
Dalia Abel-Moneim Kheirallah
Department of Zoology, Faculty of Science, Alexandria University, 21511 Alexandria, Egypt
Zeinab Ahmed El-Moaty
Department of Zoology, Faculty of Science, Alexandria University, 21511 Alexandria, Egypt
Doha Ahmed El-Gendy
Department of Zoology, Faculty of Science, Alexandria University, 21511 Alexandria, Egypt
Journal of Entomology
Year: 2016 | Volume: 13 | Issue: 3 | Page No.: 55-71







ABSTRACT
The present study aimed to evaluate the possible hazards resulting from cement dust factory established in Mariout region West of Alexandria on Mediterranean coast. Two uncultivated sites were chosen for sampling the coleopterous insect Trachyderma hispida. The first site is situated at Burg El-Arab city, which is the reference site and the second site was at El-Hammam city which is situated in the vicinity of a cement factory and considered as the polluted site. The study was planned to monitor the bioaccumulation of metal (copper (Cu), lead (Pb) cobalt (Co) zinc (Zn) cadmium (Cd) chromium (Cr)) in soil, leaf litter and soft tissues of T. hispida from the selected sites which revealed a significant increase in metals concentration in the polluted site (El-Hammam city) than in the less polluted site (Burg El-Arab city). Many alterations of the general architecture of the testis were pronounced. Determination of histopathological features revealed disruption and damage for the normal cellular organization. Shrunk of seminiferous follicles, rupture of cyst walls, decreased number of cysts per follicles and decreased number of sperms per cysts were observed. Ultrastructural changes in testes of insects illustrate spermatogenesis disruptions and implied numerous abnormalities in the nucleus and cytoplasm. So, T. hispida can serve as biomonitoring of trace metals in polluted air, which leads to identification of hazardous exposures before adverse effects appear as potential risk to humans.
PDF Abstract XML References Citation
Received: January 19, 2016;
Accepted: March 06, 2016;
Published: April 15, 2016
How to cite this article
Dalia Abel-Moneim Kheirallah, Zeinab Ahmed El-Moaty and Doha Ahmed El-Gendy, 2016. Impact of Cement Dust on the Testis of Tachyderma hispida (Forskal, 1775) (Coleoptra: Tenebrionidae), Inhabiting Mariout Region (Alexandria, Egypt). Journal of Entomology, 13: 55-71.
DOI: 10.3923/je.2016.55.71
URL: https://scialert.net/abstract/?doi=je.2016.55.71
DOI: 10.3923/je.2016.55.71
URL: https://scialert.net/abstract/?doi=je.2016.55.71
INTRODUCTION
Air pollutants affect animals much the same as they affect people (Jelaska et al., 2007) and lethal or damaging doses to animals depend upon animal’s size. An additional hazard to animals is chronic poisoning from ingesting forage contaminated by particular pollutants. Important in this connection are the heavy metals; arsenic, lead and molybdenum and compounds like organic emissions and fluorides (Jelaska et al., 2007). Biomonitoring involves the use of biological or molecular markers as indicators signaling events in organisms exposed to environmental chemicals. Therefore, biological monitoring (through analysis of cells, tissues or body fluid of exposed organisms) may lead to identification of potentially hazardous exposures before adverse effects appear (Heikens et al., 2001).
During the last decades histopathological examination was used for diagnostics of environmental pathology. Ultrastructure is a successful tool capable of revealing sensitive and selectivity even at the sublethal effects of pollutants (Au, 2004).
An important endpoint considered in toxicological studies is the alteration of testicular germ cells. Therefore analysis of testicular composition and/or processes provide the best general purpose means of detecting potential adverse effect on male fertility (Clegg and Zenick, 1989). This should provide rapid and highly sensitive indicators of toxicity especially wildlife. Several toxicants have been shown to damage or impair testicular epithelium or sperms.
Toxicants have also been shown to impair reproduction in insects. Apoptosis during spermatogenesis has been reported in Triatoma infestans (Hemiptera: Reduviidae) a blood sucking insect and vector of chagas disease after exposure heavy metals (Mello et al., 1995).
The ultrastructural pathology of cadmium on Aedes albopictus (Diptera: Culicidae) cells was studied by Braeckman and Raes (1999). After 24 h exposure, most nuclei were irregular due to deep indentation, they revealed severe chromatin clumping. The perinuclear cisternae were dilated and the outer membrane was packed with increased amount of ribosomes. In the cytoplasm, dilation of rough endoplasmic reticulum, increase of the lysosomal system, condensation and swelling of mitochondria were most conspicuous effects.
Tissue damage in the accessory gland of the reproductive system of male Drosophila melanogaster (Diptera: Drosophilidae) was evident in flies cultured on food mixed with 10.0 μL mL–1 concentration of effluent from chrome plating industry in the eastern part of India (Mukhopadhyay et al., 2003). The lesion included disorganized endoplasmic reticula with fewer ribosomes, golgi cisternae were scarce, with very few vesicles. The lumen of the gland showed fewer filamentous structures.
The ultrastructure changes in testes of the aquatic insect Sphaerodema urinator (Hemiptra: Belostomatidae) sampled from highly polluted location of Lake Mariout, Alexandria was studied by Kheirallah (2006) and indicated several spermatogenesis anomalies. This study aims to evaluate the ultrastructural changes in the testes of Trachyderma hispida due to pollution with cement dust. Data obtained from this study will be useful in diagnosis of heavy metal subtle and obverse effects in the ecological survey of polluted habitats.
MATERIALS AND METHODS
Studied insect: Preliminary collection of the studied insect from the studied sites showed that Trachyderma hispida is one of the dominant adult coleopterous insects. Trachyderma hispida belongs to Tenebrionide beetles (darkling beetles that enact in the dark) it is one of the most diverse group within Coleoptera (Fig. 1).
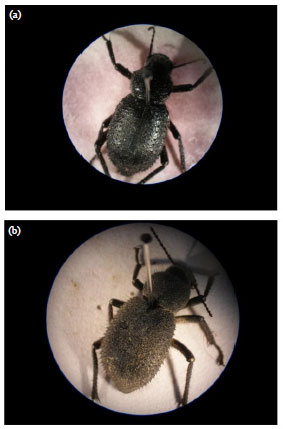 | |
| Fig. 1(a-b): | Photographs showing the studied insect collected from (a) Burg El-Arab (reference site) and (b) El-Hammam (cement dust site) (X2) |
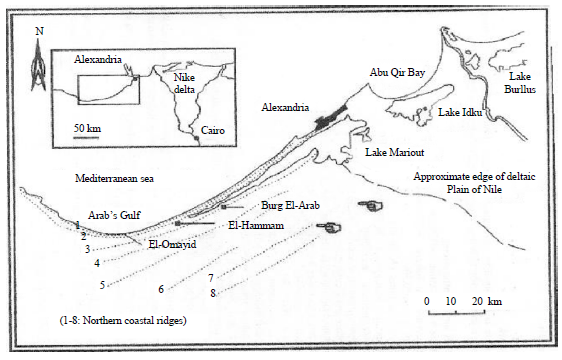 | |
| Fig. 2: | Map showing the sampling sites |
 | |
| Fig. 3: | Photograph showing the reference site (Burg El-Arab) and its shrubs |
Study areas: The present study was carried out in the Mariout region which is a strip of land about 100 km long and 30 km wide West of Alexandria on the Mediterranean cost, with an annual precipitation of about 150 mm decreasing sharply inland (Fig. 2). Two uncultivated sites were chosen for sampling the studied insects. These sites were (A) at Burg El-Arab city, which is 53 km West of Alexandria and 5 km South of the Mediterranean sea shore (Fig. 3) and (B) at El-Hammam city which is 62 km West of Alexandria and 10 km South of the sea shore (Fig. 4). The area surveyed at El-Hammam city is 1 km downwind from a recently built cement factory (Fig. 5). The area showed signs of cement dust deposition on the vegetation as well as soil surface.
