
Sri Yusmalinar
School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Tjandra Anggraeni
School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Kustiati
Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Tanjungpura, Pontianak, Indonesia
Indra Wibowo
School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Ramadhani Eka Putra
School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
Intan Ahmad
School of Life Sciences and Technology, Institut Teknologi Bandung, Indonesia
LiveDNA: 62.20145
Journal of Entomology
Year: 2017 | Volume: 14 | Issue: 5 | Page No.: 199-207







ABSTRACT
Background and Objective: Application of insecticide is the most common method to reduce housefly populations. However, in general, not all individuals would receive lethal dose of insecticide during a treatment. This condition could produce favorable biological responses, such as increased reproductive ability, known as hormesis. This study was carried out to observe the effect of topical application of sublethal concentrations or dose of imidacloprid and permethrin to reproductive ability of some strains of housefly of Indonesia. Materials and Methods: Sublethal doses tested in this study were 10 and 30% of LD50 or LC50 (equal to LD5 or LC5 and LD15 or LC15, respectively) of permethrin and imidacloprid. Permethrin was applied by topical method while imidacloprid administrated by feeding method. All treatments were replicated 10 times and assessed for 10 generations to evaluate the long term effect. Mortality of each group was analyzed by probit analysis to determine the value of LD50 or LC50. Differences in fecundity and fertility among different generations and strains were analyzed using two way ANOVA, with a significant value p<0.05 Results: Houseflies that received application of LD5 of permethrin increased its fecundity (17.05% more than application of LD15 and 56.39% more than the control) and fertility (26.73% more than application of LD15 and 63.62% more than the control). Application of LC5 of imidacloprid also increased fecundity of housefly (10.46% more than application LC15 and 45.91% more than the control) and the fertility (17.08% more than application LC15 and 49.17% more than the control). Conclusion: Continuous administration of sublethal doses of permethrin and imidacloprid may induce reproductive hormesis in the housefly through increased fecundity and fertility.
PDF Abstract XML References Citation
Received: September 02, 2017;
Accepted: October 26, 2017;
Published: November 09, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Sri Yusmalinar, Tjandra Anggraeni, Kustiati, Indra Wibowo, Ramadhani Eka Putra and Intan Ahmad, 2017. Reproductive Ability Enhancement of Housefly (Musca domestica Linn) (Diptera: Muscidae) Through Hormesis by Application of Sublethal Doses of Imidacloprid and Permethrin. Journal of Entomology, 14: 199-207.
DOI: 10.3923/je.2017.199.207
URL: https://scialert.net/abstract/?doi=je.2017.199.207
DOI: 10.3923/je.2017.199.207
URL: https://scialert.net/abstract/?doi=je.2017.199.207
INTRODUCTION
Housefly, Musca domestica, is one of the cosmopolitan urban pests that may serve as a mechanical vector of several human diseases, such as typhoid, cholera and dysentery1. In addition, houseflies also considered as urban pest that caused annoyance at residential and area where food is being restored and/or prepared. Application of insecticides is the most commonly used strategy to control the population of houseflies. In Indonesia, synthetic pyrethroid insecticides followed by a relatively new insecticide imidacloprid have been widely used to control housefly population.
However, as it is widely known, housefly is very difficult to control, due to its ability to develop resistance to many types of insecticides2,3. Resistance to insecticides is one of the major problems in attempt to control, both agricultural and urban pests, because it may lower the effectiveness of the insecticides to control the population of pests in question4. Besides, repeated insecticide pressure could lead to the new adaptation strategy by insects related to changes in their environment5.
In the field, concentration of insecticide usually decreases after initial application, which depends on the frequency of insecticide application and degradation by abiotic factors6-8. Under this condition, most individuals received sublethal dose of insecticide. Studies showed that even though sublethal dose did not directly kill the insects, it could produce negative effects on fecundity, life history and longevity9-12.
