
Mojtaba Esmaeily
Department of Plant Protection, College of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
Ali Reza Bandani
Department of Plant Protection, College of Agriculture and Natural Resources, University of Tehran, Karaj, Iran
Journal of Entomology
Year: 2016 | Volume: 13 | Issue: 5 | Page No.: 179-186







ABSTRACT
Background and Objective: The elm leaf beetle, Xanthogaleruca luteola Muller (Coleoptera: Chrysomelidae) is one of the most important municipal insect pests of elm trees (Ulmus spp.) in Iran. Aim of this study was investigating effects of proteinaceous extracts of cereal and legume seeds to the X. luteola α-amylase in in vitro condition. Materials and Methods: Gut enzymes were extracted using sodium phosphate buffer and seed proteinaceous compounds were extracted using 0.1 M NaCl. The α-amylase activity assay was performed using the dinitrosalicylic acid (DNS) procedure. Results: The results showed that mung bean, pea, broad bean, barley, triticale, alborz and kavir inhibited the enzyme activity of insect. Kavir had the best inhibition activity with 90 and 84% of inhibition, respectively. Conclusion: Inhibitors of mung bean and kavir seeds have a good potential to be use in plant protection. So, it could be included that these inhibitors are more promising candidates for this pest control and decrees the chemical insecticide with adverse effect in human health.
PDF Abstract XML References Citation
Received: February 17, 2016;
Accepted: August 13, 2016;
Published: October 15, 2016
How to cite this article
Mojtaba Esmaeily and Ali Reza Bandani, 2016. Inhibition of Larvae and Adult α-amylase in Xanthogaleruca luteola Muller (Col.: Chrysomelidae) by Cereal and Legumes Seed Proteinaceous Extracts. Journal of Entomology, 13: 179-186.
DOI: 10.3923/je.2016.179.186
URL: https://scialert.net/abstract/?doi=je.2016.179.186
DOI: 10.3923/je.2016.179.186
URL: https://scialert.net/abstract/?doi=je.2016.179.186
INTRODUCTION
The elm leaf beetle, Xanthogaleruca luteola Muller (Coleoptera: Chrysomelidae) is one of the most important municipal insect pests of elm trees (Ulmus spp.) in Iran. Both adults and larvae feed on the parenchyma of leaves, without consuming the veins and at times the damage may affect all foliage and trees become brown1.
To date, use of broad spectrum pesticides in urban area cause many problems for human and environment health. So, a more benign control method should be used in the infested area in order not to contaminate environment which could be based on physiological process. To achieve this purpose, researchers should choose methods to develop different pest control approaches using molecules that are more environmentally friendly2,3. One alternative methodology that has been suggested is the use of plant genetic engineering to increase plant resistance to elm leaf beetle.
Digestion of carbohydrates begins in the elm leaf beetle by amylase (α-1, 4 glucan 4 glucanohydrolases; EC 3.2.1.1) that catalyze hydrolysis of α-D-(1, 4)-glucan linkage in starch components to produce energy. As coleopteran pest feed exclusively on carbohydrates during larval and/or adult stages, interfering with enzyme may cause retarding in normal growth and development4.
There are six diverse classes of α-amylase inhibitor which were able to interfere amylase activity; lectin-like, knottin-like, cereal-type, Kunitz-like, c-purothionin-like and thaumatin-like5. Cereals and legumes seeds are the rich sources of digestive enzyme inhibitors5-8 and also it was reported well-characterized amylase inhibitors are originate in plants for example the bean9, wheat10, barely11 and amaranth12 and these have been shown to be active next to some insect and mammalian amylases.
So, it is worthwhile to characterize digestive enzymes as well as to do in vitro and in vivo bioassay with plant proteinaceous inhibitors in order to reach a control strategy based on digestive enzyme inhibitors13.
There are many investigations about effect of cereal and legumes seed proteinaceous extracts on digestive enzymes in many pests. For example, Esmaeily and Bandani14 studied effect of the proteinaceous extract of wheat cultivars and some other plants on Tuta absoluta (Lep.: Gelechiidae) α-amylase. They showed that wheat cultivar seed extracts produced the greatest inhibition, so that Aflak cultivar extract inhibited 81% of the insect amylase. Bannakan et al.15 studied the effect of crude extracts of mung bean seeds on Callosobruchus maculatus (Col.: Bruchidae). Their results showed that with increasing the concentration of inhibitor the enzyme activity were decreased and the best pH 6 was the best pH for amylase activity. Also, Rahimi and Bandani16 studied the effect of seed proteinaceous extracts of rice (Oryza sativa L.), bean (P. vulgaris L.) and cowpea (Vigna unguiculata L.) on Eurygaster integriceps puton (Hem.: Scutelleridae) α-amylase activity and found that rice extract did not affect considerably α-amylase, on the other hand, they showed that bean and cowpea affected the insect α-amylase significantly.
Biochemical screening will continue to play an important role in the search for inhibitors with desirable characteristics and also a thorough understanding of inhibitory effect of these inhibitors on different pest enzymes.