Sampling procedure: Samples were collected randomly from each site. All insects collected were sexed then maintained alive in native soil and plants in suitable jars until processing. Simultaneously with insect collection, soil samples at a depth of 30 cm below the surface and leaf litter samples were collected from the specific sampling sites. Air dried and passed through 0.2 mm sieve to eliminate gravel and debris. Insects were sampled in two periods.
 | |
| Fig. 4: | Photograph showing the cement dust site (El-Hammam) and its shrubs |
 | |
| Fig. 5: | Photograph showing the location of the cement factory (El-Hammam) |
Soil and leaf litter analysis: The determination of heavy metals (copper (Cu), lead (Pb), chromium (Cr), zinc (Zn), cadmium (Cd), cobalt (Co) and nickel (Ni)) concentrations in sieved soil and leaf litter samples was carried out according to Loring and Rantala (1992) using atomic absorption spectrophotometer (Berkin-Elmer model 2380) under the recommended conditions and Detection Limits (DL) in the manual for each metal.
Analysis of heavy metal in the soft tissues of male T. hispida: The concentrations level (mg g–1) of seven metals Cu, Pb, Co, Zn, Cd, Cr and Ni were determined in the soft tissues of the insect by El-Sikaily et al. (2004).
Histological and ultrastructure investigations: For histological study, the standard method of dehydration, clearing and paraffin embedding was followed (Anderson and Gordon, 1996). Xylene was used as clearing agent. Tissues were embedded in paraffin wax (65-60°C). Testes sections (5 μm thick) were stained with freshly prepared hematoxylin and eosin (H and E).
For ultrastructural study, testes of adult male insects collected from the inspected sites were removed and fixed by immersing them immediately in 4F1G in phosphate buffer solution (pH 7.2) at 4°C for 3 h. Specimens were then postfixed in 2% OsO4 in the same buffer for 2 h. Samples were washed in the buffer and dehydrated at 4°C through a graded series of ethanol. Specimens were embedded in Epon-araldite mixture in labeled beam capsules. The LKB ultra microtome was used to obtain semithin sections (1 μm thick), these were mounted on glass slide and stained with toluidine blue then examined with the light microscope to determine the general structural features and to give an orientation. Ultrathin sections (60-70 μm thick) were cut from testes of males collected from the selected sites for TEM. These ultrathin sections were picked upon 200 mesh naked copper grids. Grids were double stained with uranyl for half an hour and lead citrate for 20-30 min.
Statistical analysis: Statistical analysis was performed using the program SPSS (Norusis, 2005). Data were analyzed statistically by using one way analysis of variance (ANOVA) (Sokal and Rohlf, 1981) to determine the difference between the two studied sites.
RESULTS
Concentration of heavy metals in soil and leaf litter samples: The data obtained from the analysis of soil and leaf litter samples collected from the inspected sites are illustrated in Fig. 6 and Table 1. The studied sites appear to be rather dissimilar with respect to the concentration of the seven studied heavy metals (Cu, Pb, Co, Zn, Cd, Cr and Ni). Statistical analysis for the mean differences of each metal revealed that six means differences out of seven were proven to be highly significant. It is of considerable interest that no difference in levels of Cr in soil was detected in the two locations, however, the analyzed leaf litters were observed to contain high magnitudes of Cr in the cement dust site.
Bioaccumulation of heavy metals in the soft tissue of male insects: It is apparent from Fig. 7 and Table 2 that male insects collected from the cement dust site have significant higher levels of most heavy metals in their soft tissues compared to male insects collected from the reference site.
Macroscopic and microscopic observations
Macroscopic findings: The testis was biconvex and its periphery is obviously thin. It looked like small transparent sacs filled with globular complexes or germ cells. A layer of connective tissue sheath (peritoneal sheath) is supported on a cup-shaped calyx. The calyx is continued behind into the vas deferens.
| Table 1: | Metal concentrations (mg g–1) in the soil and leaf litter collected from reference and cement dust sites |
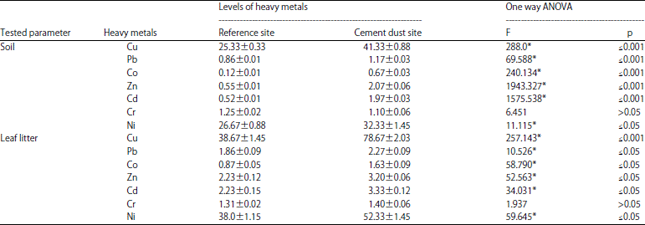 | |
| Data was expressed as Mean±SEM, F: Test (ANOVA), *Statistically significant at p≤0.05 | |
| Table 2: | Metal concentrations (mg g–1) in the soft tissues of males Tachyderma hispida collected from reference and cement dust sites |
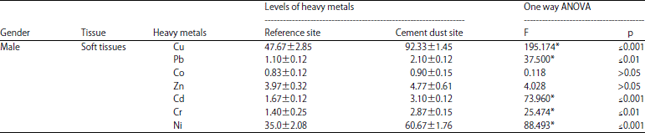 | |
| Data was expressed as Mean±SEM, F: Test (ANOVA), *Statistically significant at p≤0.05 | |
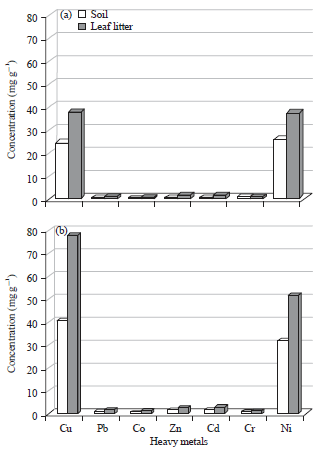 | |
| Fig. 6(a-b): | Metal concentrations (mg g–1) in the soil and leaf litter samples collected from (a) Reference site and (b) Cement dust sites |
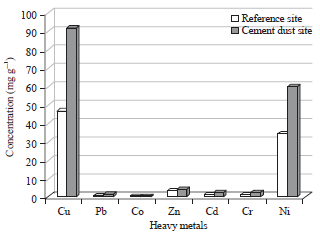 | |
| Fig. 7: | Metal concentrations (mg g–1) in the soft tissues of males T. hispida sampled from both reference and cement dust sites |
The vas deferens proceeded posteriorly and dilated at its end to form the vesicula seminalis (Fig. 8a). Males of T. hispida, collected from cement dust site exhibited deformations in shape and marked decrease in size of testis (Fig. 8b) as compared with those collected from the reference site.
 | |
| Fig. 8(a-b): | Photographs of the testis of T. hispida collected from the selected sites showing changes occurred in the shape and size, Testis collected from (a) Reference site (X12) and (b) Cement dust site showing reduction of the testicular size and dilation of the vas deference (X12) |
Microscopic findings: There are no previous documented studies described the testis structure (histologically or ultrastructurally) of the selected species T. hispida, therefore, the following description is considered the first report for such species.
Light microscopy: The histological appearance of testis of insects caught from reference site (Burg El-Arab) served as reference tissues for the diagnosis of testis aberration at the cellular and sub-cellular levels. Light microscopic preparation showed that the testis is composed of follicles encased by peritoneal sheath, wrapped together by connective tissue and tracheae (Fig. 9a). Follicle contained a number of cysts. Each cyst contained a clone of different stages of germinal and sertoli cells and held together by a connective tissue (Fig. 9b).