However, on the other hand, sublethal dose could trigger some positive physiological responses by insects, known as hormesis, such as better growth rate, higher longevity and higher fecundity as shown in Aedes aegypti, Myzus persicae, Plutella xylostella, Tribolium castaneum and Zabrotes subfasciatus13-17.
Development of hormesis in pest population could add further challenges in pest population management. Given the limited information of the hormesis phenomenon on the urban insect pests, especially from Indonesia, there is possibility that the population growth of resistant strains is being accelerated through hormesis. Therefore, the objective of study was to gain a better understanding of the possibility of hormesis, with respect to the improvement in fecundity and fertility, in resistant strains of housefly treated with insecticide for long period.
MATERIALS AND METHODS
Houseflies: In this study, Danish Pest Infestation Laboratory (DPIL) was used as insecticide-susceptible laboratory colony of houseflies. This strain originated from Denmark which has been reared in Laboratory of Entomology, School of Life Sciences and Technology, Institut Teknologi Bandung under tropical environment condition. Resistant strains were collected from six different regions: Serang (SRG), Jakarta (JKT), Bandung (BDG), Semarang (SMG), Yogyakarta (JOG) and Surabaya (SBY) (Table 1).
All adult houseflies were reared in gauze box (30×30×30 cm) and fed with a mixture of sugar, milk and yeast. Ovitrap, made of tissue paper that already dipped in milk, was provided for the nesting. Eggs were kept inside ovitrap until they hatched in 24-48 h. Hatched larvae were transferred to the media made of a mixture of husk, bran and yeast until pupation, usually in 7-8 days. Pupae transferred to gauze box designated for adult flies. This method modified from the method described by Keiding and Arevad18. All research activities were conducted at Laboratory of Entomology, School of Life Sciences and Technology, Institut Teknologi Bandung from April, 2014-October, 2016. During study period, average room temperature was between 25-28°C with RH between 75-80%.
Insecticides: Permethrin (92% active ingredient, PT. Triman Sentosatama Indonesia) and 95% imidacloprid (95% active ingredient, PT. Bayer Indonesia) were suspended in 97% acetone to make 1 L stock solution that was diluted to make the experimental solution. In all experiments, the control treatment was 97% acetone.
Resistance test: Prior to the study, each strain was tested by selected insecticide to obtain lethal concentration 50 (LC50) and lethal dose 50 (LD50). Resistance level to particular insecticide was stated as resistance ratio which was calculated by dividing lethal concentration (LC50) or lethal dose (LD50) of the field population with the corresponding value of the reference susceptible population (in this study was DPIL)19. The results of this preliminary test were shown in Table 2.
| Table 1: | Information on the collection location, year and origin of tested housefly |
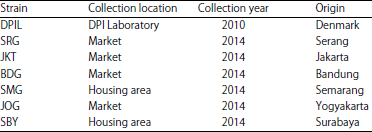 | |
DPIL: Danish pest infestation laboratory, SRG: Serang, JKT: Jakarta, BDG: Bandung, SMG: Semarang, JOG: Yogyakarta, SBY: Surabaya | |
| Table 2: | Comparative toxicity and resistance ratio of permethrin and imidacloprid to houseflies strains from various area in Java |
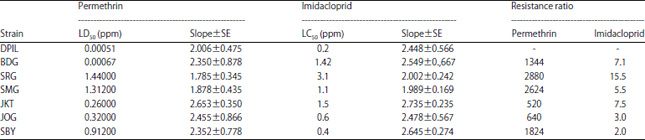 | |
| DPIL: Danish pest infestation laboratory, SRG: Serang, JKT: Jakarta, BDG: Bandung, SMG: Semarang, JOG: Yogyakarta, SBY: Surabaya | |
Bioassay: Three days old houseflies were transferred in groups of 6 individuals, 1 female and 5 males20, to disposable plastic cups, covered with tulle cloth and secured with rubber bands, prior treatment. A water-saturated cotton was placed on the bottom of each cup. Sublethal doses tested in this study were 10 and 30% of LD50 or LC50 (equal to LD5 or LC5 and LD15 or LC15, respectively) of permethrin and imidacloprid (Table 2). Permethrin was applied by topical method while imidacloprid administrated by feeding method21,22. All treatments were replicated 10 times.
