
Dwi Sutiningsih
Department of Epidemiology and Tropical Disease, Faculty of Public Health, Diponegoro University, Semarang, Indonesia
LiveDNA: 62.16151
Mustofa
Department of Pharmacology, Faculty of Medicine, Gadjah Mada University, Yogyakarta, Indonesia
Tri Baskoro Tunggul Satoto
Department of Parasitology, Faculty of Medicine, Gadjah Mada University, Yogyakarta, Indonesia
Edhi Martono
Department of Plant Pest and Diseases, Faculty of Agriculture, Gadjah Mada University, Yogyakarta, Indonesia
Journal of Entomology
Year: 2017 | Volume: 14 | Issue: 2 | Page No.: 104-111







ABSTRACT
Background and Objective: Bruceine A is a quassinoid compound isolated from the seeds of Makassar fruit (Brucea javanica L. Merr.) that has a biolarvicidal effect toward Aedes aegypti larvae with a lethal concentration fifty (LC50) and ninety (LC90) of 0.408 and 4.923 ppm, respectively. This study aimed to demonstrate the inhibitory effect of bruceine A on the growth and development of A. aegypti larvae at sublethal concentrations. Materials and Methods: The effect of bruceine A on A. aegypti larvae growth and development was tested using bioassays. Daily counts of the number of larvae that survived or died were taken until all the experimental larvae became either pupae or adults. Dead larvae were assessed for morphological and histological changes. The statistical analysis of growth and development inhibition was performed by using Kruskal Wallis test followed by Mann Whitney. The data from observation on morphological and histological changes were descriptively analyzed. Results: Inhibitory effects of bruceine A on the growth and development of A. aegypti larvae were seen at concentrations as low as 0.01 ppm. The morphological changes seen in larvae included damage to the gastrointestinal tract manifested as black spots, folded breathing tubes and detached cuticle and setae as well as smaller body sizes compared to the control group. Histological examination of the larvae indicated diffuse necrosis in the gastrointestinal epithelium accompanied by epithelial cell shrinkage, whereas the core was not visible. Conclusion: It is concluded that bruceine A inhibits growth and development of A. aegypti larvae, as shown by cuticle damage and necrosis of gastrointestinal and respiratory epithelial cells.
PDF Abstract XML References Citation
Received: December 16, 2016;
Accepted: January 31, 2017;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dwi Sutiningsih, Mustofa, Tri Baskoro Tunggul Satoto and Edhi Martono, 2017. Inhibitory Effects of Bruceine A Biolarvicide on Growth and Development of Aedes aegypti Larvae. Journal of Entomology, 14: 104-111.
DOI: 10.3923/je.2017.104.111
URL: https://scialert.net/abstract/?doi=je.2017.104.111
DOI: 10.3923/je.2017.104.111
URL: https://scialert.net/abstract/?doi=je.2017.104.111
INTRODUCTION
The use of larvicides in vector control can reduce vector populations and in turn risk of disease transmission1,2. Larvicidal agents at sufficient concentrations will cause complete or nearly complete mortality in the exposed larval population. However, under various environmental conditions in real practice, achieving uniform coverage of the treated habitat that exposes all target organisms to uniform, lethal concentrations of a larvicide is impossible. As such, in practice, some organisms will have exposure to lethal or above-lethal concentrations, whereas others will be exposed to only sublethal concentrations3.
Cloyd4 stated that long-term, continuous use of insecticides could decrease susceptibility of targets such as mosquitoes. Likewise, Hudayya and Jayanti5 stated that constant pest control using insecticides increases target resistance and negatively affects non-target organisms, including natural enemies (predators) of the target pest. Continuous pest control can also cause environmental pollution. Therefore, research and development of vector control methods that are more environmental friendly and less costly is needed6. One possibility is to use insecticides and larvicides that are derived from plants for vector control. Botanical insecticides/larvicides offer a natural, ‘Environmentally friendly’ approach for pest control as compared to synthetic insecticides. Screening of plant extracts for deleterious effects on insect larvae is one approach used to identify novel botanical insecticides7,8.
