANCE STATEMENTS
This study discovered two terpenoid compounds isolated from n-hexane fraction of bitter gourd leaves, i.e. 6.10.14-trimethyl 2-pentadecanone (ketone) and 3.7.11.15-tetramethyl-2-hexadecen-1-ol (aldehyde). These compounds were attractive for male and female Z. cucurbitae and thus providing a novelty regarding. The compounds serve as a connection between Z. cucurbitae and its main host plant. This study serve as the basis for subsequent exploration of similar compounds in other plants for the purpose of developing control tactics for Z. cucurbitae.
ACKNOWLEDGMENTS
We thank the Ministry of Research Technology and Higher Education of Indonesia for sponsorship that enabled this research (grant number 079/SP2H/LT/DRPM/II/2016). We would also like to thank to Andi Fitriany and Febria Elvy Susanty in the Department of Chemistry, Bogor Agricultural University, for their assistance in classifying chemical compounds based on its functional groups.
REFERENCES
- Haldhar, S.M., D.K. Samadia, R. Bhargava and D. Singh, 2017. Host plant genotypes determine bottom-up effect of Cucumis melo var. callosus against melon fruit fly. Crop Prot., 98: 157-165.
CrossRefDirect Link - Dhillon, M.K., R. Singh, J.S. Naresh and H.C. Sharma, 2005. The melon fruit fly, Bactrocera cucurbitae: A review of its biology and management. J. Insect Sci., Vol. 5.
Direct Link - Keng-Hong, T. and R. Nishida, 2005. Synomone or kairomone?-Bulbophyllum apertum flower releases raspberry ketone to attract Bactrocera fruit flies. J. Chem. Ecol., 31: 497-507.
CrossRefDirect Link - Khoo, C.C.H. and K.H. Tan, 2000. Attraction of both sexes of melon fly, Bactrocera cucurbitae to conspecific males-a comparison after pharmacophagy of cue-lure and a new attractant-zingerone. Entomol. Exp. Applic., 97: 317-320.
CrossRefDirect Link - Nishida, R., 2014. Chemical ecology of insect-plant interactions: Ecological significance of plant secondary metabolites. Biosci. Biotechnol. Biochem., 78: 1-13.
CrossRefDirect Link - Kimbokota, F., P.G.N. Njagi, B. Torto, S. Ekesi and A. Hassanali, 2013. Responses of Bactrocera invadens (Diptera: Tephritidae) to volatile emissions of fruits from three hosts. J. Biol. Agric. Health, 3: 53-60.
Direct Link - Sapkota, R., K.C. Dahal and R.B. Thapa, 2010. Damage assessment and management of cucurbit fruit flies in spring-summer squash. J. Entomol. Nematol., 2: 7-12.
Direct Link - Prokopy, R.J., N.W. Miller, J.C. Pinero, L. Oride, N. Chaney, H. Revis and R.I. Vargas, 2004. How effective is GF-120 fruit fly bait spray applied to border area sorghum plants for control of melon flies (Diptera: Tephritidae)? Florida Entomol., 87: 354-360.
CrossRefDirect Link - Aluja, M. and R.J. Prokopy, 1993. Host odor and visual stimulus interaction during intratree host finding behavior of Rhagoletis pomonella flies. J. Chem. Ecol., 19: 2671-2696.
CrossRefDirect Link - Brevault, T. and S. Quilici, 2010. Flower and fruit volatiles assist host-plant location in the Tomato fruit fly Neoceratitis cyanescens. Physiol. Entomol., 35: 9-18.
CrossRefDirect Link - Gerofotis, C.D., C.S. Ioannou, C.T. Nakas and N.T. Papadopoulos, 2016. The odor of a plant metabolite affects life history traits in dietary restricted adult olive flies. Sci. Rep., Vol. 6.
CrossRef - Papadopoulos, N.T., N.A. Kouloussis and B.I. Katsoyannos, 2006. Effect of plant chemicals on the behavior of the mediterranean fruit fly. Proceedings of the 7th International Symposium on Fruit Flies of Economic Importance: From Basic to Applied Knowledge, September 10-15 2006, Salvador, Brazil, pp: 97-106.
- Finidori-Logli, V., A.G. Bagneres and J.L. Clement, 1996. Role of plant volatiles in the search for a host by parasitoid Diglyphus isaea (Hymenoptera: Eulophidae). J. Chem. Ecol., 22: 541-558.
CrossRefDirect Link - Addesso, K.M. and H.J. McAuslane, 2009. Pepper weevil attraction to volatiles from host and nonhost plants. Environ. Entomol., 38: 216-224.
CrossRefDirect Link - Siderhurst, M.S. and E.B. Jang, 2010. Cucumber volatile blend attractive to female melon fly, Bactrocera cucurbitae (Coquillett). J. Chem. Ecol., 36: 699-708.
CrossRefDirect Link - Sarles, L., A. Verhaeghe, F. Francis and F.J. Verheggen, 2015. Semiochemicals of Rhagoletis fruit flies: Potential for integrated pest management. Crop Prot., 78: 114-118.