After treatment, flies returned to the gauze box where the observation of fecundity and fertility was conducted for 14 days. Some of the eggs were used to continue the study for the next generation. This procedure was conducted for each group until 10 generations.
Statistical analysis: Data on mortality of each group were analyzed using probit analysis using POLO-PC software23 to determine the value of LD50 or LC50. Differences in fecundity and fertility among different generations and strains were analyzed using two way ANOVA, with a significant value p<0.05, followed by least square difference (LSD) test as a post hoc test.
RESULTS
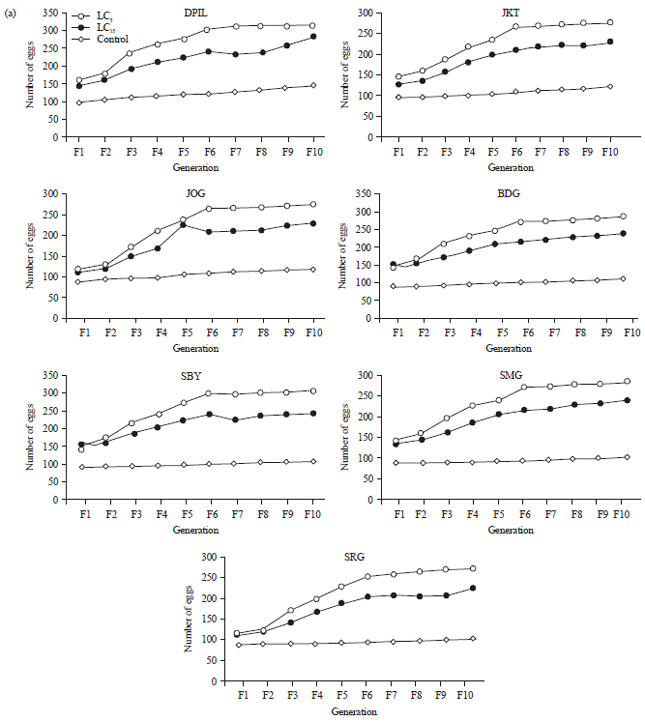
Effect of sublethal doses of imidacloprid to fecundity and fertility: The results showed that application of sublethal doses of imidacloprid significantly increase the fecundity and fertility of resistant strains after 10 generations (p<0.05) (Fig. 1). This study also showed that susceptible strain was more likely to increase their fecundity and fertility as the response to the sublethal dose of insecticide. The DPIL strain increased their fecundity and fertility about 143.5 and 65.5%, respectively, when compared to SRG as the most resistant strain.
However, the fecundity and fertility of first and second generation on all test strains did not differ significantly which indicating that hormesis was not developed instantly. Fecundity and fertility in all strains of flies gradually increased from 3rd-10th generation for all tested concentration. On the other hand, flies that received treatment of LC5 of imidacloprid, on average, improved their fecundity about 10.46 and 45.91% and fertility about 17.08 and 49.17% compared with the ones that received treatment of LC15 and control, respectively.