Flores et al.9 examined the effects of six types of quassinoid compounds isolated from the plant Scilamea soulameoide on Heliothis virescens and Spodoptera frugiperda larvae and also tested 46 quassinoid compounds from Simaba multiflora against H. virescens and Agrotis ipsilon larvae. They found that these quassinoid compounds inhibited feeding and physical development of the larvae, with an increased mortality of pupae9,10. Meanwhile, four quassinoid compounds, indakuasin, amaderin A, samaderin B and samaderin C, which were isolated from the seeds and bark of Samadera indica plants, showed an inhibitory effect on the development of Spodoptera litura larvae11. Moreover, Syahputra12 and Lina et al.13,14 demonstrated that the active ingredient contained in the extracts of B. javanica (L) Merr. has a larvicidal effect against Crocidolomia pavonana (Lepidoptera: Crambidae). Extracts of B. javanica (L) Merr. can inhibit feeding, decrease growth rates and inhibit nesting of C. pavonana imago14. Sutiningsih and Nurjazuli15 found that brusatol isolated from Brucea javanica (L) Merr. seeds has larvicidal activity against A. aegypti larvae with a lethal concentration fifty and ninety (LC50, LC90) of 0.669 and 8.331 ppm, respectively.
Bruceine A is a quassinoid derived from the fruit of B. javanica (L) Merr.16 plant. Bruceine A ([15]-3-methyl-2-bruseolid butanol) has the molecular formula C26H34O11 and a mass of 522.54 g mol–1. Purified bruceine A exists as an amorphous powder that has a bitter taste. Bruceine A has insecticidal, anti-feeding and growth inhibitory effects against tobacco budworm (Heliothis virescens) and armyworm Spodoptera frugiperda17 as well as larval instar IV of Mexican beetle (Epilachna varivestis Mulsant) and larval instar V of Spodoptera eridania Crawler18. Sutiningsih19 found that bruceine A has larvicidal activity toward A. aegypti with LC50 and LC90 of 0.453 and 4.962 ppm, respectively. These results again demonstrated the potential of bruceine A as a biolarvicide that can be used to control mosquito vectors, especially A. aegypti. Since sub-lethal doses of several chemical larvicides and mosquito control agents can produce delayed effects (i.e., beyond the treatment stage) on surviving larvae3. This study was aimed to assess the inhibitory effect of bruceine A on the growth and development of A. aegypti larvae and examined how sub-lethal concentrations of bruceine A affect insect morphological and tissue histology.
MATERIALS AND METHODS
The study was conducted between January and April 2016. Bruceine A extraction and isolation was carried out at the Pharmaceutical Biology Laboratory, Faculty of Pharmacy, Gadjah Mada University. Experiments examined inhibition of growth and development of A. aegypti (L.) larvae as well as morphology tests were carried out at the Parasitology Laboratory, Faculty of Medicine, Gadjah Mada University. Histological examinations were performed at the Laboratory of Anatomy Pathology, Faculty of Medicine, Gadjah Mada University. The A. aegypti instar III larvae up to the beginning of instar IV were acquired and developed at the Laboratory of Parasitology, Faculty of Medicine, Gadjah Mada University. All commercial reagents and other chemicals used in this study were of analytical quality with the highest purity available and purchased from commercial suppliers.
Bruceine A isolation from B. javanica (L) Merr. seeds: Bruceine A was isolated according to a procedure described by Mangunsong20. Makassar fruit (B. javanica L. Merr.) purchased from a medicinal plant wholesaler (Aneka Herbal Yogyakarta) was analyzed at the Laboratory of Pharmaceutical Biology, Faculty of Pharmacy, Gadjah Mada University, to reconfirm seed identity. Hexane, methanol and dichloromethane used for extraction and isolation of bruceine A were of the highest grade available and were purchased from Merck.
Assay of A. aegypti larvae growth and development inhibition: Inhibition of growth and development of A. aegypti larvae was measured using methods described by Flores et al.3 with slight modifications. Up to 25 larvae were put in plastic cups containing either 100 mL water with bruceine A at various sub-lethal concentrations (0.01; 0.03; 0.05; 0.1; 0.2 and 0.4 ppm) or 100 mL water alone. The cups were kept at room temperature. The number of dead and surviving larvae was counted daily until all the larvae reached pupa or imago stages.
Morphological test: Morphological tests were carried out according to the method described by Sharma et al.21, with slight modifications. The A. aegypti instar III to early instar IV larvae were put in plastic cups containing either 100 mL water (control) or 100 mL water with 0.2 ppm bruceine A. After 24 h the dead larvae were separated and morphologic changes were observed under a light microscope (National optical DC5-163-ASC Digital Binocular Compound Microscope, USA).
