
Vimalanathan Arunprasanna
Laboratory of Insect Molecular Biology, Department of Environmental Biotechnology, Bharathidasan University, Tiruchirappalli 620 024, Tamil Nadu, India
LiveDNA: 91.12520
Mani Kannan
Laboratory of Insect Molecular Biology, Department of Environmental Biotechnology, Bharathidasan University, Tiruchirappalli 620 024, Tamil Nadu, India
LiveDNA: 91.12522
Sankarappan Anbalagan
Department of Zoology, Sethupathy Government Arts College, Ramanathapuram 623502, Tamil Nadu, India
LiveDNA: 91.4208
Muthukalingan Krishnan
Laboratory of Insect Molecular Biology, Department of Environmental Biotechnology, Bharathidasan University, Tiruchirappalli 620 024, Tamil Nadu, India
LiveDNA: 91.12523







ABSTRACT
Background: The complete understanding of feeding and reproduction strategies during metamorphosis of silkworm Bombyx mori is very essential, for that a comparative proteomics analysis was used to investigate the proteins extracted from the head of the larva and adult of Bombyx mori. Materials and Methods: Proteins were separated and identified by using 2D-PAGE and MALDI-TOF-TOF-MS analysis respectively. The functions of each proteins were annotated using STRAP GO analysis. The expression level of each protein spots between larvae and adult were analyzed using 2D image platinum 7. Results: Using 2-DE gels stained with coomassie staining revealed, 315±3 and 296±3 proteins spots with pI ranges of 4-10 and 3.5-8 in the larva and adult head, respectively. Thirty proteins were identified based on differential and unique expression in the head of the larva and adult of B. mori. Among the 30 identified proteins, comparatively 10 proteins were up regulated and 6 proteins were down regulated in the larvae, whereas 6 proteins were up regulated and 10 were down regulated in the adult. In addition, 6 and 8 unique proteins expressions were observed in the larvae and adult, respectively. Database search combined with STRAP GO analysis revealed that few up regulated and unique protein in the larvae such as odorant binding proteins, argonaute protein, apolipophorin III precursor, enolase and iron-ion transport proteins were localized to regulate the feeding activity of larva. Furthermore, some proteins are highly and uniquely expressed from adult head such as glycoprotein hormone, N-acetyltranferase, cytochrome p450, DNA binding and intracellular transport protein, clathrin coat assembly protein and metabolic enzyme which may regulate adult ecolsion and reproduction. Conclusion: The results suggest and confirmed that the protein expression in each stage could alter the behaviors of the insect related to larval feeding, growth and reproduction in the adult. Further the characterization and functions of other proteins identified are discussed. This study will help to improve the silk production which leads to economy improvement in sericulture industry.
PDF Abstract XML References Citation
Received: August 05, 2016;
Accepted: November 07, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Vimalanathan Arunprasanna, Mani Kannan, Sankarappan Anbalagan and Muthukalingan Krishnan, 2017. Comparative Proteomic Analysis of Larva and Adult Heads of Silkworm, Bombyx mori (Lepidoptera: Bombycidae). Journal of Entomology, 14: 1-12.
DOI: 10.3923/je.2017.1.12
URL: https://scialert.net/abstract/?doi=je.2017.1.12
DOI: 10.3923/je.2017.1.12
URL: https://scialert.net/abstract/?doi=je.2017.1.12
INTRODUCTION
An insect head is composed of several proteins and is specialized for structural development, remodelling, neural signalling, olfaction include feeding and reproduction. Although head proteins of insect are primarily involving for structural development and remodelling, focusing on its feeding and reproduction strategy is vital to develop novel technology for control of insect pests and increase the yield of products from beneficial insects. In this study, the Odorant Binding Proteins (OBPs) act as a carrier for volatile compounds and mediate the signal to specific olfactory receptors for feeding and reproduction behaviours1,2. Tanaka et al.3 also highlighted the involvement of head proteins includes olfactory receptors for the highly selective tuning of a silkworm to feeding mulberry leaves. In addition, gene expression profile in the head of silkworm at different stage has been studied by Xia et al.4. Nevertheless, there is no link between the expression profile and translation products5. Hence, the proteomics study is very important to focus on the active products in the cells rather than gene expression study by using microarray and RT-PCR analysis6,7.
Two-dimensional gel electrophoresis combined with mass spectrometry analysis has been frequently used in classical proteomics research to identify and functional characterization of the specific proteins from different organs and tissues of Bombyx mori such as collateral glands8, haemolymph9-11, fat body12,13, silk glands14,15, protharasic gland16, endocrine organs17, larval head, diapauses and non-diapauses egg and embryo18-20, integument, trachea and adult scale21, midgut and peritrophic membrane22-25, larval gonads and testis 26,27, Malpighian tubules28, wing discs29 and newly hatched larva30. Further, proteome profiling of several organs and tissues analysis revealed that these proteins play a crucial role in the nervous system development, digestion, protein synthesis, transport of ions and O2, reproduction, stress response and apoptosis in silkworm Bombyx mori.
Particularly, the involvements of head proteins in the feeding and reproduction strategy during metamorphosis of B. mori are still unclear and received few attentions. The present study would focus to identify the functional traits in the larva and adult of B. mori is mediated by the head proteins. Hence, the objective of the present study was to identify the functional characterization of the head proteins in the larva and adult of B. mori (Bombycidae: Lepidoptera) by using two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) with MALDI-TOF-TOF/MS analysis.