However there is no study about effect of different seed proteinaceous extracts against the Xanthogaleruca luteola α-amylase. Thus, the aim of the present study was to investigate the effect of proteinaceous extracts of cereal and legume seeds to the X. luteola α-amylase. These plant seeds are mung bean (Vigna radiate L.), pea (Pisum sativum L.), broad bean (Vicia faba L.), barley (Hordeum vulgare L.), Triticale (×Triticosecale wittmack) and two wheat (Triticum aestivum L.) cultivars (Alborz and kavir).
MATERIALS AND METHODS
Insects: The elm leaf beetles were collected from infested elm trees in the Faculty of Agriculture, the University of Tehran at Karaj. The insects were reared in laboratory conditions at 25±1°C with 16:8 h photoperiod and 65±5% humidity as described by Tatli et al.17.
Midgut pH determination: To find out gut pH, standard indicator dyes were used. These indicators consist of bromophenol blue (pH 3.0-4.6), methyl red (pH 4.4-6.2), bromcresol purple (pH 5.2-6.8), bromophenol blue (pH 6.2-7.6), natural red (pH 6.8-8.0), cresol red (pH 7.2-8.8), thymol blue (8.0-9.6) and alizarin yellow (pH 10-12). The gut pH was determined based on Bignell and Anderson18 procedure with slight modifications. Last instars larvae and adults were dissected and their midguts were separated and kept in watch glass, then 10 μL of each pH indicator was added to the sample and the color was recorded.
Enzyme extraction: Enzyme samples were prepared by the method of Cohen19 and Bandani et al.20 with some modifications. Adults and larvae were at random selected and individual guts were dissected under a light microscope in ice cold 0.02 M sodium phosphate buffer (pH 6). The midgut was separated from the alimentary canal, rinsed in the buffer, placed in a pre-cooled homogenizer and ground into the phosphate buffer. The homogenates from midguts were transferred to centrifuge tubes and centrifuged at 15000×g for 15 min at 4°C. The supernatant was pooled and stored at -20°C for later analysis.
Protein extraction: Proteinaceous seed extracts of cereal and legumes including bean, mung bean, pea, broad bean, barley, triticale and two wheat cultivars (Alborz, kavir) were extracted according to the methods of Baker21 and Melo et al.22. Each seed (30 g) was grounded and mixed with a solution of 0.1 M NaCl and stirred for 1.5 h at 4°C, followed by centrifugation at 8,000 g for 30 min at 4°C. The supernatant was taken and heated at 70°C for 30 min to inactivate endogenous enzymes. Fractionation of the supernatant was done using 20, 40, 60, 80% ammonium sulfate saturations. Seventy percent ammonium sulfate saturated solution containing the maximum fraction of α-amylase inhibitors was dissolved in phosphate buffer (0.02 M and pH 7) and dialyzed against the same buffer overnight. This dialyzed solution was used as a supply of proteinaceous extract.
α-amylase activity assay: The α-amylase activity assay was performed using the dinitrosalicylic acid (DNS) procedure Bernfeld23 and Esmaeily and Bandani14, using 1% soluble starch (Merck, Darmstadt, Germany) as the substrate. Ten microliters of the enzyme sample were incubated for 30 min at 35°C with 500 μL MES buffer (pH 6) and 40 μL soluble starch. The reaction was stopped with the adding of 100 μL DNS and heating in hot water for 10 min. The DNS is a color reagent which reacts with the reducing groups released from starch by α-amylase action. The boiling water stops the α-amylase activity and catalyzes the reaction between DNS and the reducing groups of starch. Absorbance was then measured at 540 nm.
α-amylase inhibition assay: Effect of the seed extracts on larvae and adult α-amylase activity was determined as described by Mehrabadi et al.24. Enzyme extracts was pre-incubated with proteinaceous extracts (Mung bean, pea, broad bean, barley, triticale and two wheat cultivars including alborz and kavir for 30 min at 35°C followed by determination of the enzyme activity as described before using dinitrosalicilic acid (DNS) method. Appropriate blanks were included in the experiments as well. The inhibition percentage (I%) was calculated as follows:
where, A540 control is the absorbance for the control and A540 Exp is the absorbance for the experiment.
Effect of pH on α-amylase inhibition: To determine the effect of pH on inhibitory activity of the seed extracts, highest concentration of (10 μg protein) of kavir and mung bean extracts were incubated along with the enzyme for 30 min at pH set at 3-9 and then enzyme activity was recorded.