Histological test: Histological tests were carried out according to the method of Narciso et al.22, with slight modifications. Histological evaluations of the digestive system were performed using L3 A. aegypti larvae treated with either 100 mL water alone (control) or 100 mL water with 0.2 ppm bruceine A. After fixation in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (Sigma-Aldrich), pH 7.4 for 4 h, the samples were dehydrated with increasing amounts of ethanol (Sigma-Aldrich; 70, 0, 90, 96 and 100%) by immersion in each of the solutions for 15 min. Subsequently, the samples were embedded in Historesin JB4 and the resulting blocks were sliced using a microtome to obtain a series of 3 μm thick sections. The sections were stained with hematoxylin-eosin and then examined and photographed using a light microscope.
Data analysis: The percentage of growth and development inhibition was expressed as Mean±Standard Error of Mean (SEM). Statistical analyses were performed using Kruskal Wallis and Mann Whitney tests to compare differences between the inhibition of growth and development of A. aegypti larvae at various bruceine A concentrations. Statistically significant differences were indicated at p<0.05. The data from observation on morphological and histological changes were descriptively analyzed23.
RESULTS
The complete results for observations concerning bruceine A-mediated inhibition of growth and development of A. aegypti larvae are presented in Table 1. At 0.1 ppm bruceine A, no development of A. aegypti larvae into pupae or imago was seen. Meanwhile, at 0.01 ppm bruceine A, larval development of A. aegypti into pupae and imago still occurred, but at a rate that was 12 and 6%, respectively, that of the control. Kruskal Wallis analysis showed there were significant differences (p<0.05) between growth and development of A. aegypti larvae treated with bruceine A compared to the control, although Mann Whitney test results showed that not all bruceine A concentrations produced significantly different (p<0.05) effects on the development of A. aegypti larvae into a pupa or imago. Overall, bruceine A had a dose-dependent effect on A. aegypti larvae growth wherein 0.01 ppm bruceine A, the lowest concentration tested also had the smallest effect (Table 1).
In terms of morphology, A. aegypti larvae treated with 0.2 ppm bruceine A had structural damage in the digestive tract that was manifested as black spots. The treated larvae also had abnormally folded respiratory tubes as well as detached setae and cuticles (Fig. 1b).
| Table 1: | Percentage of larval A. aegypti pupae and imago after treatment with bruceine A at sub-lethal concentrations relative to control larvae |
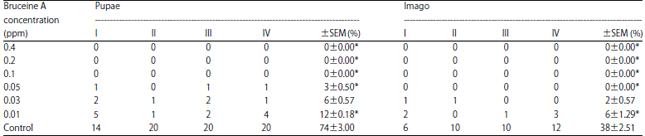 | |
*p<0.05, when compared with control, Kruskal Wallis, Mann Whitney, No. of A. aegypti larvae on each test: 25, ppm: Part per million, SEM: Standard error of mean | |
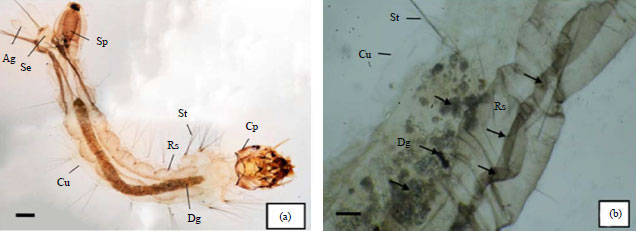 | |
| Fig. 1(a-b): | Control A. aegypti larvae and larvae treated with 0.2 ppm bruceine A, (a) Control (without treatment) (40X) gastrointestinal and respiratory tracts are complete and intact and (b) Larva treated with 0.2 ppm bruceine A (100X) damage to the digestive tract, respiratory tracts, setae and cuticle is visible |
Cp: Caput, Dg: Digestive tract, Rs: Respiratory tract, St: Setae, Cu: Cuticle, Sp: Siphon, Se: Saddle, Ag: Anal gills | |
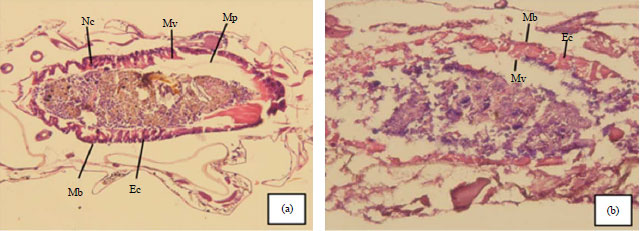 | |
| Fig. 2(a-b): | Longitudinal sections of gastrointestinal tracts from untreated A. aegypti larvae and larvae treated with 0.2 ppm bruceine A, (a) Control (without treatment) (400X). The shape and size of the gastrointestinal epithelial cells appear normal and (b) Treatment with 0.2 ppm bruceine A (400X). Gastrointestinal epithelial cell necrosis is apparent |
Mb: Basement membrane, Mp: Peritrophic membrane, Mv: Microvilli, Ec: Epithelial cells, Nc: Nucleus | |
Control larvae that were not exposed to bruceine A showed no morphological changes in any part of the body (Fig. 1a).