MATERIALS AND METHODS
Rearing: A hybrid strain (Tamil Nadu white×NB4D2, Bombyx mori) of the silkworm was used for this experiment, which was maintained in our Insect Molecular Biology Laboratory at Bharathidasan University, Tamil Nadu, India. Disease free eggs of B. mori were kept at ambient temperature (27±2°C) and a relative humidity (75±5%) until hatching of larvae as reported previously31. The hatched larvae fed with chopped tender leaves of the mulberry variety (MR2) until III instar and with coarse leaves provided until the end of the last instar.
Protein extraction: The heads were collected from 20 individuals of 5th instar larvae and freshly emerged adults of B. mori, washed twice with milli-Q water taken in a centrifuge tube and stored at -85°C for further use. The heads were grounded in 200 μL of extraction buffer containing 7 M urea, 2 M thio urea, 4% CHAPS, 20 mM DTT, 5 mM EDTA, 2 mM PMSF, 1.2% biolyte, 10% glycerol, RNase (1 mg mL–1) and DNAse (1 mg mL–1). The mixture was centrifuged at 15,000 rpm for 20 min at 4°C. The supernatant was collected for further centrifugation at 15,000 rpm for 20 min at 4°C. The supernatant was prepared as aliquots and stored at -85°C until electrophoresis performed. The protein concentrations were determined according to Bradford method32.
02D-elecrophoresis: A 300 μg of protein taken from the each head sample and mixed in 350 μL of rehydration buffer containing 7 M urea, 2 M thiourea, 2% CHAPS, 65 mM DTT 0.8% biolytes, 0.002% bromophenol blue. Each sample was loaded onto 17 cm immobiline dry strips (17 cm, pH 3-10 Linear, Bio-rad) to passive rehydration for overnight. Iso Electric Focusing (IEF) was performed using Ettan IPGphor 3 U (GE healthcare) at 20°C. The IEF program was set as: Step 500 V for 1 h, gradient 3000 V for 1 h, gradient 8000 V for 3 h and step 8000 V for 7 h (a total of 55 028 Vh). The isoelectric focused strips were incubated for 15 min in an equilibration buffer containing 50 mM tris-HCl buffer pH 8.8, 6 M urea, 2% SDS and 30% glycerol and 1% DTT (1%) and the strip was again equilibrated for another 15 min with same buffer except that DTT was replaced with 2.5% iodoacetamide. The equilibrated strips were transferred to 12% SDS-PAGE33 and sealed with 0.5% agarose to prevent the shaking of the strip on the top of the PAGE. The SDS-PAGE was performed at 15°C using protean II Xi cell unit (Bio-rad) at constant power of 2 W gel–1 for 1 h followed by 6 W gel–1 until the bromophenol blue frontier reached the bottom of the gel. The gel was stained for 2 h or overnight in 0.1% of coomassie brilliant blue R-250 solution (Sigma) in 40% methanol and 10% glacial acetic acid. Then the gel was rinsed thrice with milli-Q for a minute and the remain stain was removed using destaining solution containing 10% glacial acetic acid and 40% methanol in milli-Q until clear protein spots were visualized. Then the gel was stored in 10% ethanol at 4°C if for long times. Molecular mass and pI were calculated from digitalized 2D images using protein ladder (Bench MarkTM protein ladder, life technologies, Cat No: 10747-012). Triplicate analysis was performed for each sample and a comparison of relative intensity of protein spots between the larvae and adult heads were conducted.
2D-PAGE spot analysis: Two-dimensional spot images were scanned at an optical resolution of 400 dpi with gray scale using high-resolution image scanner. The scanned gels were analyzed using Image MasterTM 2D platinum 7.0 software according to the instruction manual given by GE healthcare. The triplicate images were merged and subjected to analysis including background subtraction, spot detection, spot matching and quantitative intensity normalization analysis, etc. All the statistical analysis was done with PAST version 2.08.
In-gel trypsin digestion: Based on the comparative and the differential expression, 30 protein spots were manually excised from the Coomassie blue stained gel and subjected to in-gel trypsin digestion34. Briefly, the gels were washed twice in milli-Q water. The gel pieces were placed in a solution containing 50% of methanol and then dehydrated by acetonitrile (ACN) and dried in a SpeedVac (Thermo Savant). The dried gel pieces were rehydrated using 0.02 μg μL–1 trypsin and 25 mM ammonium bicarbonate containing a 10% ACN solution on ice and digested at 37°C for 16 h. The enzymatic reactions were stopped using 50 μL of a solution including 5% trifluoroacetic acid (TFA) and 67% ACN. After centrifugation, the peptide mixture was transferred into a centrifuge tube and sonicated for 15 min and then concentrated in a SpeedVac before being analyzed in a mass spectrometer. These MS/MS analysis were replicated twice for each protein sample in this experiment.
MALDI-TOF/TOF MS analysis and database searching: Prepared peptide samples w e re resolved in 0.1% TFA, mixed with an equal volume of matrix solution α-cyano-4-hydroxy-cinnamic acid (CHCA, Sigma, St., Louis, MO) and spotted on the target plate. The samples were allowed to air-dry and analyzed UltrafleXtreme MALDI-TOF/TOF-MS/MS (Bruker Daltonik, UK). Protein identification using peptide mass fingerprinting was performed using the Mascot search engine version 2 (Matrix Science, London, UK) against the NCBI and Swiss-prot protein database. The following search parameters were used: Taxonomy: Bombyx mori, Database: NCBI nr database or Swiss-prot database, enzyme: trypsin with one max missed cleavage, monoisotopic masses, fixed modifications of carbamidomethyl, variable modifications of oxidation on methionine, peptide charge state of 1+ and peptide MS/MS tolerance of ±0.7. Protein identification with significant hits as defined by a mascot probability analysis (Protein score greater than 95%, p<0.05) was accepted. The identified proteins were further classified based on their biological, cellular and molecular functions based on the functional annotations in the software tool for rapid identification of proteins (STRAP online tool, version 0.15)35.