Figure 9c shows the different stages of the spermatogensis; the spermatogonia had large spherical nuclei with a reduced cytoplasmic area.
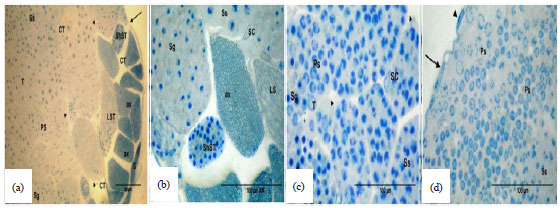 | |
| Fig. 9(a-d): | Semithin sections showing testicular follicles that are held together by Connective Tissue (CT) and Trachea (T). Peritoneal sheath (arrow), spermatogonia (Sg), primary spermatocytes (Ps), secondary spermatocytes (Ss), Long spermatids (Lst), short spermatids (ShST), Long Spermatozoa (LS), short spermatozoa (ShS), axoneme (ax), Sertoli Cells (SC) (specimens fixed in 4F1G and stained with toluidine blue) |
Spermatocytes appeared larger than spermatogonia with different sized nuclei and an increase in the size of cytoplasmic areas. These areas could be distinguished into primary and secondary spermatocytes (Fig. 9b and d). Spermatids contain densely stained nuclei, occupying most of the cell (Fig. 9a). Two developmental stages of spermatids; short (Fig. 9b) and long (Fig. 9a) are obviously distinguished. Nearly mature spermatozoa had completely condensed nuclei (Fig. 9a and b).
Both short and long sperms were observed (Fig. 9a and b, respectively) and the axons of spermatocytes are arranged in condensed bundles (Fig. 9a and b). Moreover, on some stages of spermatogenesis, it could be noted from these figures that sertoli cells lied peripherally in each group of germ cells (Fig. 9a, b).
In the present study, the influence of cement dust pollution on the morphology of testes of T. hispida collected from the cement dust site (El-Hammam) revealed various structural abnormalities of the follicles, loss of contact between the cysts and the follicular barrier (Fig. 10a), exfoliation and fibrosis of epithelial (Fig. 10b and c), one of the most important changes in some follicles of this group was the loss of germ cells which were occasionally observed (Fig. 10d). Also vacuolation of the follicles has been noticed (Fig. 10e). Necrosis of short sperms and fibrosis of interfollicular connective tissue were also observed (Fig. 10f). Necrotic bodies could occasionally be observed, necrosis of the sperms and rupture of cyst wall (Fig. 10f and g). In addition, hypertrophied sertoli cells were occasionally observed (Fig. 10g).
Electron microscopy: In the electron micrographs, the spermatogonia of the reference group appeared with a large nucleus, spherical and central in position with clear nuclear envelope. It appeared euchromatic with dense chromatin, homogenous in appearance and contained one large dense nucleoli (Fig. 11a). The cytoplasm contained mitochondria with variable shapes and sizes. They lied close together on one side of the nucleus forming a more or less dense mass. Within the cytoplasm free ribosomes were also observed.
The nucleus of each primary spermatocyte contained clumps of condensed heterogeneous chromatin appeared as synaptonemal complexes. At the beginning of the first division of maturation, they were found without nucleoli (Fig. 11b). It is obvious that nucleus appeared with a distinct and regular nuclear envelope. A dense mass of mitochondria was found adjacent to the nuclear membrane and golgi complex as well as free ribosomes appeared throughout the cytoplasm. Figure 11c shows that secondary spermatocytes exhibited a different appearance of their chromatin, which was more compact than in the primary spermatocytes. No synaptonemal complex was observed.
A second division of maturation occured in the secondary spermatocytes giving rise to spermatids. Two kinds of spermatids with different nuclear volumes were already observed. Judging from the location of both acrosome and centriole in these sections, the nuclear diameter of the large spermatids was larger than that of the small spermatids (Fig. 11d and e).
Early spermatids were distinguished by rounded nuclei. Nuclei of these spermatids appeared euchromatic and spherical in shape. In this stage, centriole was firstly recognized clearly attached to the posterior nuclear membrane. The centriole acts as a basal body that grows a flagellar axoneme.
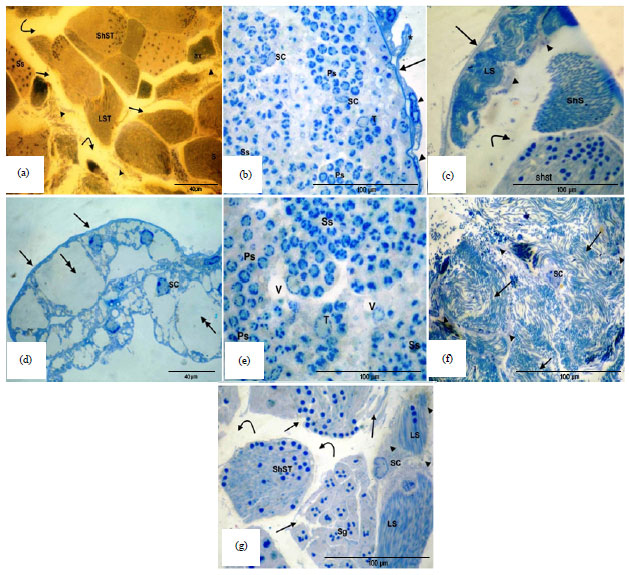 | |
| Fig. 10(a-g): | Semithin sections showing alterations in testicular follicles, (a) Shrinkage follicles, loss of contact between the cyst and follicular barrier (curved arrow), fibrosis of interfollicular connective tissue (arrow head) and rupture of cyst wall (arrows), (b) Exfoliation and fibrosis of epithelial wall (arrows), expanding in the flat nucleus of the epithelial sheath (arrow head), convolution of epithelial sheath (*) and hypertrophied Sertoli Cell (SC), (c) Fibrosis of follicular wall (arrow), fibrosis of interfollicular connective tissue (arrow head), loss of contact between the cysts (curved arrow), necrosis of Long Sperms (LS) and hypertrophied Sertoli Cell (SC), (d) Loss of germ cell (double head arrows), peritoneal sheath (arrows) and hypertrophied Sertoli Cell (SC), (e) Vacuolation (V) of the follicles, (f) Necrotic sperms (arrows), necrotic bodies (arrow heads) and hypertrophied Sertoli Cell (SC) and (g) Hypertrophied Sertoli Cell (SC), rupture of cyst wall (arrows), loss of contact between the cysts (curved arrow), fibrotic interfollicular region (arrow heads), necrosis of short spermatids (ShST) and Long Sperms (LS) (Specimens fixed in 4F1G and stained with toluidine blue) |
Early spermatids were recognized by roundish Nebenkern which formed by aggregated and fused mitochondria. Golgi bodies were located in the area between the nucleus and the Nebenkern. Also the cytoplasm contained rough endoplasmic reticulum and free ribosomes (Fig. 11d). The oldest spermatides were characterized by elongated nucleus, distinct nuclear envelope and conical acrosome cover the anterior of the nucleus in the opposite site of the centriole and the axoneme (Fig. 11e). In this stage Nebenkern underwent a structural differentiation and then divided into two separate mitochondrial derivatives toward the posterior end. Mitochondrial derivatives further extended to the posterior end and had cristae regularly arranged perpendicular to the axoneme (Fig. 11f). Centeriole or basal body in the insect spermatid is typically surrounded by a very dense granular adjunct called centriole adjunct. This is flanked by microtubules, as it elongates to form a rod in the tail. The rod is the only remains of the adjunct in the mature sperm (Fig. 11g).