Histological analysis of A. aegypti larvae after treatment with 0.2 ppm bruceine A showed diffuse necrosis of gastrointestinal epithelial cells, characterized by shrunken cells and a diminished core (karyolysis). Although necrotic epithelial cells still attached to the basement membrane, microvilli and the peritrophic membrane were necrotic and diffuse. In addition, the arrangement of gastrointestinal epithelial cells was disorganized (Fig. 2b). In contrast, cells in control larvae showed no histological damage, as the shape and size of gastrointestinal epithelial cells was normal and had a compactly-stained cytoplasm, spherical core and clear chromatin threads. Moreover, the gastrointestinal epithelial cells had a regular composition and the peritrophic membrane remained visible. Most gastrointestinal epithelial cell microvilli appeared normal and entire epithelial cells still attached to the basement membrane (Fig. 2a).
DISCUSSION
In this study, untreated A. aegypti larvae development normally from pupae to imago at rates of 74±0.90 and 38±0.76%, respectively. Meanwhile, A. aegypti larvae treated with bruceine A at concentrations as low as 0.01 ppm had inhibited growth and development.
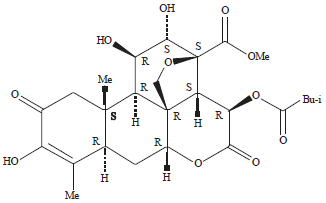 | |
| Fig. 3: | Chemical structure of bruceine A |
| Source: Feng et al27 | |
As the bruceine A concentration increased to 0.05 ppm, only 3% progressed to the pupae stage and none transitioned from pupa to imago, whereas bruceine A treatments above 0.05 ppm had completely suppressed larvae development. Kruskal-Wallis analysis showed significant differences (p<0.05) in the effects of various bruceine A concentrations. This finding is consistent with that of Yunita et al.24 for the effects of the biolarvicide extracted from Eupatorium riparium leaves on A. aegypti larval development.
The mechanism by which bruceine A suppresses larval growth and development likely involves an inhibitory effect on feeding (antifeedant). The bitter taste of bruceine A may impair larvae feeding patterns leading to starvation and eventual retardation of growth and development into pupae. Antifeedant compounds typically inhibit insect feeding activity, but they do not directly kill, expel or entrap insects. Indeed, Syahputra12 reported that 0.06-0.25% B. javanica (L.) Merr. extracts can inhibit feeding activity and decrease the growth rate of Crocidolomia pavonana larvae, whereas concentrations of 0.12-3.0% inhibited C. pavonana nesting by 66.2-96.5% relative to control. A decrease in the rate of feeding after treatment with Brucea javanica fruit extract indicates that this fruit contains active components that are either antifeedants or have toxic effects that disturb normal feeding as well as digestion and food absorption. Similar results reported by Zhang et al.25, brusatol isolated from B. javanica has antifeedant effects on Spodoptera exigua larvae.