RESULTS
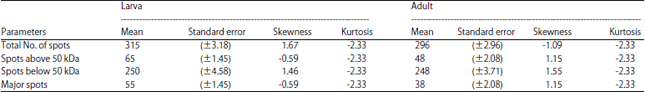
2D-PAGE pattern of proteins from larvae and adult head of B. mori: The greatest difference between the 2D gel patterns of head proteins from larval and adult of B. mori was showed in (Fig. 1, 2). The triplicates of each head proteins patterns were highly reproducible and acceptable. In total, ~315±3 and 296±3 protein spots were detected from larvae and adult heads, respectively in the pI range of 3-10 and molecular masses of ~5-120 kDa (Table 1). In this 315±3 spots on the larvae head, around 65±1 spots are present at >50 kDa and the spots present below 50 kDa are nearly 250±5, whereas 55±1 spots are observed as a major spot in larvae. In the adults head, 296±3 protein spots were observed of which 48±2 proteins are present above 50 kDa and 248±4 spots below 50 kDa and 38±2 spots are observed to be the major spots in adult head out of 224. The variation of proteins spots was measured using one-way ANOVA.
| Table 1: | Univariate statistics of proteins spots in heads from larva and adult of Bombyx mori obtained by 2-DE |
 | |
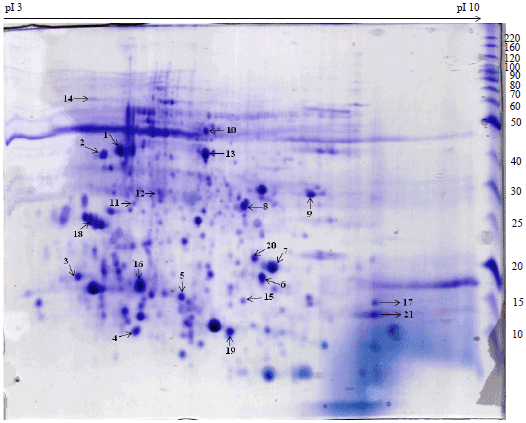 | |
| Fig. 1: | 2D-PAGE analysis of proteins from larvae head of Bombyx mori |
 | |
| Fig. 2: | 2D-PAGE analysis of proteins from adult head of Bombyx mori |
| Table 2: | Distributional variation of protein spots obtained from the head of larva and adult of Bombyx mori using 2-DE |
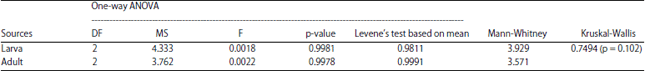 | |
Intra and inter variation of protein spots distributed in 2-DE in the heads of larva and adult were not significantly varied and this result was tested with Levene’s test based on mean (Table 2). The variation of protein spots of 2-DE between the larva and adult was tested with Mann-Whitney and Kruskal-Wallis analyses (Table 2). The larval head protein spots were more significant than the adult proteins spots revealed by these analyses and it was further tested with Monte Carlo (0.8076). The theoretical isolelectric point (pI) and Molecular Weight (MW) of the identified proteins were calculated using the compute pI/MW tool (http://web.expasy.org/compute_pi/) according to the predicted amino acid sequence. The results showed that distribution of major protein spots in the larvae within the range of pI 5-7 and in adult within the range of pI 4-5 and 7-9 (Fig. 3). The spots present in between each pI is calculated to be 11±1, 64±1, 132±3, 77 ±1, 20±1, 9±1 and 3±1 for 3-4, 4-5, 5-6 , 6-7, 7-8, 8-9 and 9-10 pI, respectively. The spots present in each pI is calculated to be 9±2 spots in 3-4, 66±1 spots in 4-5, 117±1 spots in 5-6, 62±2 spots in 6-7, 31±1 spots in 7-8, 10±1 spots in 8-9 and 2 ±1 spots in 9-10.
Qualitative comparisons of 2-DE protein patterns: The enlarged view of comparatively analysed spots and its expression in 3D view are represented in (Table 3) through the analysis of Image MasterTM 2D platinum 7.0 (GE healthcare). The results revealed that the majority of proteins were expressed at different level in both heads. Of 10 proteins (Spot 1-9, 16) were found to be a two fold higher expression in the larval head and 6 proteins (Spot 10-15) were two fold expression in the adult head. In addition, 6 proteins (Spot 17-22) were unique in the larval stage and whereas 8 were unique in the adult stage (Spot 23-30).
Mass spectrometric analysis of protein spots: The comparative and differentially expressed proteins from the head of larvae and adult were carefully analysed and subjected to mass spectrometry analysis. In total 30 proteins were identified from both larvae and adult heads using mass spectrometric approach and the results were shown on Table 4.