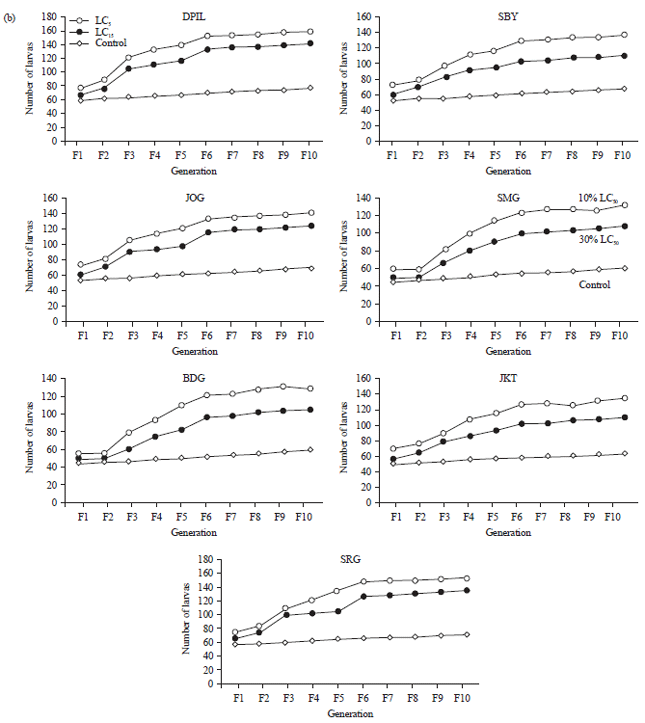
Effects sublethal doses of permethrin to fecundity and fertility: The results showed the application of LD5 and LD15 of permethrin significantly (p<0.05) increase fecundity and fertility in all strains of houseflies after 10 generations compared to control group (Fig. 2). Flies received treatment of LD5 of permethrin also significantly increase their fecundity and fertility compared with the ones treated with LD15. Similar to results of imidacloprid treatment, susceptible strain significantly increased their fecundity and fertility in the rate which higher than any groups use in this study. Compared to the most resistance strain, SRG, the average fecundity and fertility of DPIL strain was 185.8 and 102.6% higher.
Application of LD5 improved the average fecundity about 17.05 and 56.39% and fertility about 26.73 and 63.62% compared with LD15 and control group, respectively. Similar to result of imidacloprid test, DPIL as the most susceptible strain significantly improved their fecundity and fertility more than other resistance strains while SRG as the most resistance strain produced the least improvement.
DISCUSSION
The results showed that application of permethrin and imidacloprid at sublethal doses induced hormesis as reproduction ability of both susceptible and resistance strains significantly increased after 10 generations (p<0.05). The resistance level of resistance to insecticides negatively affected the level of improvement on reproductive ability as less improvement was recorded for strains with higher resistance level. Lower hormesis response in strains with higher resistance level could be caused by higher energy investment for insecticide detoxification (overcompensation theory)24. The results of this study were consistent with several studies on the effect of a sublethal dose of insecticide to agricultural pest, like aphids15,25,26.
Furthermore, this result may explained increasing population of less resistant strain in the field which may related to hormesis response as reported by studies on agricultural pests27-34.
The results also showed that level of fecundity and fertility improvement was much higher in houseflies treated with permethrin. Longer field application of permethrin allowed insects to develop adaptation to toxic, thus, reduced their investment on detoxification process compared to much newer insecticide, like imidacloprid35.