Antifeedant compounds inhibit appetite (feeding inhibition) in insects. Antifeedants also suppress and deter the biting activity and continuation of feeding26. Bioactive compounds can directly or indirectly kill insects in a short time frame and antifeedants fall into the indirect category9. At a molecular level, compounds with antifeedant activity typically have: a carbonyl group in ring A; an α, β unsaturated carbonyl or a methylenedioxy bridge on ring C and a θ-lactone group on ring D27. The bruceine A structure consists of five fused rings containing a heterocyclic oxygen ring and lactone ring, which is consistent with its antifeedant activity (Fig. 3). Similar to other antifeedant compounds, quassinoids such as bruceine A can kill larvae indirectly by interfering with signal transduction pathways that stimulate feeding (phagostimulant) of A. aegypti larvae. Failure to transmit taste stimuli renders larvae unable to recognize surrounding food sources, where a slow feeding activity results in insufficient energy stores to drive larval growth and development that leads to eventual death.
The results of this study are consistent with those of Leskinen et al.18, which demonstrated that bruceine A has antifeedant activity on fourth instar Mexican beetle (Epilachna varivestis Mulsant) larvae and fifth instar larvae of Spodoptera eridania Crawler. Klocke et al.17 also showed that bruceine A has insecticidal activity, as well as antifeedant and growth inhibition activity against tobacco budworm (Heliothis virescens) and Spodoptera frugiperda. Inhibition of larval growth and development to pupa and then imago may also be caused by disruption of growth hormone production. Chapman28 found that insect molting and metamorphosis depends on several growth hormones. Disruption of production of one type of hormone due to inhibited cell respiration processes in a hormone-producing organ can impact overall hormone function to inhibit insect growth and development.
Bruceine A as a biolarvicide can also affect A. aegypti larvae behavior after entering the imago phase, as bruceine A treatment can slow the emergence of imago and shorten the imago lifespan. Treatment with 0.01 ppm bruceine A in this study resulted in only 12% of A. aegypti larvae developing into pupae and of these only 6% reached the imago stage and the percentages were even lower for 0.03 and 0.05 ppm bruceine A (Table 1). According to Govindachari et al.11, quassinoid compounds can change feeding behavior and regulate insect growth as was seen for the quassinoid kaparramarin29 from Castela tortuosa plants that inhibits growth of Heliothis virescens.
Bruceine A may also inhibit larval growth by binding dietary proteins that in turn blocks food digestion. Moreover, the bitter taste of bruceine A can irritate the stomach following ingestion30. Bruceine A enters through the larval body wall and mouth because larvae usually ingest food from their surroundings. According to Lu and Kacew31 and Matsumura32, the body wall of insects can absorb toxic substances in large quantities. These substances can then easily penetrate the cuticle and farther into the insect body because insects are generally so small that the body surface area is exposed to relatively higher concentrations than that of mammals33,34. In addition, the hydrophobic and lipophilic nature of the insect cuticle allows easy penetration by non-polar bioactive compounds. Thus, bruceine A disrupts both the epicuticle lipid and protein layer before entering the larva body. Moreover, bruceine A dissolved in the lipid cuticle will accelerate the rate of penetration of toxins into the hemocoel (body cavity). The rate of cuticle penetration of toxic compounds such as bruceine A depends in part on the structure and thickness of the cuticle in a given body section. Toxic substances tend to enter through larval body parts that are coated by a thin cuticle, such as inter-segment membranes, membranes at joints and chemoreceptors on the tarsus35. The amount of toxic substances that enter causes the cuticle and the setae feathers of the larvae to become damaged/detached (Fig. 1b). Furthermore, these toxic substances are absorbed through the body wall of the insects and are then transported in bodily fluids to sites of action. Toxic compounds such as bruceine A can disrupt the digestive, respiratory and nervous system in larvae. The digestive tract of mosquito larvae can be divided into three regions: front, middle and back, wherein food digestion and absorption occurs in the middle of the gastrointestinal tract or midgut36. The middle gastrointestinal tract/midgut is covered by epithelial tissue and cells of this tissue can be lysed by toxic substances and in turn undergo necrosis (Fig. 2b). Such necrosis can reduce surface tension of midgut mucous membranes that then inhibits food digestion and absorption and eventually compromises survival37.
Bruceine A as a contact and abdominal poison can kill A. aegypti larvae by damaging its skin/cuticles and digestive system, which disrupts molting/metamorphosis and food absorption, respectively, to inhibit overall growth and development of the larvae (life cycle). Bruceine A can also decrease the rate of transition from pupa to imago to decrease populations of mosquitoes acting as disease vectors. Results from this study reinforce the potential of bruceine A as a biolarvicide for control of disease vectors, especially A. aegypti. Future studies should examine how bruceine A affects growth hormone production as well as molting and metamorphosis.