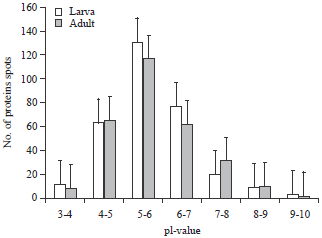 | |
| Fig. 3: | Distribution of proteins based on the isoelectric point in both larvae and adult head of Bombyx mori |
The list of comparatively identified proteins between the head of larvae and adult as follows cytoskeleton proteins (Spot 1), structural constituent and chitin based cuticle protein (Spots 2, 3, 6, 7), odorant binding proteins (Spot 4), three metabolic enzymes identified as characterized proteins such as glycerophosphodiester, enolase and monooxygenase/reductase (Spot 8, 10 and 13) and two uncharacterized proteins are also associated with enzymatic activity includes Phosphatase (Spot 9) and N-acetyltransferase activity (Spot 12), insect neuro hormone (Spot 11) and few uncharacterized proteins (5, 14, 15, 16) are associated with molecular binding activities.
Six proteins were identified uniquely in the larval head as follow: Actin-4 (Spot 17), apolipophorin-III (Spot 18), centromere-associated protein E like (Spot 19), iron-ion binding (Spot 20), cuticular protein RR2 motif 59 precursor (21) and one uncharacterized protein with phosphatase activity (Spot 22). Eight proteins from adult head were found to be unique namely, metabolic activity (23), uncharacterized protein with no function (Spot 24, 25, 29), glycoprotein hormone alpha2 (Spot 26), DNA binding activity, ALY protein (Spot 27, 30), clathrin coat assembly protein (Spot 28). In the present study, all spots had significant peptide hits but their pI/MW was not consistent with comparative theoretical and computational/values due to database limit and protein mass variation for homologous protein in different organisms. The identified proteins were classified based on the molecular, cellular and biological functions (Fig. 4).
| Table 3: | Selected 2D-PAGE gel areas, corresponding 3D profile of selected proteins identified in larvae and adult of B. mori |
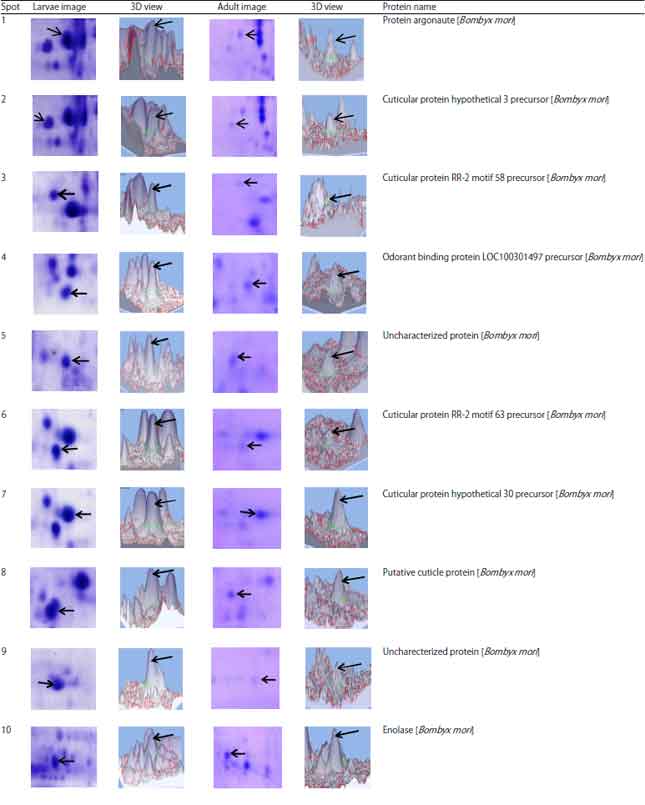  | |
| Spot volumes were generated from Image MasterTM 2D platinum 7.0 (GE Lifescience, USA). Protein identifications related to spots numbers are presented in this table | |
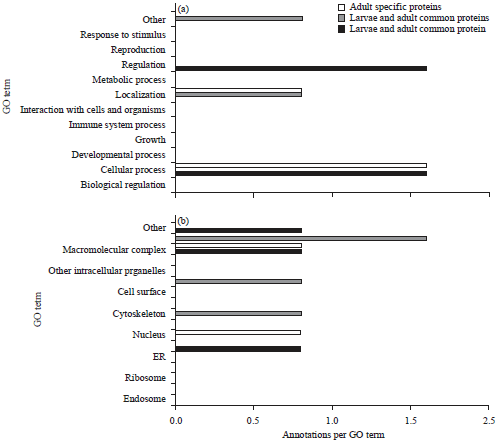 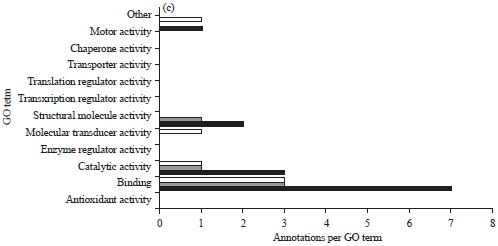 | |
| Fig. 4(a-c): | Gene Ontology (GO) categories of the common and unique expressed proteins from larval and adult heads of Bombyx mori, (a) Biological process (b) Cellular component and (c) Molecular function |
DISCUSSION
In the present study, the total protein isolated from larval as well as adult head of Bombyx mori was subjected to 2D-PAGE analysis. The 2-DE results clearly showed 315±3 protein spots from adult head and 296±3 spots in the larval head of B. mori. Most of the protein spots in the larval and adult head are observed in the pI range of 4-10 and 3.5-8, respectively. Like other kinds of organs/tissues of B. mori exhibited a huge variation in the protein spots through proteomics analysis. For instance, 400 protein spots were identified in silk glands12, 128 spots in hemolymph9, 566 spots in male midgut and 547 spots in female midgut24. The results of the present study suggests that enormous numbers of protein are synthesized during the larval stage (active feeding stage) for the growth and development whereas in adult showed less number of protein in the narrow range of pI due to changes in the adult feeding trait and non-synthesizing of major proteins during the adult development other than the ecolsion and reproduction.