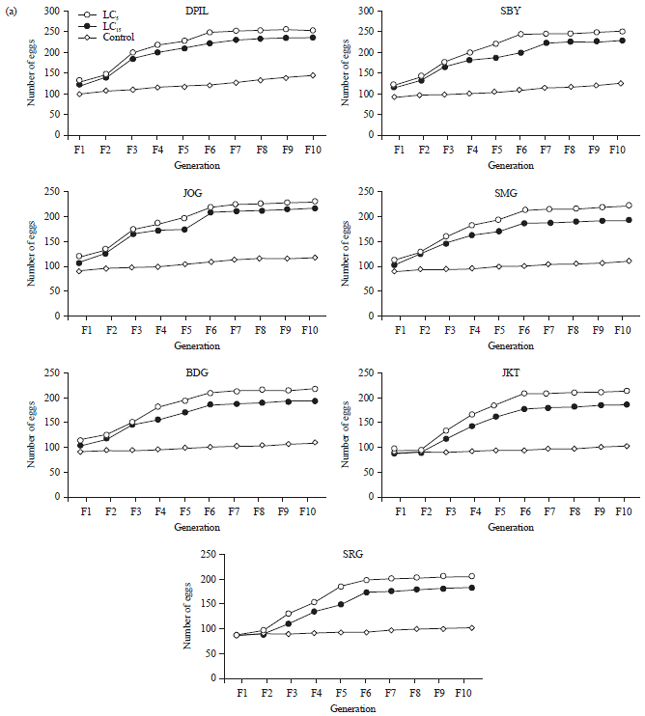  | |
| Fig. 1(a-b): | Response of Musca domestica to sublethal doses of imidacloprid for 10 generations, (a) Fecundity and (b) Fertility |
| DPIL: Danish pest infestation laboratory, SRG: Serang, JKT: Jakarta, BDG: Bandung, SMG: Semarang, JOG: Yogyakarta, SBY: Surabaya | |
It was hypothesized that the increase in reproduction capability was caused by the increase of juvenile hormones and protein total which have roles to regulate egg development. Other researchers also hypothesized the mechanism of reproductive hormesis as a response against exposure to insecticides in sublethal doses, i.e. (1) Increasing the rates of food consumption which may increase production of juvenile hormones and encourage the formation of egg cells in higher numbers36, (2) Increasing the proportion of hatched eggs and vitellin production which related to the increase of eggs production in adult insects37, (3) Inducing the production of sex pheromones that may increase the chance of mating success and egg production38. However, until present day, although many studies on hormesis have been done, the exact mechanism how insects able to increase their fecundity and fertility after had been given the treatment of insecticide in sublethal dose is not yet fully understood (G.C. Cutler, personal communication, December 15, 2016).
Most of the phenomenon of the resurgence in agricultural pest triggered by the loss of natural enemies and parasites are reduced due to insecticide application27,39,40. However, unlike agricultural pests, the resurgence in urban pests is not triggered by the loss of natural enemies since the presence of natural enemies is highly ignored due to human activities. This study showed the possibility of hormesis response, in this case fecundity and fertility, as important mechanism of occurrence of the population explosion of urban pest. Interestingly, similar condition also reported in agricultural pests and other pest groups which indicated that the response is common in the insect world41-44. Under this circumstance, rotation of insecticide types applied and reduction of application periods will be need to be done to prevent development of the hormesis.
 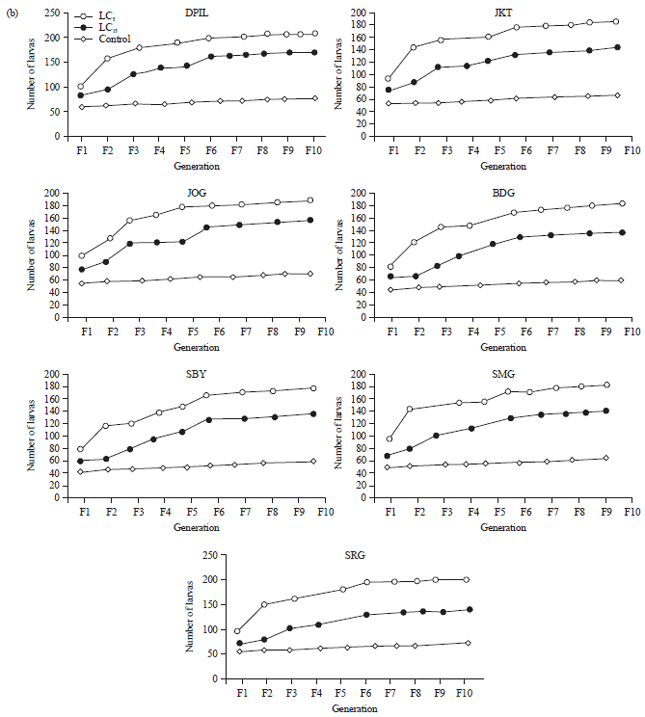 | |
| Fig. 2(a-b): | Response of Musca domestica to sublethal doses of permethrin for 10 generations, (a) Fecundity and (b) Fertility |
| DPIL: Danish pest infestation laboratory, SRG: Serang, JKT: Jakarta, BDG: Bandung, SMG: Semarang, JOG: Yogyakarta, SBY: Surabaya | |
CONCLUSION
During application of insecticide to control the population of urban pest, such as houseflies, not all individual received lethal dose/concentration of insecticide. Houseflies that received sublethal dose/concentration of insecticide, in long term significantly, develop better fecundity and fertility. Under this condition, the possibility of population outbreak could be increased.