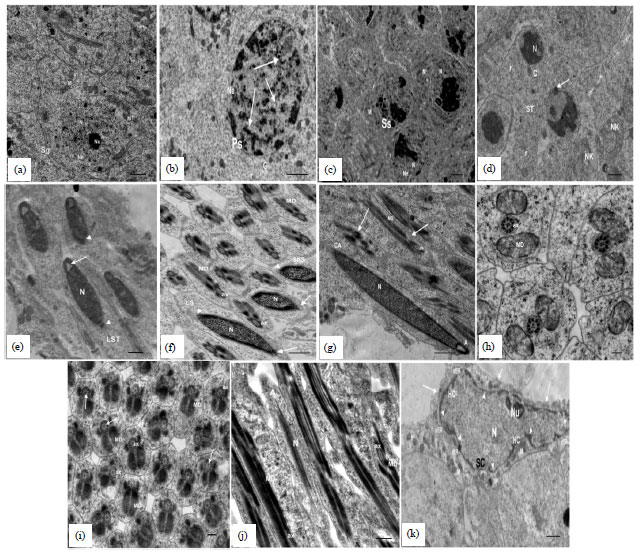 | |
| Fig. 11(a-k): | TEM: (a) Revealing spermatogonium (Sg) with large euchromatic nucleus (N) having electron dense nucleolus (Nu) and distinct Nuclear envelope (Ne), Mitochondria (M) lie on one side of the nucleus, (b-c) Primary spermatocytes (Ps) with nucleus containing synaptonemal complex (arrows), regular nuclear envelope (Ne), (d) Early spermatides (ST) with round large nucleus (N), neberkern (NK), Centeriole (C), arrows indicates centriole adjunct, (e) Long nuclear Spermatid (LST) with an acrosome (arrow) and centriole adjunct accumulating along posterior nucleus membrane (arrow head), (f) Long Sperm (LS), short sperm (ShS), transverse section of middle piece (arrows) showing: Axoneme (ax) with 9+9+2 arrangement of microtubules and Mitochondrial Derivatives (MD), Late Spermatids (ST) with large Nucleus (N) (g, h) Centriole Adjunct (CA) in a latero-posterior position to the nucleus (A) acrosome at tip of nucleus, (i) Arrows indicates the connection between the intertubulars and doublets, longitudinal section through nearly mature sperm(S) in their middle pieces and heads and (j) Sertoli Cell (SC) with Nucleus (N) contains few patches of heterochromatin (HC), large electron dense nucleolus (Nu), elongated and spherical mitochondria (M) with electron dense matrices and plasma membrane (arrows), rER: Small cisternae of rough endoplasmic reticulum, sER: Smooth endoplasmic reticulum, arrows heads indicate indentation of nuclear envelope. In all preparations; r: Free ribosomes and G: Golgi complex. Scale bar = 0.5 μm in a-k, scale bar = 0.1 μm in h, i (specimens fixed in 4F1G, post-fixed in OsO4 and stained with uranyl acetate-lead citrate) |
Most of sperm tail consisted of a flagellar axoneme and two mitochondrial derivatives. In transverse sections through the flagellum or middle-pieces, nine accessory tubules in a circle were observed around the doublets. Accordingly the axoneme was described as having the familiar 9+9+2 array. The arrays consisted of nine accessory tubules, nine microtubular doublets and two central microtubules (Fig. 11h). The developed nine accessory tubules lie in a circle outside and concentric with that of microtubular doublets. Initially some granular materials were seen near to the two mitochondrial derivatives (Fig. 11h).
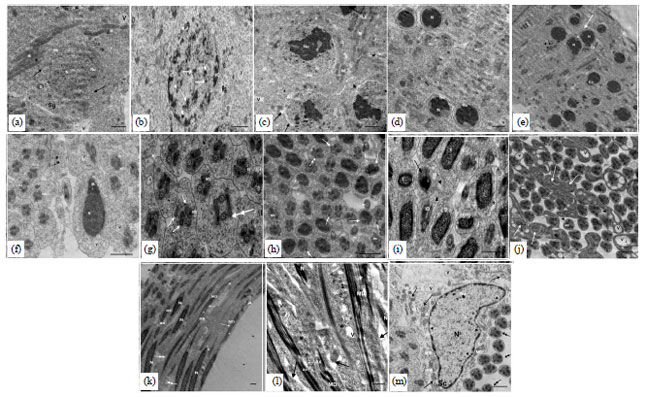 | |
| Fig. 12(a-m): | TEM illustrating ultrastructure changes in spermatogenic elements, (a) Deformed spermatogonia (Sg) with Nucleus (N), Nucleolus (Nu), Ne: Nuclear envelope, M: Mitochondria, arrows indicate blebs in nuclear envelope, (b) Morphologically altered Primary Spermatocytes (Ps), Hetero-Chromatin (HC), large chromatin granules (arrows heads), arrows indicates synaptonemal complex, Nuclear envelope (Ne), (c) Secondary spermatocytes (Ss) in meiotic division metaphase II; nuclear envelope (Ne), arrows indicate the blebs of nuclear envelope, lysis of cytoplasm in different region (arrows heads), C: Centriole, (d) Early malformed spermatids; nucleus (N), Nuclear Envelope (Ne), MD: Mitochondrial derivatives, (e) Malformed early spermatids with nucleus (N), double head spermatids (arrow), centriole adjunct (Ca), (f) Late spermatids, long nucleus (N), centriole adjunct (Ca), A: Acrosome, transverse section of middle piece with Mitochondrial Derivatives (MD) and axoneme (ax), (g) Cross section in middle piece of spermatids, double tail spermatids (double head arrows), rupture of plasma membrane (*), arrows indicate malformed mitochondrial derivatives, (h) Transverse section in late spermatids illustrating middle pieces and heads; cells without axoneme (*), MD: Degenerated mitochondrial derivatives, arrows heads indicate to some cell without MD, shrunken head of large nuclear spermatids (arrows), double head arrows indicate to accessory body, (i) Transverse section of both long (LSt) and short (ShSt) spermattides and middle piece, Note: Shrunken head (arrow) and lysis of plasma membrane (arrows heads) and cytoplasm (*), (j) Cross section in middle piece of spermatids with malformed middle piece and degenerated Mitochondrial Derivatives (MD), agglutination of spermatids in middle piece (arrows), arrows heads indicate rupture of plasma membrane, V: Vacuolation of the middle piece, (k) Longitudinal section of mature Sperm (S) with large Nucleus (N), Note: Convoluted plasma membrane (double head arrows), arrows indicate disintegrated Mitochondrial Derivatives (MD), degenerated axoneme (*), (l) Longitudinal section through nearly mature sperm illustrating abnormal middle piece, Note: degenerated axoneme (ax) and disintegrated Mitochondrial Derivatives (MD), arrows indicate convoluted plasma membrane, agglutination of sperms in some region (*) and (m) Hypertrophied sertoli cell, arrows heads indicate indentation of nuclear envelope, nucleus (N), nucleolus (Nu), lysosomes (ly), arrows indicate large amount of cell debris, M: Disintegrated mitochondria, rER: Short cisternae of rough endoplasmic reticulum, sER: Smooth endoplasmic reticulum, double head arrows indicate sperms in their middle piece. In all preparations, N: Nucleus, Nu: Nucleolus M: Mitochondria, A: Acrosome, ax: Axoneme V: vacuoles, r: Free ribosomes and G: Golgi complex. Scale bar = 0.5 μm in a-m, scale bar = 0.1 μm in g. (Specimen fixed in 4F1G, post-fixed in OsO4 and stained with uranyl acetate-lead citrate) |
At later stages, they are transformed into two longitudinal rods that flank the axoneme which are called accessory bodies. It can be observed in Fig. 11i that there was a synapomorphic feature which was represented by connecting bridges between axonemal doublets microtubules and the mitochondrial derivatives. The bulk of the intertubular material was connected to the doublets but not to the accessory tubules. In mature spermatozoa, during chromatin condensation, the nucleus diminished in size and attained small rod shape between thread like acrosome and the long flagellum (Fig. 11j).