When we carefully analysed the protein spots of larvae and adult head, a total of 30 similar and dissimilar proteins were observed, out of this results in more than 99% contrast with the study of Li et al.18. However, these proteins were individually observed from different tissues of B. mori for example collateral glands8, silk glands, haemolymph and fat body10,12, midgut23 and newly hatched larva30.
Interestingly, we observed that the expression of Odorant Binding Proteins (OBPs), apolipophorin III precursor (Apo-Lp III) and iron-ion transport protein (Spot 4, 18, 20) were higher in the larval head than the adult head. This result indicates that larval feeding activity requires OBPs, Apo-Lp III and iron-ion transport protein. In the larval stage, the OBPs help to transport of hydrophobic odorant from mulberry leaves to olfactory receptors2,36,37, Apo-Lp III and iron-ion transport protein may have the role to provide energy for feeding of larva. However, Apo-Lp III transports of diacylglycerol (DAG) from the fat body lipid storage deport to flight muscles in the adult38,39 and Iron-Ion transport protein is regulated to electron transport, neurotransmitter and synaptic-vesicle exocytosis40.
The RNA silencing process in B. mori is mediated by argonaute protein41. In the present study, the argonaute protein (Spot 1) expression was found to be two fold higher in the larval head when compared to the adult head, which reveal that agronaute protein occupy the major role to regulate the translation of proteins during larval developmental stage42,43. In addition, we identified the Centromere-associated protein-E (CENP-E) (Spot 19) from the larval head. The CEBP-E may conduct to mitotic kinesin, required for efficient and stable microtubule capture at kinetochores. The CEBP-E also directly binds to a kinetochore-associated kinase (BubR1) implicated in the mitotic checkpoint, the major cell cycle control pathway in which unattached kinetochores prevent anaphase onset44.
Most of the cuticular proteins are synthesized during the larval stage by dermal cells and secreted to its surface after each moulting in B. mori30. Similarly the foremost expressions of cuticular proteins (Spot 2- 3, 6-7, 21) were discovered in the larval and adult head of our study. In addition, the abundance of Cuticular protein RR-2 motif 59 precursors (Spot 21) and the uncharacterized protein (Spot 22) which are found to be involved in Cuticular development and metabolic process in the larval stage only.
| Table 4: | List of identified proteins from larval and adult heads of silkworm, B. mori |
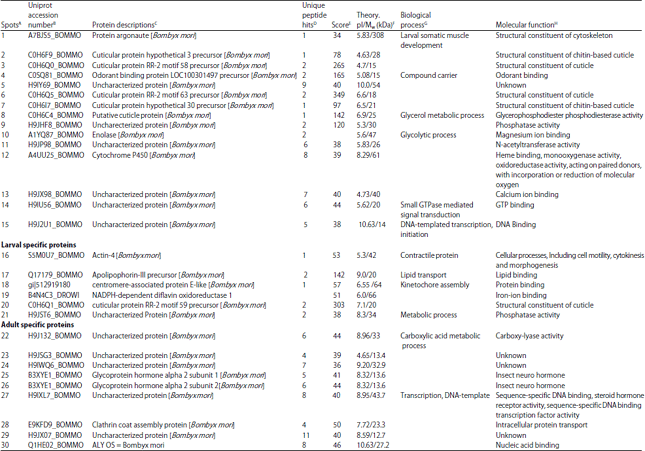 | |
ASpot ID is the symbol for every identified protein, which also noted on the 2-DE patterns (Fig. 1),BAccession number is the protein identifier in the NCBI or uniprot database (http://www.ncbi.nlm.nih.gov/), CSpecies indicate the organism that the matched proteins identified from NCBI or uniprot database, DMatched peptides are the number of paring experimental spectrums to a known protein and protein score is the results from mascot searching and is the main parameter for identification confidence,EScore value, FThe theoretical pI/MW represents the values of isoelectric point (pI) and Molecular Weight (MW) retrieved from protein databases of NCBI, GBiological functional annotation of the identified success protein and HMolecular function of the identified success proteins in each heads | |
These cuticular proteins are found to be playing a significant role in protecting the insect’s body from dehydration, physical injury, infection and penetration of environmental factors45-47,18. The actin-4 was identified in spot 17 of our 2-DE of larval head, which is playing an important role in Synaptogenesis, for example cell motility, cytokinesis and morphogenesis48-52.
In total, five enzymes were identified in the study. Of these, three characterized enzymes are as follows: Enolase, cytochrome P450 and glycerol phosphodiesterase and the other two are uncharacterized enzymes. Enolase is a glycolytic enzyme (spot 10), involved in the regulation and control of transcription, apoptosis and cell differentiation53 and feeding activity of the larva54,55. The cytochrome P450 enzyme (Spot 8) involved metabolism/detoxification of drugs, environmental pollutants and pesticides18 and glycerolphosphodiesterase (Spot 13) for glycerol metabolism7. The other two uncharacterized enzymes are involved in phosphatase and N-acetyl transferase activity (Spot 9 and 12) in both larvae and adult heads. Similarly, the N-acetyl transferase activity found in the silkworm head and presumed the function as a neurochemical mediator of photoperiodic control and event such as moulting and ecolsion56,57.