CrossRefDirect Link - Liburd, O.E., 2004. Identification of host volatile compounds for monitoring blueberry maggot fly. Small Fruits Rev., 3: 307-312.
CrossRefDirect Link - Malo, E.A., L. Cruz-Lopez, J. Toledo, A. del Mazo, A. Virgen and J.C. Rojas, 2005. Behavioral and electrophysiological responses of the Mexican fruit fly (Diptera: Tephritidae) to guava volatiles. Florida Entomol., 88: 364-371.
CrossRefDirect Link - Light, D.M. and E.B. Jang, 1987. Electroantennogram responses of the oriental fruit fly, Dacus dorsalis, to a spectrum of alcohol and aldehyde plant volatiles. Entomol. Exp. Applic., 45: 55-64.
CrossRefDirect Link - Light, D.M., E.B. Jang and R.A. Flath, 1992. Electroantennogram responses of the Mediterranean fruit fly, Ceratitis capitata, to the volatile constituents of nectarines. Entomol. Exp. Applic., 63: 13-26.
CrossRefDirect Link - Tabilio, M.R., D. Fiorini, E. Marcantoni, S. Materazzi, M. Delfini, F.R. de Salvador and S. Musmeci, 2013. Impact of the Mediterranean fruit fly (medfly) Ceratitis capitata on different peach cultivars: The possible role of peach volatile compounds. Food Chem., 140: 375-381.
CrossRefDirect Link - Nojima, S., C. Linn Junior and W. Roelofs, 2003. Identification of host fruit volatiles from flowering dogwood (Cornus florida) attractive to dogwood-origin Rhagoletis pomonella flies. J. Chem. Ecol., 29: 2347-2357.
CrossRefDirect Link - Biasazin, T.D., M.F. Karlsson, Y. Hillbur, E. Seyoum and T. Dekker, 2014. Identification of host blends that attract the African invasive fruit fly, Bactrocera invadens. J. Chem. Ecol., 40: 966-976.
CrossRefDirect Link - Malheiro, R., S. Casal, S.C. Cunha, P. Baptista and J.A. Pereira, 2016. Identification of leaf volatiles from olive (Olea europaea) and their possible role in the ovipositional preferences of olive fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae). Phytochemistry, 121: 11-19.
CrossRefDirect Link - Hansson, B., M.C. Larsson and W.S. Leal, 1999. Green leaf volatile-detecting olfactory receptor neurones display very high sensitivity and specificity in a scarab beetle. Physiol. Entomol., 24: 121-126.
CrossRefDirect Link - Park, S.J., R. Morelli, B.L. Hanssen, J.F. Jamie, I.M. Jamie, M.S. Siderhurst and P.W. Taylor, 2016. Raspberry ketone analogs: Vapour pressure measurements and attractiveness to Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). PloS One, Vol. 11.
CrossRef - Alagarmalai, J., D. Nestel, D. Dragushich, E. Nemny-Lavy, L. Anshelevich, A. Zada and V. Soroker, 2009. Identification of host attractants for the Ethiopian fruit fly, Dacus ciliatus Loew. J. Chem. Ecol., 35: 542-551.
CrossRefDirect Link - Robacker, D.C., W.C. Warfield and R.A. Flath, 1992. A four-component attractant for the Mexican fruit fly, Anastrepha ludens (Diptera: Tephritidae), from host fruit. J. Chem. Ecol., 18: 1239-1254.
CrossRefDirect Link - Bruce, T.J.A. and J.A. Pickett, 2011. Perception of plant volatile blends by herbivorous insects-finding the right mix. Phytochemistry, 72: 1605-1611.
CrossRefDirect Link - Light, D.M., E.B. Jang and J.C. Dickens, 1988. Electroantennogram responses of the Mediterranean fruit fly, Ceratitis capitata, to a spectrum of plant volatiles. J. Chem. Ecol., 14: 159-180.
CrossRefDirect Link - Konstantopoulou, M.A., F.D. Krokos and B.E. Mazomenos, 2004. Chemical composition of corn leaf essential oils and their role in the oviposition behavior of Sesamia nonagrioides females. J. Chem. Ecol., 30: 2243-2256.
CrossRefDirect Link - Kashiwagi, T., E. Mikagi, D.B. Mekuria, A.D. Boru, S.I. Tebayashi and C.S. Kim, 2005. Ovipositional deterrent on mature stage of sweet pepper, Capsicum annuum, against Liriomyza trifolii (Burgess). Zeitschrift fur Naturforschung C, 60: 739-742.
CrossRefDirect Link - Sasaerila, Y., R. Gries, G. Gries, G. Khaskin, S. King and S. Takacs, 2003. Sex pheromone components of male Tirathaba mundella (Lepidoptera: Pyralidae). Chemoecology, 13: 89-93.
CrossRefDirect Link - De Bruyne, M. and T.C. Baker, 2008. Odor detection in insects: Volatile codes. J. Chem. Ecol., 34: 882-897.
CrossRefDirect Link