Electron microscope preparations illustrated sertoli cells (Fig. 11k) in the periphery of the follicles, which comprised the wall of the germinal cysts containing spermatogenic elements. The nucleus of the sertoli cell was large having a polymorphous shape with highly irregular nuclear envelope. The chromatin was evenly distributed and few patches chromatin were associated with the nuclear membrane. One or two large electron dense nucleoli were located peripherally or centrally in the nucleus. The cytoplasm contained elongated and spherical mitochondria and large number of free ribosomes. Rough and smooth endoplasmic reticulum (rER, sER) were also observed.
At the ultrastructural level, the testes of T. hispida collected from cement dust site appeared with signs of various rate of injury. The germ cells underwent necrosis and spermatogenesis was disturbed. In general, cysts containing spermatogonia appeared with abnormal profile. The changes included both the nucleus and cytoplasm. Nuclear deformations were more pronounced than cytoplasmic changes. Nuclear changes revealed severely dilations of nuclear envelope, formations of blebs in the nuclear envelope and disintegration of nucleolus. Also nuclei of some cells were greatly reduced in size (Fig. 12a). In the cytoplasm, vacuoles were obvious. Mitochondria appeared with electron dense matrices. In some cells the cytoplasm was reduced, while free ribosomes were increased (Fig. 12a).
Primary spermatocytes showed some degenerative changes. The nucleus appeared with discrete patches of abnormal chromatin condensation near the nuclear envelope, disorganized distribution of heterochromatin clumps and dilation of nuclear envelope. Mitochondria displayed lysis of their matrices (Fig. 12b).
Secondary spermatocytes were irregular in shape. The nuclear envelope showed dilation and had blebs. Vacuolation and lysis of cytoplasm were noticed. Mitochondria showed signs of disintegration, degeneration and disorganization (Fig. 12c).
Spermatids showed degenerated changes such winding nuclear envelope (Fig. 12d). Large nuclear spermatids had an oval nucleus, appeared with abnormal chromatin condensation, dilated nuclear envelope and double head spermatids can be observed (Fig. 12e). In the cytoplasm aggregation of mitochondria takes place to form the Nebenkern which appeared to be disintegrated. Lysis of cytoplasm was also noticed. At later stage of development necrotic and abnormal spermatids (large nuclear) were commonly observed and vacuoles were commonly seen in cytoplasm (Fig. 12f).
Malformed spermatids with double tail rupture of plasma membrane and agglutinated ones were commonly seen in some specimens (Fig. 12g). Sometimes spermatids without axoneme were appeared (Fig. 12h), shrunken head of spermatids were also observed (Fig. 12i). Middle-pieces appeared with degeneration of mitochondrial derivatives and rupture of plasma membrane (Fig. 12j). Vacuolated and agglutinated middle pieces were commonly noticed. Moreover, degeneration of axoneme and agglutinaed middle pieces was also observed as well (Fig. 12j).
Sperm morphology revealed a variable degree of deformation, such as convoluted plasma membrane and vacuolated cytoplasm (Fig. 12k). Degenerated axonemes, rupture of plasma membrane and disintegrated mitochondrial derivatives were frequently noted (Fig. 12k). Furthermore, agglutination of sperms at several regions was observed as well (Fig. 12l).
Changes in Sertoli cells were frequently observed. They were hypertrophied, displayed high phagocytic activity and expanded cytoplasm (Fig. 12m). The nucleus appeared with severe chromatolysis. Meanwhile, the nucleolus appeared highly electron dense and sometimes exhibited segregation of its components. Degenerative changes among these cells like disintegrated mitochondria and vacuolated cytoplasm were also observed. Cisternae of rough endoplasmic reticulum (rER) and dilated smooth endoplasmic reticulum (sER) were further found in the cytoplasm. Moreover, an increase in the amount of cellular debris and large number of lysosomes were also noticed.
DISCUSSION
Environmental contaminants are one of the most challenging problems that face the world today. The growing human population and movement towards global market have driven competition in industrial and technological development at a high speed.
One of the most important reasons which justify the need for a broad-scale, interdisciplinary assessment of different type of environmental contamination is cement dust pollution. There have been many reports on the occurrence of visible and subtle injuries induced by cement dust to humans, plants and animals (Fatima et al., 2001).
Bioaccumulation: Cement dust in the present study was found to contain several metals. Metals enter the body of insect via the integument or through the intake of contaminated food. Metals are thus brought into contact with the organs and tissues of insects and consequently accumulated to a different extent in different organs/tissues. It was found in this study that the heavy metal concentrations in the soil were significantly higher in the cement dust site than reference site. These findings were in agreement with those obtained by Poon and Chen (1999) who demonstrated that cement dust and its products incorporated a high quantity of lead. Leaf litter was found to contain significantly higher concentration of heavy metals in the cement dust than reference site. These data confirm those obtained by some previous studies such as, Fakhry (1994) who studied the effects of cement dust on plants present around the cement factory at El-Hammam area and she found a decrease in both richness and diversity in the most affected site, where only half the number of plant species could resist the disturbance of the cement factory.
The concentration of tested heavy metals in soft tissues of the male insect T. hispida sampled from the heavily polluted site (cement dust) displayed higher concentration of all tested heavy metals comparable to the less polluted reference site (Burg El-Arab). This gives an indication that T. hispida is incapable to regulate metals uptake. Body residues are often better estimates of a chemical at the site of toxic action in an organism than ambient concentrations. Metal concentrations can provide a more direct way to evaluate the biological impact of trace metals (Luoma, 1996).
The appreciable increase in the concentrations of trace metals in body of adult males T. hispida relative to soil concentrations could be due to irregulation of the internal metal concentrations. Insects have an efficient detoxification mechanism such as binding to protein or stored in granules. Darlington and Gower (1990) observed Cu containing granules in the cells of the Malpighian tubules and sub-cuticular region of larvae Plectrocnmia conspersa (Trichoptera: Polycentropodidae). These granules provide a mechanism for taking potentially toxic concentrations of Cu and possibly other metals out of circulation. Among the studied heavy metals, Cu and Ni are two metals occurring in higher levels in soil and leaf litter of the two studied sites as well as in body of the studied insect. Similarly, Heliovaara et al. (1987) showed that the insects, Aradus cinnamomeus (Heteroptera: Aradidae) living in the environment around the industrialised town of Harjavalta in Southwestern Finland accumulated heavy metals, particularly Cu and Ni. It has been found that Aradus cinnamomeus (Heteroptera: Aradidae), Petrova resinella (Lepidoptera: Tortricidae) and Chrysomela lapponica (Coleoptera: Chrysomelidae) living in habitats severely contaminated by heavy metals accumulate heavy metals, in particular Zvereva et al. (2003). Copper is one of the most toxic heavy metals (Nor, 1987). Excess Cu causes a variety of physiological and histological effects. Also Cu impairs growth. Nickel reduced respiratory rate and increased mortality in the insect Pterostichus oblongopunctatus (Coleoptera: Carabidae) (Bednarska and Laskowski, 2009).
Histological and ultrastructural studies: Although, numerous literatures have described the histopathological changes resulted from exposure of testis to different classes of pollutants invertebrate animals but these literatures were rare with respect of insects. The present study is considered among the few report describing changes in testis of insects collected from habitat contaminated with cement dust.