In the present study, expression of a glyco-protein hormone alpha 2 subunit observed as heterodimer in the head of the larva (Spot 11) and monomer in the adult head (Spot 26). It is evidenced to the study of Sudo et al.58 that the glyco-protein hormone is an insect neurohormone, it is a heterodimer and participate in developmental regulation includes cuticle formation, testis and ovary differentiation in B. mori. Also this glyco-protein hormone which regulates ion transport in hindgut of adult Aedes aegypti59. Further a unique expression of adult specific uncharacterized protein (Spot 23) which is involved in carboxlic acid metabolic process observed in the present study.
CONCLUSION
We speculate that all these adult unique protein and other highly expressed proteins may have a potential role in adult development and reproduction. The uncharacterized proteins (Spot 5, 24, 25, 29) from adult head observed with no functions. Further study need to be carried out for better understanding of the molecular network involves in biological functions of Bombyx mori. This study might lay a strong foundation to understand the feeding behaviour of larvae and adult development in B. mori and also in other lepidopteron insect pests.
ACKNOWLEDGMENT
We thank the Central Silk Board, Bangalore, India (CSB-31/2(PROJECT)/2010-11-RCS dated 16.09.2011) for financial support. V. Arun Prasanna is grateful to the Central Silk Board, Bangalore, India for the award of Junior Research Fellowship. We also thank to Dr. L. Uma, Director, National Facility for Marine Cyanobacteria (NFMC), Bharathidasan University, Tiruchirappalli, India for permitting to analyse the 2D-PAGE image. We sincerely thank Mr. A Jaleel, Mass Spectrometry and Proteomics Core Facility, Rajiv Gandhi Centre for Biotechnology, Thiruvananthapuram-695 014, Kerala, India.
REFERENCES
- Krieger, J., E. Grosse-Wilde, T. Gohl and H. Breer, 2005. Candidate pheromone receptors of the silkmoth Bombyx mori. Eur. J. Neurosci., 21: 2167-2176.
CrossRefPubMedDirect Link - Pelosi, P., I. Iovinella, A. Felicioli and F.R. Dani, 2014. Soluble proteins of chemical communication: An overview across arthropods. Front Physiol., 5: 320-320.
CrossRefDirect Link - Tanaka, K., Y. Uda, Y. Ono, T. Nakagawa, M. Suwa, R. Yamaoka and K. Touhara, 2009. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol., 19: 881-890.
CrossRefDirect Link - Xia, Q., D. Cheng, J. Duan, G. Wang and T. Cheng et al., 2007. Microarray-based gene expression profiles in multiple tissues of the domesticated silkworm, Bombyx mori. Genome Biol., Vol. 8.
CrossRefDirect Link - Pandey, A. and M. Mann, 2000. Proteomics to study genes and genomes. Nature, 405: 837-846.
CrossRefDirect Link - Gorg, A., W. Weiss and M.J. Dunn, 2004. Current two-dimensional electrophoresis technology for proteomics. Proteomics, 4: 3665-3685.
CrossRefPubMedDirect Link - Li, A. and D.L. Denlinger, 2009. Pupal cuticle protein is abundant during early adult diapause in the mosquito Culex pipiens. J. Med. Entomol., 46: 1382-1386.
CrossRefDirect Link - Jin, Y.X., Y.Y. Chen, Y.H. Jiang and M.K. Xu, 2006. Proteome analysis of the silkworm (Bombyx mori L.) colleterial gland during different development stages. Arch. Insect Biochem. physiol., 61: 42-50.
CrossRefDirect Link - Zhou, Z.H., H.J. Yang, M. Chen, C.F. Lou and Y.Z. Zhang et al., 2008. Comparative proteomic analysis between the domesticated silkworm (Bombyx mori) reared on fresh mulberry leaves and on artificial diet. J. Proteome Res., 7: 5103-5111.
CrossRefDirect Link - Hou, Y., Y. Zou, F. Wang, J. Gong, X. Zhong, Q. Xia and P. Zhao, 2010. Comparative analysis of proteome maps of silkworm hemolymph during different developmental stages. Proteome Sci., Vol. 8.
CrossRefDirect Link - Krishnan, M. and S. Konig, 2011. Progress of proteomic analysis in silkworm Bombyx mori. Biomacromol. Mass Spectrom., 2: 179-188.
Direct Link - Zhang, P., Y. Aso, K. Yamamoto, Y. Banno and Y. Wang et al., 2006. Proteome analysis of silk gland proteins from the silkworm, Bombyx mori. Proteomics, 6: 2586-2599.
CrossRefDirect Link - Moghaddam, S.H.H., X. Du, J. Li, J. Cao, B. Zhong and Y.Y. Chen, 2008. Proteome analysis on differentially expressed proteins of the fat body of two silkworm breeds, Bombyx mori, exposed to heat shock exposure. Biotechnol. Bioprocess Eng., 13: 624-631.
CrossRefDirect Link - Yang, H., Z. Zhou, H. Zhang, M. Chen, J. Li, Y. Ma and B. Zhong, 2010. Shotgun proteomic analysis of the fat body during metamorphosis of domesticated silkworm (Bombyx mori). Amino acids, 38: 1333-1342.
CrossRefDirect Link - Yi, Q., P. Zhao, X. Wang, Y. Zou and X. Zhong et al., 2013. Shotgun proteomic analysis of the Bombyx mori anterior silk gland: An insight into the biosynthetic fiber spinning process. Proteomics, 13: 2657-2663.
CrossRefDirect Link - Wu, X.F., X.H. Li, W.F. Yue, B. Roy and G.L. Li et al., 2009. Proteomic identification of the silkworm (Bombyx mori L) prothoracic glands during the fifth instar stage. Biosci. Rep., 29: 121-129.