SIGNIFICANCE STATEMENT
This study revealed the development of hormesis effect on houseflies that received sublethal doses of insecticide in long term. This study will help the researchers to develop understanding on the insecticide resistance of urban pests. Thus, a better pest control strategy could be developed.
ACKNOWLEDGMENT
Authors are grateful to G. Christoper Cutler (Dalhousie University) and Guedes R.N.C (Carleton University) for valuable discussion and suggestion on the topic being questioned. Authors thank to PT. Triman Sentosatama and PT. Bayer Indonesia for providing insecticides use in this study.
REFERENCES
- Abbas, T., M.A. Nadeem, A. Tanveer, A.Zohaib and T. Rasool, 2015. Glyphosate hormesis increases growth and yield of chickpea (Cicer arietinum L.). Pak. J. Weed Sci. Res., 21: 533-542.
CrossRef - Kustiati, M.I. Tan, S. Yusmalinar, T.B. Ambarningrum and I. Ahmad, 2016. Monitoring permethrin and imidacloprid resistance in Indonesian house fly Musca domestica L. (Diptera: Muscidae). J. Entomol., 13: 40-47.
CrossRefDirect Link - Ahmad, I., S. Astari and M. Tan, 2007. Resistance of Aedes aegypti (Diptera: Culicidae) in 2006 to pyrethroid insecticides in Indonesia and its association with oxidase and esterase levels. Pak. J. Biol. Sci., 10: 3688-3692.
CrossRefPubMedDirect Link - Martins, A.J., C.D.M. Ribeiro, D.F. Bellinato, A.A.Peixoto, D. Valle and J.B.P. Lima, 2012. Effect of insecticide resistance on development, longevity and reproduction of field or laboratory selected Aedes aegypti populations. Plos One, Vol. 7.
CrossRef - Desneux, N., X. Fauvergue, F.X. Dechaume-Moncharmont, L. Kerhoas, Y. Balanger and L. Kaiser, 2005. Diaeretiella rapae limits Myzus persicae populations after applications of deltamethrin in oilseed rape. J. Econ. Entomol., 98: 9-17.
CrossRef - Biondi, A., N. Desneux, G. Siscaro and L. Zappala, 2012. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere, 87: 803-812.
CrossRef - Vanaclocha, P., C. Vidal-Quist, S. Oheix, H. Monton and L. Planes et al., 2015. Acute toxicity in laboratory tests of fresh and aged residues of pesticides used in citrus on the parasitoid Aphytis melinus. J. Pest Sci., 86: 329-336.
CrossRefDirect Link - Adkisson, P.L. and S.G. Wellso, 1962. Effect of DDT poisoning on the longevity and fecundity of the pink bollworm. J. Econ. Entomol., 55: 842-845.
CrossRef - Duncan, J., 1963. Post-treatment effects of sublethal doses of dieldrin on the mosquito Aedes aegypti (L.). Ann. Applied Biol., 52: 1-6.
CrossRefDirect Link - Mebrahtu, Y.B., J. Norem and M. Taylor, 1997. Inheritance of larval resistance to permethrin in Aedes aegypti and association with sex ratio distortion and life history variation. Am. J. Trop. Med. Hyg., 56: 456-465.