The present study showed that the testes of T. hispida sampled from reference site consisted of testicular follicles encased by peritoneal sheath, warped together by connective tissue and tracheae partitioned into a number of germinal cysts contained a clone of different stages of germinal and Sertoli cells which support the gamete producing cells. These results are in agreement with the results of Freitas et al. (2003) and Kheirallah (2006) worked on different species of hemipteran tripes and species.
Histologically the testis of insects sampled from cement dust site displayed collapsed and shrinkage of semineferous follicles. Similar observation was reported by Kheirallah (2006) worked on the testes of the aquatic insect Sphaerodema urinator (Hemiptera: Belostomatidae) sampled from moderately and high polluted location of lake Mariout, Alexandria.
Histopathological alterations in the form of convolution and/or fibrosis of follicular walls, thickening of interfollicular regions and rupture of cyst walls have also been observed in insects sampled from cement dust site. McGee et al. (1992) reported that changes in the cell surface morphology including irrigulation of membrane contour and focal thickening often occur at an early stage in the course of cell injury, since the cell membrane is the first point of contact with any toxic substance. The structure of the cell membrane often changes as an abnormal cell loses surface receptors and antigenic determinants or gain new ones. The receptors may become abnormally clumped in one part of the cell (Ritchie, 1990).
The observed disruption of germ cell differentiation in testes of insects sampled from the cement dust site might have been caused by an alteration in the endocrine system which relies largely on ecdysteroids and on terpenoids (juvenile hormones), which regulate moulting, differentiation, metamorphosis and involved in reproductive process such as ovulation and spermiogenesis (Segner et al., 2003).
Necrosis and degenerative spermatogenic elements were apparent in testes of insects sampled from cement dust site. Such decrease in the quality of gametes may in-turn impair the reproductive success of adult organisms. Similar results were obtained by Kheirallah (2006) worked on the aquatic insect S. urinator caught from a polluted location of Lake Mariout, Alexandria.
The present ultrastructural observation of testis of insects sampled from reference site revealed spermatogonia with large, central and homogenous nuclei. The cytoplasm contained mitochondria with variable shapes and sizes. They lied close together on one side of the nucleus forming a dense mass. In addition free ribosomes were also present in the cytoplasm of the spermatogonia. Similar ultrastructural criteria were observed in the aquatic insect S. urinator Kheirallah (2006).
In the present study, it was found that, the structure of primary and secondary spermatocytes showed distinctive changes. The nucleus appeared with discrete patches of abnormal chromatin condensation. The distinctive changes observed from spermatogonia to spermatocytes were as follow: Increased size, increased nuclear cytoplasmic ratio and the mitochondria became uniformly distributed in the cytoplasm. These results resemble those recorded by Moriber (1956) worked on the hemiptern insect Gerris sp.
In the present study, it was found that the structure of primary and secondary spermatocytes showed nuclei with thick clumps of chromatin. In parallel, Payne (1966) described similar feature in spermatocytes of the hemipteran insect Gelastocoris oculatus (Hemiptera: Gelastocoridae). It was always possible to distinguish secondary spermatocytes (usually short-lived) from longer-lived primary spermatocytes (Berry et al., 1992). It is worthy to mention that formation of a synaptonemal complex is one of the characteristics of primary spermatocytes (Pachytene stage) observed in our preparations. Similar ultrastructural feature was described in the stick insect Carausius morosus (Orthoptera: Phasmatodea) by Pijnacker and Ferwerda (1986).
Spermiogenesis in T. hispida showed several features that are typical of insects. In the present study, the development of the nucleus involves changes in shape and in degree of chromatin condensation. This is in agreement with Fernandes et al. (2001) worked on the hemipteran insects Euchistus heros (Hemiptera: Pentatomidae).
The acrosome is formed by golgi complex and presents unusual morphological features during its development. This concept is in general agreement with the conclusion of Fernandes et al. (2001) and Kheirallah (2006). The centriole in T. hispida appeared as a straight rod along the posterior nuclear membrane. In earlier studies, on the hemipteran insect Gelastocoris oculatus (Hemiptera: Gelastocoridae) Payne (1966) observed that the centrioles were first attain a V-shaped rods near the nuclear membrane then continued their movement until they occupied positions on opposite sides of the nucleus, at which time the V-shape changed to straight rod and divide into two equal parts.
An electron dense material, the centriole adjunct accumulated around the centriole along the posterior nuclear membrane. This structure may be involved in the microtubule organization during spermiogenesis as suggested by Fernandes and Bao (1998). Similar results have been reported by Kubo-Irie et al. (2003) worked on the hemipteran insect Graptosaltria nigrofuscata (Hemiptera: Auchenorrhyncha).
The present study revealed two kinds of sperm with different nuclear lengths. Similar results have been obtained in the hemipteran insect, Graptopsaltria nigrofuscata (Kubo-Irie et al., 2003).
In the present study, the spatial constructions of long and short sperm of T. hispida were basically similar to each other, but nuclear length and the total length were shorter in the short sperm than in the long sperm. This is in consistent with Jamieson et al. (1999) who reported a distinct difference in the nuclear volume of the two spermatids in hemipterans but acrosome, flagellar formation and nuclear condensation were similar to those occurring in typical spermiogenesis of other insects. Spermatozoa in T. hispida possess an axoneme array 9+9+2 in microtubule pattern and two mitochondrial derivatives. Mitochondria in early spermatids of many insects aggregate and form a round body (the Nebenkern). This body undergoes a structural differentiation, which divides into separate equal-sized bodies (Yue et al., 1999). In this structure body identity of the individual mitochondria is lost but their essential structure remained and acrosomal and flagellar organization had been initiated (Payne, 1966). This comes in full agreement with the findings in the present species T. hispida. No separate mitochondria were found after Nebenkern formation.
Ultrastructurally, the testis of the insects sampled from cement dust site showed many pathological lesions. In spermatogonial cells, rupture of plasma membrane and nuclear distortion were commonly observed in these germs cells. Similar changes resulting from metal toxicity in cell line of the dipteran Aedes albopictus (Diptera: Culicidae) was reported by Braeckman and Raes (1999) and Braeckman et al. (1999). Changes in the plasma membrane may be due to increased lipid peroxidation. Lipid peroxidation of the membranes has been considered for a long time to be the most plausible mechanism for the damage induced by free radicals (Freeman and Crapo, 1982). The appearance of distorted nuclei is noteworthy referring to the damaged of RNA metabolism (Bayomi et al., 1979). Appearance of blebs were also noticed as previously reported by Braeckman and Raes (1999), who noticed obvious blebbing in insect’s cell line A. albopictus exposed to methyl mercury (MeHg).
The present study demonstrated apoptotic nuclei and vesicles in the polluted groups. The appearance of these structures resembled heavy metal treatment in germ line cells of the hemipteran insect Triatoma infestans (Hemiptera: Reduviidae) (Mello et al., 1995). Moreover, disruption of mitochondria in the cytoplasm of beetles collected from cement dust site was observed. Pathological reactions of mitochondria to toxins were similarly observed in invertebrate intoxicated cells (Braeckman et al., 1999). Mitochondrial alterations reflect deregulation of mitochondrial membrane transport (Braeckman et al., 1999). In parallel, Au et al. (2003) showed that phenol disrupt cytoplasmic membranous structures (mitochondrial cristae, distortion of mitochondrial membranes and nucleus envelope) in sea urchin, Anthocidans crassipina (Camarodonta: Echinometridae). The presence of myelin figures indicated a degenerative process which is usually formed when hydrophilic phospholipids are released from degenerating membranes as suggested by Jones and Hunt (1983).