CrossRefPubMedDirect Link - Li, J.Y., X. Chen, W. Fan, S.H.H. Moghaddam and M. Chen et al., 2009. Proteomic and bioinformatic analysis on endocrine organs of domesticated silkworm, Bombyx mori L. for a comprehensive understanding of their roles and relations. J. Proteome Res., 8: 2620-2632.
CrossRefDirect Link - Li, J., S.H.H. Moghaddam, X. Chen, M. Chen and B. Zhong, 2010. Shotgun strategy-based proteome profiling analysis on the head of silkworm Bombyx mori. Amino Acids, 39: 751-761.
CrossRefDirect Link - Li, J.Y., S.H.H. Moghaddam, J. Chen, M. Chen and B.X. Zhong, 2010. Shotgun proteomic analysis on the embryos of silkworm Bombyx mori at the end of organogenesis. Insect Biochem. Mol. Biol., 40: 293-302.
CrossRefDirect Link - Fan, L., J. Lin, Y. Zhong and J. Liu, 2013. Shotgun proteomic analysis on the diapause and non-diapause eggs of domesticated silkworm Bombyx mori. PLoS ONE, Vol. 8.
CrossRefDirect Link - Fu, Q., P. Li, Y. Xu, S. Zhang and L. Jia et al., 2011. Proteomic analysis of larval integument, trachea and adult scale from the silkworm, Bombyx mori. Proteomics, 11: 3761-3767.
CrossRefDirect Link - Zhang, S., Y. Xu, Q. Fu, L. Jia, Z. Xiang and N. He, 2011. Proteomic analysis of larval midgut from the silkworm (Bombyx mori). Comp. Funct. Genom.
CrossRef - Hu, X., L. Chen, X. Xiang, R. Yang, S. Yu and X. Wu, 2012. Proteomic analysis of peritrophic membrane (PM) from the midgut of fifth-instar larvae, Bombyx mori. Mol. Biol. Rep., 39: 3427-3434.
CrossRefDirect Link - Qin, L., H. Shi, H. Xia, L. Chen, Q. Yao and K. Chen, 2014. Comparative proteomic analysis of midgut proteins from male and female Bombyx mori (Lepidoptera: Bombycidae). J. Insect Sci., Vol. 14.
CrossRefDirect Link - Kannan, M., V. Suryaaathmanathan, M. Saravanakumar, A. Jaleel, D. Romanelli, G. Tettamanti and M. Krishnan, 2016. Proteomic analysis of the silkworm midgut during larval-pupal transition. Invertebr. Survival J., 13: 191-204.
Direct Link - Chen, J.E., J.Y. Li, Z.Y. You, L.L. Liu and J.S. Liang et al., 2013. Proteome analysis of silkworm, Bombyx mori, larval gonads: Characterization of proteins involved in sexual dimorphism and gametogenesis. J. Proteome Res., 12: 2422-2438.
CrossRefDirect Link - Zhang, Y., Z. Dong, P. Gu, W. Zhang and D. Wang et al., 2014. Proteomics analysis of adult testis from Bombyx mori. Proteomics, 14: 2345-2349.
CrossRefDirect Link - Zhong, X.W., Y. Zou, S.P. Liu, Q.Y. Yi and C.M. Hu et al., 2013. Proteomic-based insight into Malpighian tubules of silkworm Bombyx mori. PLoS ONE, Vol. 8.
CrossRefDirect Link - Zhang, Y.L., R.Y. Xue, G.L. Cao, Y.X. Zhu, Z.H. Pan and C.L. Gong, 2013. Shotgun proteomic analysis of wing discs from the domesticated silkworm (Bombyx mori) during metamorphosis. Amino Acids., 45: 1231-1241.
CrossRefDirect Link - Liu, P., Y. Wang, X. Du, F. Shi and Z. Meng, 2014. A comparative proteomic analysis of parthenogenetic lines and amphigenetic lines of silkworm. Biotechnol. Bioprocess Eng., 19: 641-649.
CrossRefDirect Link - Nirmala, X., V. Vanishree and M. Krishnan, 1999. Changes in embryonic protein profile and economic characters of Bombyx mori (Lepidoptera:Bombycidae) following UV irradiation. Indian J. Exp. Biol., 37: 560-566.
PubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Rosenfeld, J., J. Capdevielle, J.C. Guillemot and P. Ferrara, 1992. In-gel digestion of proteins for internal sequence analysis after one- or two-dimensional gel electrophoresis. Anal. Biochem., 203: 173-179.
CrossRefDirect Link - Bhatia, V.N., D.H. Perlman, C.E. Costello and M.E. McComb, 2009. Software tool for researching annotations of proteins: Open-source protein annotation software with data visualization. Anal. Chem., 81: 9819-9823.
CrossRefDirect Link - Gong, Y., T.C.S. Pace, C. Castillo, C. Bohne, M.A. O'Neill and E. Plettner, 2009. Ligand-interaction kinetics of the pheromone- binding protein from the gypsy moth, L. dispar: Insights into the mechanism of binding and release. Chem. Biol., 16: 162-172.
CrossRefDirect Link - Harada, E., J. Nakagawa, T. Asano, M. Taoka and H. Sorimachi et al., 2012. Functional evolution of duplicated odorant-binding protein genes, Obp57d and Obp57e, in Drosophila. PLoS ONE, Vol. 7.