CONCLUSION
Bruceine A as biolarvicide can inhibit the growth and development of A. aegypti larvae by promoting damage to the gastrointestinal tract/midgut and cuticles as well as necrosis of the gastrointestinal epithelial cells in the midgut. Bruceine A has the potential to be used as a biolarvicide to control various disease vectors, especially A. aegypti.
SIGNIFICANCE STATEMENT
Findings from this study could contribute to a better understanding of the mechanism of action of bruceine A larvicide, which is derived from Makassar fruit. These results also support the potential of natural, plant-based larvicides for environmentally sensitive control of disease vectors such as A. aegypti.
ACKNOWLEDGMENTS
We are deeply grateful to drh. Sitarina Widyarini and drh. Sugiyono, Department of Pathology, Faculty of Veterinary Medicine who helped to interpret the histological preparations. We also thank Dra. Rr. Upik Ngesti WA, DAP and E, MBiomed from the Department of Parasitology, Faculty of Biology, University of Gadjah Mada who helped to interpret the morphological preparations in this study.
REFERENCES
- Candra, A., 2010. [Dengue haemorhagic fever: Epidemiology, pathogenesis and risk factor of transmission]. Aspirator, 2: 110-119, (In Indonesian).
Direct Link - Sanchez, L., V. Vanlerberghe, L. Alfonzo, M.D.C. Marquetti, M.G. Guzman, J. Bisset and P. van der Stuyft, 2006. Aedes aegypti larval indices and risk for dengue epidemics. Emerging Infect. Dis., 12: 800-806.
Direct Link - Flores, A.E., G.P. Garcia, M.H. Badii, M.A.R. Tovar and I.F. Salas, 2004. Effects of sublethal concentrations of vectobac on biological parameters of Aedes aegypti. J. Am. Mosq. Control Assoc., 20: 412-417.
PubMedDirect Link - Cloyd, R., 2004. Natural indeed: Are natural insecticides safer and better than conventional insecticides? Illinois Pesticide Rev., 17: 1-7.
Direct Link - Gray, M., S. Ratcliffe and M. Rice, 2009. The IPM Paradigm: Concepts, Strategies and Tactics. In: Integrated Pest Management: Concepts, Tactics, Strategies and Case Studies, Radcliffe, E.B., W.D. Hudchison and R.E. Cancelado (Eds.). Chapter 1, Cambridge University Press, Cambridge, UK., ISBN-13: 9780521875950, pp: 3-10.
- Leatemia, J.A. and M.B. Isman, 2004. Insecticidal activity of crude seed extracts of Annona spp., Lansium domesticum and Sandoricum koetjape against lepidopteran larvae. Phytoparasitica, 32: 30-37.
CrossRefDirect Link - Flores, G., L. Hilje, G.A. Mora and M. Carballo, 2008. Antifeedant activity of botanical crude extracts and their fractions on Bemisia tabaci (Homoptera: Aleyrodidae) adults: III. Quassia amara (Simaroubaceae). Revista Biologia Tropical, 56: 2131-2146.
Direct Link - Martinez, M.L., G. von Poser, A. Henriques, M. Gattuso and C. Rossini, 2013. Simaroubaceae and picramniaceae as potential sources of botanical pesticides. Ind. Crop Prod., 44: 600-602.
CrossRefDirect Link - Govindachari, T.R., G.N.K. Kumari, G. Gopalakrishnan, G. Suresh, S.D. Wesley and T. Sreelatha, 2001. Insect antifeedant and growth regulating activities of quassinoids from Samadera indica. Fitoterapia, 72: 568-571.
CrossRefDirect Link - Syahputra, E., 2008. [Bioactivity of Brucea javanica fruit as nabatical insectiside for agricultural insect pest]. Bul. Littro, 19: 56-67.
Direct Link - Lina, E.C., Dadang, S. Manuwoto, G. Syahbirin and D. Prijono, 2013. Synergistic action of mixed extracts of Brucea javanica (Simaroubaceae), Piper aduncum (Piperaceae) and Tephrosia vogelii (Leguminosae) against cabbage head caterpillar, Crocidolomia pavonana. J. Biopesticides, 6: 77-83.
Direct Link - Sutiningsih, D. and Nurjazuli, 2017. Effect of brusatol biolarvicide administration on behavioral response of Aedes aegypti and its toxicity on vero cells. J. Biol. Sci., 17: 127-135.