CrossRef - Pavela, R., 2007. Lethal and sublethal effects of thymeoil (Thymus vulgaris L.) on the housefly (Musca domestica Lin.). J. Essent. Oil Bear. Plants, 10: 346-356.
CrossRefDirect Link - Ayyanath, M.M., G.C. Cutler, C.D. Scott-Dupree, B. Prithiviraj, S. Kandasamy and K. Prithiviraj, 2014. Gene expression during imidacloprid-induced hormesis in green peach aphid. Dose-Response, 12: 480-497.
Direct Link - Ayyanath, M.M., C.D. Scott-Dupree and G.C. Cutler, 2015. Effect low doses of precocene on reproduction and gene expression in green peach aphid. Chemosphere, 128: 245-251.
CrossRef - Guedes, R.N. and G.C. Cutler, 2014. Insecticide-induced hormesis and arthropod pest management. Pest Manage. Sci., 70: 690-697.
CrossRefDirect Link - Mallqui, K.S., J.L. Viera, R.N. Guedes and L.M. Gontijo, 2014. Azadirachtin-induced hormesis mediating shift in fecundity-longevity trade-off in the Mexican been weevil (Chrysomelidae: Bruchinae). J. Econ. Entomol., 107: 860-866.
CrossRef - Keiding, J. and K. Arevad, 1964. Procedure and equipment for rearing a large number of housefly strains. Bull. World Health Organiz., 31: 527-528.
PubMedDirect Link - Marcon, P.C.R.G., G.D. Thomas, B.D. Siegfried, J.B. Campbell and S.R. Skoda, 2003. Resistance status of house flies (Diptera: Muscidae) from Southeastern Nebraska beef cattle feedlots to selected insecticides. J. Econ. Entomol., 96: 1016-1020.
CrossRefDirect Link - Carillo, J., A. Danielson-Francois, E. Siemann and L. Meffert, 2012. Male-biased sex ratio increases female egg laying and fitness in the housefly, Musca domestica. J. Ethol., 30: 247-254.
CrossRef - Kristensen, M. and J.B. Jespersen, 2004. Susceptibility of spinosad in Musca domestica (Diptera: Muscidae) field populations. J. Econ. Entomol., 97: 1042-1048.
CrossRefDirect Link - Kristensen, M. and J.B. Jespersen, 2008. Susceptibility to thiamethoxam of Musca domestica from Danish livestock farms. Pest Manage. Sci., 64: 126-132.
CrossRefDirect Link - Ayyanath, M.M., G.C. Cutler, G.C, C.D. Scott-Dupree and P.K. Sibley, 2013. Transgenerational shifts in reproduction hormesis in green peach aphid exposed to low concentrations of imidacloprid. Plos One, Vol. 8.
CrossRef - Cutler, G.C., K. Ramanaidu, T. Astatkie and M.B. Isman, 2009. Green peach aphid, Myzus persicae (Hemiptera: Aphididae), reproduction during exposure to sublethal concentrations of imidacloprid and azadirachtin. Pest Manage. Sci., 65: 205-209.
CrossRef - Yu, Y., G. Shen, H. Zhu and Y. Lu, 2010. Imidacloprid-induced hormesis on the fecundity and juvenile hormone levels of the green peach aphid Myzus persicae (Sulzer). Pest. Biochem. Physiol., 98: 238-242.
CrossRef - Chelliah, S. and E.A. Heinrich, 1980. Factors affecting insecticide-induced resurgence of the brown Planthopper, Nilaparvata lugens1 on rice. Environ. Entomol., 9: 773-777.
CrossRefDirect Link - Reissig, W.H., E.A. Heinrichs, and S.L. Valencia, 1982. Insecticides-induced resurgence of the brown planthopper, Nilaparvata lugens, on rice varieties with different levels of resistance. Environ. Entomol., 11: 165-168.