CrossRefDirect Link - Wang, J., B.D. Sykes and R.O. Ryan, 2002. Structural basis for the conformational adaptability of apolipophorin III, a helix-bundle exchangeable apolipoprotein. Proc. Natl. Acad. Sci. USA., 99: 1188-1193.
CrossRefDirect Link - Liu, Q.N., K.Z. Lin, L.N. Yang, L.S. Dai and L. Wang et al., 2015. Molecular characterization of an apolipophorin-III gene from the chinese oak silkworm, Antheraea pernyi (Lepidoptera: Saturniidae). Arch. Insect. Biochem. Physiol., 88: 155-167.
CrossRefDirect Link - Wang, G.H., L. Jiang, L. Zhu, T.C. Cheng, W.H. Niu, Y.F. Yan and Q.Y. Xia, 2013. Characterization of Argonaute family members in the silkworm, Bombyx mori. Insect Sci., 20: 78-91.
CrossRefDirect Link - Parker, J.S. and D. Barford, 2006. Argonaute: A scaffold for the function of short regulatory RNAs. Trends Biochem. Sci., 31: 622-630.
CrossRefDirect Link - Peters, L. and G. Meister, 2007. Argonaute proteins: Mediators of RNA silencing. Mol. Cell, 26: 611-623.
CrossRefDirect Link - Weaver, B.A.A., Z.Q. Bonday, F.R. Putkey, G.J.P.L. Kops, A.D. Silk and D.W. Cleveland, 2003. Centromere-associated protein-E is essential for the mammalian mitotic checkpoint to prevent aneuploidy due to single chromosome loss. J. Cell Biol., 162: 551-563.
CrossRefDirect Link - Andersen, S.O., P. Hojrup and P. Roepstorff, 1995. Insect cuticular proteins. Insect Biochem. Mol. Biol., 25: 153-176.
CrossRefDirect Link - Dittmer, N.T., G. Tetreau, X. Cao, H. Jiang, P. Wang and M.R. Kanost, 2015. Annotation and expression analysis of cuticular proteins from the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol., 62: 100-113.
CrossRefDirect Link - Futahashi, R., S. Okamoto, H. Kawasaki, Y.S. Zhong, M. Iwanaga, K. Mita and H. Fujiwara, 2008. Genome-wide identification of cuticular protein genes in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol., 38: 1138-1146.
CrossRefDirect Link - Hennessey, E.S., D.R. Drummond and J.C. Sparrow, 1993. Molecular genetics of actin function. Biochem. J., 291: 657-671.
Direct Link - Sandiford, S.L., Y. Dong, A. Pike, B.J. Blumberg, A.C. Bahia and G. Dimopoulos, 2015. Cytoplasmic actin is an extracellular insect immune factor which is secreted upon immune challenge and mediates phagocytosis and direct killing of bacteria and is a Plasmodium antagonist. PLoS Pathog., Vol. 11.
CrossRefDirect Link - Nogales, E., 2001. Structural insights into microtubule function. Annu. Rev. Biophys. Biomol. Struct., 30: 397-420.
CrossRefDirect Link - Cingolani, L.A. and Y. Goda, 2008. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci., 9: 344-356.
CrossRefDirect Link - Bosch, M., J. Castro, T. Saneyoshi, H. Matsuno, M. Sur and Y. Hayashi, 2014. Structural and molecular remodeling of dendritic spine substructures during long-term potentiation. Neuron, 82: 444-459.
CrossRefDirect Link - Gerlt, J.A., P.C. Babbitt and I. Rayment, 2005. Divergent evolution in the enolase superfamily: The interplay of mechanism and specificity. Arch. Biochem. Biophys., 433: 59-70.
CrossRefDirect Link - Li, Q., D. Wang, S. Lv and Y. Zhang, 2014. Comparative proteomics and expression analysis of five genes in Epicauta chinensis larvae from the first to fifth instar. PLoS ONE, Vol. 9.
CrossRef - El‐Shesheny, I., I. El‐Hawary, I. Mesbah and N. Killiny, 2016. Comparative proteomic analysis between fifth‐instar nymphs and adults of Asian citrus psyllid Diaphorina citri. Physiol. Entomol., 41: 162-184.
CrossRefDirect Link - Itoh, M.T., A. Hattori, T. Nomura, Y. Sumi and T. Suzuki, 1995. Melatonin and arylalkylamine N-acetyltransferase activity in the silkworm, Bombyx mori. Mol. Cell. Endocrinol., 115: 59-64.
CrossRefDirect Link - Mohamed, A.A.M., Q. Wang, J. Bembenek, N. Ichihara, S. Hiragaki, T. Suzuki and M. Takeda, 2014. N-acetyltransferase (nat) is a critical conjunct of photoperiodism between the circadian system and endocrine axis in Antheraea pernyi. PLoS ONE, Vol. 9.
CrossRefDirect Link - Sudo, S., Y. Kuwabara, J.I. Park, S.Y. Hsu and A.J.W. Hsueh, 2005. Heterodimeric fly glycoprotein hormone-α2 (GPA2) and glycoprotein hormone-β5 (GPB5) activate fly leucine-rich repeat-containing G protein-coupled receptor-1 (DLGR1) and stimulation of human thyrotropin receptors by chimeric fly GPA2 and human GPB5 Endocrinology, 146: 3596-3604.
CrossRefPubMedDirect Link - Paluzzi, J.P., M. Vanderveken and M.J. O'Donnell, 2014. The heterodimeric glycoprotein hormone, GPA2/GPB5, regulates ion transport across the hindgut of the adult mosquito, Aedes aegypti. PLoS ONE, Vol. 9.
CrossRef