In the present study, it was found that the nuclei of primary spermatocytes exhibited synaptonemal complex with unpaired elements. The synaptonemal complex forms the axes of meiotic chromosomes during the synapsis of homologues and is composed of two lateral elements connected by transverse filaments and ladder-like central element (Moens et al., 1998). Two types of abnormalities were observed when studying the synaptonemal of male chromosomes; desynapsis and terminal union abnormalities were observed between identical and different synaptonemal complex. Kheirallah (2006) observed similar change in the hemipteran insect Sphaerodema urinator inhabiting polluted lakes (Maruit and Edku). Indeed, univalent formation has been suspected to be the outcome of chromosomal asynapsis, on account of reciprocal translocation (Acharya et al., 2003). Furthermore, the lysis of nuclei in primary spermatocytes is supposedly associated with a damage of some of the grossly affected chromosomes in their spermatogonial stages as reported by Eason and Turck (2002). The observed vacuolated lytic areas in primary and secondary spermatocytes is an evidence of cytotoxicity, the injuries might be a consequence of the release of lysosomal hydrolases into the cytoplasm by Vandenbulcke et al. (1998).
During the present study, ultrastructural diagnosis identified various sperm morphological abnormalities in the cement dust site. Incidences of structurally abnormal sperm shapes are reported to be genetically controlled by numerous autosomal and sex-linked genes (Palumbo et al., 1994). Thus, the obvious relative decrease in the density of chromatin condensation seen in sperms of insects from cement dust site might be associated with chromosomal abnormalities. Similar abnormalities in chromosomes in mitotic and meiotic cells of the orthopteran insect Tetrix tenuicornis (Orthoptera: Tetrigidae) due to heavy metal accumulation were recorded by Warchalowska-Sliwa et al. (2005).
In some affected specimens, the mid-piece of the sperm showed convoluted plasma membrane. Au et al. (2001) indicated that exposure to toxicants (Cd and phenol) changed the size and shape of the mid-piece of the spermatozoa of the mussel and sea urchin and this might affect the balance of spermatozoa in their swimming. They also observed disorganization of mitochondrial membranes and cristae, thus disrupt ATP supply for sperm movement.
During the present study, ultrastructural diagnosis identified mitochondrial abnormalities accompanied by axonemal degeneration in the polluted group. Similarly, Au et al. (2001) reported cytological distortion of sperm tail and mitochondrial cristae deformation in sperm cells of sea urchin chronically exposed to Cd. Furthermore, biflagellate sperms were observed in insects from the polluted site. Similarly, Kheirallah (2006) described sperm abnormalities in head and tail of the hemipteran insect Shaerodema urinator inhabiting polluted sites in lake Mariout and Edku.
Sperm axonemal alterations are known to be the primary cause of sperm immotility and it was also found that a reduction of axonemal components caused lowered motility (Hancock and de Kretser, 1992). Fuller (1993) reported that defects in spermatogenesis resulted in either complete absence of mature sperm or the presence of highly disorganized, non-motile sperm. Au et al. (2003) suggested that chronic exposure to phenol lower the quality of sperm and reproduction success in sea urchins, which may threaten their survival and demonstrated motility impairment and concomitant reduction of fertilization capability in sea urchin with dark tail and/or distorted mitochondria.
The present study demonstrated that sperm agglutinations were obvious in specimens collected from cement dust site. It seems likely that sperm agglutination results from the influence of autologous IgM, which comes to penetrate the efferent duct as autoimmune responses in the testis progress as suggested by Kheirallah (2006). Worthy to mention that, the micro-morphological changes in spermatozoa presented in the present study should be included as a model for predicting environmental hazards.
During the present ultrastructural study of sertoli cells from specimens collected from cement dust site revealed irregularity shaped nucleus, elongated and spherical mitochondria, well-developed system of sER, strands of rER and dispersed free ribosomes. Moreover, in some specimens sertoli cells were hypertrophied. On the basis of these cytoplasmic features, Buckland-Nicks and Chia (1986) showed evidence of an extensive intracellular digestion that is used effectively to phagocytosis the residual cytoplasm. Pathological changes in sertoli cells have also been documented in invertebrates.
Electron micrographs also showed large phagolysosomes filled with degenerating spermatozoa, cellular debris, vacuolization of the cytoplasm increase in number of lysosomes was noticed in these preparations. Similar observations were demonstrated in hemipteran insects inhabiting polluted sites of lake Mariout and Edku (Kheirallah, 2006).
Sertoli cell phagocytize germ cells that degenerate in the normal course of spermatogenesis (Van Vuren and Soley, 1990). Thus, the presence of multiple sertoli cells with distended cytoplasm filled with degenerate spermatozoa in specimens from the cement dust site might be due to an increase in necrosis or apoptosis in metal-exposed insects. In some other specimens, sertoli cells became irregular in shape with less dense euchromatic material and degenerated nucleoli. The endoplasmic reticulum and ribosomes became very few. Kheirallah (2006) reported similar results in the testes of the aquatic insect S. urinator sampled from moderately and high polluted location of lake Mariout, Alexandria. They suggested that deformed sertoli cells with degenerated cytoplasmic organelles reflected cessation of metabolic processes and malformation of these cells, which may lead to disturbance in spermatogenesis in the form of altered, deformed sperm structure and orientation.
It is obvious that germ cells and sertoli cells organelles were affected by pollution, which suggests that pollutants disturb cell function at more than one site.
CONCLUSION
The cement kilns are major source of toxic air emission in the industrial area. Insects may not be able to detoxify complex chemical compounds that they inhale or ingest with contaminated food. Residues of such persistent chemicals are laid down and transferred to a higher level in the food chain. This data revealed that insects are good in biomonitoring environmental pollution.
ACKNOWLEDGEMENT
The study was partially funded by Alexandria University. The authors are thankful to Zoology Department, Faculty of Science.
REFERENCES
- Jelaska, L.S., M. Blanusa, P. Durbesic and S.D. Jelaska, 2007. Heavy metal concentrations in ground beetles, leaf litter, and soil of a forest ecosystem. Ecotoxicol. Environ. Safety, 66: 74-81.
CrossRefDirect Link - Heikens, A., W.J.G.M. Peijnenburg and A.J. Hendriks, 2001. Bioaccumulation of heavy metals in terrestrial invertebrates. Environ. Pollut., 113: 385-393.
CrossRefDirect Link - Au, D.W.T., 2004. The application of histo-cytopathological biomarkers in marine pollution monitoring: A review. Mar. Pollut. Bull., 48: 817-834.
CrossRefDirect Link - Mello, M.L.S., F.S. Kubrusly, M.A.F. Randi, V.L.C.C. Rodrigues and A.N.F. Filho, 1995. Effect of heavy metals on chromatin supraorganization, nuclear phenotypes and survival of Triatoma infestans. Entomol. Exp. Applied, 74: 209-218.
CrossRefDirect Link - Braeckman, B., K. Brys, U. Rzeznik and H. Raes, 1999. Cadmium pathology in an insect cell line: Ultrastructural and biochemical effects. Tissue Cell, 31: 45-53.
CrossRefDirect Link - Mukhopadhyay, I., D.K. Saxena and D.K. Chowdhuri, 2003. Hazardous effects of effluent from the chrome plating industry: 70 KDa heat shock protein expression as a marker of cellular damage in transgenic Drosophila melanogaster (hsp70-lacZ). Environ. Heath Perspect., 111: 1926-1932.
PubMedDirect Link - Loring, D.H. and R.T.T. Rantala, 1992. Manual for the geochemical analyses of marine sediments and suspended particulate matter. Earth-Sci. Rev., 32: 235-283.