CrossRefDirect Link - Bawm, S., H. Matsuura, A. Elkhateeb, K. Nabeta and Subeki et al., 2008. In vitro antitrypanosomal activities of quassinoid compounds from the fruits of a medicinal plant, Brucea javanica. Vet. Parasitol., 158: 288-294.
CrossRefPubMedDirect Link - Klocke, J.A., M. Arisawa, S.S. Handa, A.D. Kinghorn, G.A. Cordell and N.R. Farnsworth, 1985. Growth inhibitory, insecticidal and antifeedant effects of some antileukemic and cytotoxic quassinoids on two species of agricultural pests. Cell. Mol. Life Sci., 41: 379-382.
CrossRefDirect Link - Leskinen, V., J. Polonsky and S. Bhatnagar, 1984. Antifeedant activity of quassinoids. J. Chem. Ecol., 10: 1497-1507.
CrossRefDirect Link - Sharma, A., S. Kumar and P. Tripathi, 2015. Impact of Achyranthes aspera leaf and stem extracts on the survival, morphology and behaviour of an Indian strain of dengue vector, Aedes aegypti L. (Diptera: Culicidae). J. Mosquito Res., Vol. 5.
CrossRef - Narciso, J.O.A., R.O. de Araujo Soares, J.R. dos Santos Mallet, A.E. Guimaraes, M.C. de Oliveira Chaves, J.M. Barbosa-Filho and M. Maleck, 2014. Burchellin: study of bioactivity against Aedes aegypti. Parasites Vectors, Vol. 7.
CrossRefDirect Link - Yunita, E.A., N.H. Suprapti and J.W. Hidayat, 2009. [Effect of Teklan leaf extract (Eupatorium riparium) on mortality and growth of Aedes aegypti larvae]. Bioma, 11: 11-17, (In Indonesian).
Direct Link - Zhang, L., X. Feng, D. Ma, J. Yang, H. Jiang, Y. Zhang and W. He, 2013. Brusatol isolated from Brucea javanica (L.) Merr. induces apoptotic death of insect cell lines. Pesticide Biochem. Physiol., 107: 18-24.
CrossRefDirect Link - Herlt, J.H., L.N. Mander, E. Pongoh, R.J. Rumampuk and P. Tarigan, 2002. Two major saponins from seeds of Barringtonia asiatica: Putative antifeedants toward Epilachna sp. larvae. J. Nat. Prod., 65: 115-120.
CrossRefDirect Link - Feng, X.H., Y.N. Zhang, W.Z. He, L. Zhang and H.Y. Jiang, 2010. Bruceine A. Acta Crystallographica Sect. E: Struct. Rep. Online, 66: o854-o855.
CrossRefDirect Link - Bernardi, O., P.M. Dourado, R.A. Carvalho, S. Martinelli, G.U. Berger, G.P. Head and C. Omoto, 2014. High levels of biological activity of Cry1Ac protein expressed on MON 87701 × MON 89788 soybean against Heliothis virescens (Lepidoptera: Noctuidae). Pest Manage. Sci., 70: 588-594.
CrossRefDirect Link - Shahida, A.N., T.W. Wong and C.Y. Choo, 2009. Hypoglycemic effect of quassinoids from Brucea javanica (L.) Merr. (Simaroubaceae) seeds. J. Ethnopharmacol., 124: 586-591.
CrossRefDirect Link - Krieger, R.I., 2010. Haye's Handbook of Pesticide Toxicology. 3rd Edn., Academic Press, Cambridge, Massachusetts, ISBN: 978-0-12-374367-1, Pages: 2342.
Direct Link - Dono, D., S. Ismayana, Idar, D. Prijono and I. Muslikha, 2010. [Status and biochemical resistance of Crocidolomia pavonana (F.) (Lepidoptera: Crambidae) to organophosphate insecticide and its sensitivity to botanical insecticide]. J. Entomol. Indonesia, 7: 9-27, (In Indonesian).
Direct Link - Dharmagadda, V.S.S., S.N. Naik, P.K. Mittal and P. Vasudevan, 2005. Larvicidal activity of Tagetes patula essential oil against three mosquito species. Bioresour. Technol., 96: 1235-1240.
CrossRefDirect Link