CrossRef - Holt, J., G.A. Norton and D.R. Wareing, 1992. Strategies of insecticide use to avoid resurgence of Nilaparvata lugens (Homoptera: Delphacidae) in tropical rice: A simulation analysis. J. Econ. Entomol., 85: 1979-1989.
CrossRef - Wang, A.H., J.C. Wu, Y.S. Yu, J.L. Liu, J.F. Yue and M.Y. Wang, 2005. Selective insecticide-induced stimulation on fecundity and biochemical changes in tryporyza incertulas (Lepidoptera: Pyralidae). J. Econ. Entomol., 98: 1144-1149.
Direct Link - Yin, J.L., H.W. Xu, J.C. Wu, J.H. Hu and G.Q. Yang, 2008. Cultivar and insecticide application affect the physiological development of the brown planthopper, Nilaparvata lugens (Stal) (Hemiptera: Delphacidae). Environ. Entomol., 37: 206-212.
CrossRefDirect Link - Guedes, R.N., L.C. Magalhaes and L.V. Cosme, 2009. Stimulatory sublethal response of a generalist predator to permethrin: Hormesis, hormoligosis, or homeostatic regulation? J. Econ. Entomol., 102: 170-176.
PubMedDirect Link - Mironidis, G.K., D. Kapantaidaki, M. Bentila, E. Morou, M. Savopoulou-Soultani and J. Vontas, 2013. Resurgence of cotton bollworm Helicoverpa armigera in northern Greece associated with insectiside resistance. Insect Sci., 20: 505-512.
CrossRef - Guedes, R.N., G. Smagghe, J.D. Stark and N. Desneux, 2016. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Ann. Rev. Entomol., 61: 43-62.
CrossRef - Rix, R.R., M.M. Ayyanath and G.C. Cutler, 2016. Sublethal concentrations of imidacloprid increase reproduction, alter expression of detoxification genes and prime Myzus persicae for subsequent stress. J. Pest Sci., 89: 581-589.
CrossRefDirect Link - Yu, H.S., Z.M. Liu, X.Y. Yu, A.Q. Song, N. Liu and H. Wang, 2013. Low-dose radiation induces antitumor effects and erythrocyte system hormesis. Asian Pac. J. Cancer Prev., 14: 4121-4126.
CrossRef - El-Sayed, A.A.A., 2016. Effect of sublethal dose of Hexaflumuron on the ovary and egg maturation of spinybollworm, Earias insulana (Boisd.). J. Entomol., 13: 26-32.
CrossRef - Rabhi, K.K.,K. Esancy, A. Voisin, L. Crespin and J. Le Corre et al., 2014. Unexpected effect of low doses of a neonicotinoid insecticide on behavioral response to sex pheromone in a pest insect. PLos One, Vol. 9.
CrossRef - Hull, L.A.,E.H. Beers and R.L. Meagher, 1985. Impact of selective use of the synthetic pyrethroid fenvalerate on apple pests and natural enemies in large-orchard trials. J. Econ. Entomol., 78: 163-168.
CrossRef - Calabrese, E.J. and L.A. Baldwin, 2003. The hormetic dose-response model is more common than the threshold model in toxicology. Toxicol. Sci., 71: 246-250.
CrossRef - Lockwood, J.A., W.P. Kemp and J.A. Onsager, 1988. Long-term, large-scale effects of insecticidal control on rangeland grasshopper populations (Orthoptera: Acrididae). J. Econ. Entomol., 81: 1258-1264.
CrossRef - Gu, X., F. Gao, and S. Tian, 2009. Interaction between short-term heat pretreatment and fipronil on diamondback moth [Plutella xylostella (Linnaeus)]. Chinese J. Eco-Agric., 17: 709-714.
CrossRef - Gu, X., S. Tian, D. Wang and F. Gao, 2010. Interaction between short-term heat pretreatment and avermectin on 2nd instar larvae of diamondback moth, Plutella xylostella (Linn). Dose-Response, Vol. 7.
CrossRefDirect Link