CrossRefDirect Link - El-Sikaily, A., A. Khaled and A. El Nemr, 2004. Heavy metals monitoring using bivalves from Mediterranean Sea and Red Sea. Environ. Monit. Assess., 98: 41-58.
CrossRefDirect Link - Sokal, R. and F. Rohlf, 1981. Biometry: The Principles and Practice of Statistics in Biological Research. 2nd Edn., WH Freeman, San Francisco, CA., USA., ISBN-13: 9780716712541, Pages: 859.
Direct Link - Fatima, S.K., P.A. Prabhavathi, P. Padmavathi and P.P. Reddy, 2001. Analysis of chromosomal aberrations in men occupationally exposed to cement dust. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 490: 179-186.
CrossRefDirect Link - Poon, C.S. and Z.Q. Chen, 1999. Comparison of the characteristics of flow-through and flow-around leaching tests of solidified heavy metal wastes. Chemosphere, 38: 663-680.
CrossRefDirect Link - Luoma, S.N., 1996. The developing framework of marine ecotoxicology: Pollutants as a variable in marine ecosystems? J. Exp. Mar. Boil. Ecol., 200: 29-55.
CrossRefDirect Link - Darlington, S.T. and A.M. Gower, 1990. Location of copper in larvae of Plectrocnemia conspersa (Curtis) (Trichoptera) exposed to elevated metal concentrations in a mine drainage stream. Hydrobiologia, 196: 91-100.
CrossRefDirect Link - Heliovaara, K., R. Vaisanen, H. Braunschweiler and M. Lodenius, 1987. Heavy metal levels in two biennial pine insects with sap-sucking and gall-forming life-styles. Environ. Pollut., 48: 13-23.
CrossRefDirect Link - Zvereva, E., V. Serebrov, V. Glupov and I. Dubovski, 2003. Activity and heavy metal resistance of non-specific esterases in leaf beetle Chrysomela lapponica from polluted and unpolluted habitats. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 135: 383-391.
CrossRefDirect Link - Nor, Y.M., 1987. Ecotoxicity of copper to aquatic biota: A review. Environ. Res., 43: 274-282.
CrossRefDirect Link - Bednarska, A.J. and R. Laskowski, 2009. Environmental conditions enhance toxicant effects in larvae of the ground beetle Pterostichus oblongopunctatus (Coleoptera: Carabidae). Environ. Pollut., 157: 1597-1602.
CrossRefDirect Link - Segner, H., K. Caroll, M. Fenske, C.R. Janssen and G. Maack et al., 2003. Identification of endocrine-disrupting effects in aquatic vertebrates and invertebrates: Report from the European IDEA project. Ecotoxicol. Environ. Saf., 54: 302-314.
CrossRefDirect Link - Moriber, L.G., 1956. A cytochemical study of hemipteran spermatogenesis. J. Morphol., 99: 271-327.
CrossRefDirect Link - Payne, F., 1966. Some observations on spermatogenesis in Gelastocoris oculatus (hemiptera) with the aid of the electron microscope. J. Morphol., 119: 357-381.
CrossRefDirect Link - Berry, A.J., J. Purvis and K.V. Radhakrishnan, 1992. Reproductive system and spermatogenesis in the opisthobranch gastropod Retusa obtusa (Montagu). J. Moll. Stud., 58: 357-367.
CrossRefDirect Link - Pijnacker, L.P. and M.A. Ferwerda, 1986. Development of the synaptonemal complex of two types of pachytene in oocytes and spermatocytes of Carausius morosus Br. (Phasmatodea). Chromosoma, 93: 281-290.
CrossRefDirect Link - Fernandes, A.P., G. Curi, FG.R. Francca and S.N. Bao, 2001. Nuclear changes and acrosome formation during spermiogenesis in Euchistus heros (Hemiptera: Pentatomidae). Tissue Cell, 33: 286-293.
CrossRefDirect Link - Fernandes, A.P. and S.N. Bao, 1998. Spermiogenesis in phytophagous bug (Hemiptera, Pentatomidae): An ultrastructural study. J. Submicrosc. Cytol. Pathol., 30: 485-493.
PubMedDirect Link - Kubo-Irie, M., M. Irie, T. Nakazawa and H. Mohri, 2003. Ultrastructure and function of long and short sperm in cicadidae (Hemiptera). J. Insect Physiol., 49: 983-991.
CrossRefDirect Link - Yue, L., T.L. Karr, D.F. Nathan, H. Swift, S. Srinivasan and S. Lindquist, 1999. Genetic analysis of viable Hsp90 alleles reveals a critical role in Drosophila spermatogenesis. Genetics, 151: 1065-1079.
Direct Link - Braeckman, B. and H. Raes, 1999. The ultrastructural effect and subcellular localization of mercuric chloride and methylmercuric chloride in insect cells (Aedes albopictus C6/36). Tissue Cell, 31: 223-232.
CrossRefDirect Link - Freeman, B.A. and J.D. Crapo, 1982. Biology of disease: Free radicals and tissue injury. Lab. Invest., 47: 412-426.
PubMedDirect Link - Au, D.W.T., O.V. Yurchenko and A.A. Reunov, 2003. Sublethal effects of phenol on spermatogenesis in sea urchins (Anthocidaris crassispina). Environ. Res., 93: 92-98.
CrossRefDirect Link - Acharya, U.R., R.M. Rathmore and M. Mishra, 2003. Role of vitamin C on lead acetate induced spermatogenesis in Swiss mice. Environ. Toxicol. Pharmacol., 13: 9-14.
CrossRefDirect Link - Eason, C.T. and P. Turck, 2002. A 90-day toxicological evaluation of compound 1080 (sodium monofluoroacetate) in Sprague-Dawley rats. Toxicol. Sci., 69: 439-447.
CrossRefDirect Link - Vandenbulcke, F., C. Grelle, M.C. Fabre and M. Descamps, 1998. Ultrastructural and autometallographic studies of the nephrocytes of Lithobius forficatus L. (Myriapoda, Chilopoda): Role in detoxification of cadmium and lead. Int. J. Insect Morphol. Embryol., 27: 111-120.
CrossRefDirect Link - Palumbo, G., M. Berloco, L. Fanti, M. Bozzetti and S. Massari et al., 1994. Interaction systems between heterochromatin and euchromatin in Drosophila melanogaster. Genetics, 94: 267-274.
CrossRefDirect Link - Warchalowska-Sliwa, E., M. Niklinska, A. Gorlich, P. Michailova and E. Pyza, 2005. Heavy metal accumulation, heat shock protein expression and cytogenetic changes in Tetrix tenuicornis (L.) (Tetrigidae, Orthoptera) from polluted areas. Environ. Pollut., 133: 373-381.
CrossRefDirect Link - Au, D.W.T., A.A. Reunov and R.S.S. Wu, 2001. Reproductive impairment of sea urchin upon chronic exposure to cadmium. Part II: Effects on sperm development. Environ. Pollut., 111: 11-20.
CrossRefDirect Link - Hancock, A.D. and D.M. de Kretser, 1992. The axonemal ultrastructure of spermatozoa from men with asthenospermia. Fertil. Steril., 57: 661-664.
PubMedDirect Link - Buckland-Nicks, J. and F.S. Chia, 1986. Fine structure of Sertoli cells in three marine snails with a discussion on the functional morphology of Sertoli cells in general. Cell Tissue Res., 245: 305-313.
CrossRefDirect Link - Van Vuren, J.H.J. and J.T. Soley, 1990. Some ultrastructural observations of leydig and sertoli cells in the testis of Tilapia rendalli following induced testicular recrudescence. J. Morphol., 206: 57-63.
CrossRefDirect Link