In gel α-amylase inhibition assay: The effect of two seed extracts including mung bean and kavir that produced the maximum amount of α-amylase inhibition were tested in the gel assays. The concentrations of protein extracts for both mung bean and kavir were used in gel assays were 10, 5, 2.5, 1.25 and 0.625 μg protein. Electrophoretic detection of amylolytic activity in the gel was done based on the procedure described by Laemmli25. In brief, PAGE was performed in 10% (w/v) gel for separating gel and 5% for stacking gel with 0.05% SDS. Electrophoresis was conducted at a voltage of 90 V until the blue dye reached the underside of the gel. The gel was rinsed with distilled water and washed by 1% (v/v) triton X100 for 20 min after that the gel was incubated in MES buffer (pH 6) containing 1% starch solution, 2 mM CaCl2 and 10 mM NaCl for 1.5 h. Finally, the gel was treated with a solution of 1.3% I2 and 3% KI to stop the reaction and to stain the un-reacted starch background. Regions of α-amylase activities appeared at the light band against the dark background.
Protein determination: Protein concentrations were measured using the method of Bradford26 and using bovine serum albumin (Bio-Rad, München, Germany) as a standard.
Statistical analysis: One-way analysis of variance (ANOVA) was used to compare the data followed by an Least Significant Difference (LSD, 5%) test, when significant differences were found SAS27.
RESULTS
Determination of the midgut pH: It was found that the both larvae and adults of X. luteola midgut pH were acidic. When larvae and adults were dissected and their contents were mixed with dye indicators, it was found that pH were 6.0±0.5 and 6.5±0.5, respectively.
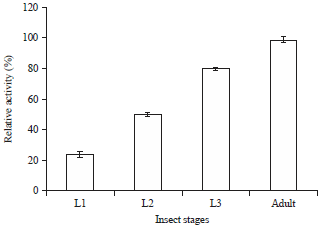 | |
| Fig. 1: | Digestive α-amylase activities during development of X. luteola |
 | |
| Fig. 2: | Effect of seed proteinaceous extracts on adult α-amylase of X. luteola |
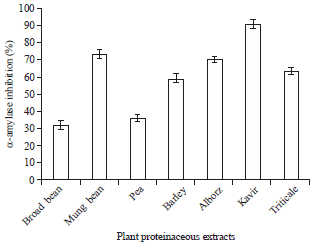 | |
| Fig. 3: | Effect of seed proteinaceous extracts on larvae α-amylase of X. luteola |
α-amylase activity in different developmental stages: Figure 1 shows α-amylase activity in different developmental stages of X. luteola. The activity of enzyme shows a firm state increase from the first instar to adult, so that last instar larvae and adults have most activity of α-amylase.
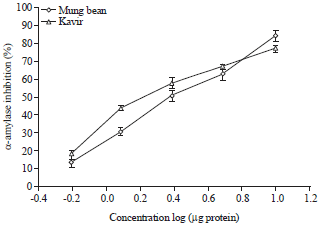 | |
Fig. 4: | Dose-response curve of inhibition of larvae α-amylase by kavir and mung bean proteinaceous extracts |
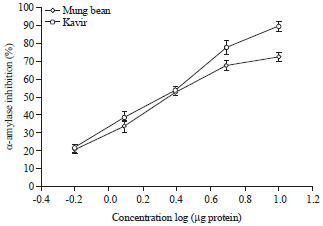 | |
| Fig. 5: | Dose-response curve of inhibition of adult α-amylase by kavir and mung bean proteinaceous extracts |
Therefore, these were chosen for more studies and were examined effect of proteinaceous extracts on their α-amylase activity.
Effect of seed proteinaceous extracts on α-amylase activity: When seed extracts were assayed against larvae and adults α-amylase, the results showed that mung bean, pea, broad bean, barley, triticale, alborz and kavir inhibited the enzyme activity with the percentage of 73, 36, 32, 59, 63, 70, 90 and 77, 21, 26, 69, 63, 73, 84, respectively (Fig. 2, 3).
Mung bean and kavir extracts produced the highest amount of α-amylase inhibition. Therefore, based on these results, these extracts were chosen for more studies. When five different concentrations of seed extracts were assayed against last instar larvae and adult’s α-amylase, a concentration dependent trend was observed (Fig. 4, 5).
Effect of pH on inhibitory activity of the seed extracts: Effect of pH on the inhibition of the larvae and adult α-amylase showed that the most inhibition of mung bean and kavir seed extracts were observed at pH 6.0 that is the optimum pH for the activity of this enzyme in the in vitro condition (Fig. 6, 7).
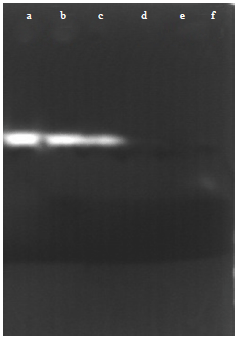
In gel inhibition assay of α-amylase: Gel assays showed that larvae and adult α-amylase of the insect gut were affected by the presence of the seed extracts as it has been shown in the spectrophotometric assays. Also, it was found that in the different concentrations of the extracts, the intensity of the bands varied due to differences in enzyme inhibition. So that, in both larvae and adults, when a high dose was used, no α-amylase band was observed and as low doses was employed, the intensity of α-amylase band was decreased in comparison with control (Fig. 8-11).
 | |
| Fig. 6: | Effect of pH values on the inhibitory activity of kavir and mung bean α-amylase inhibitors towards adult α-amylase |
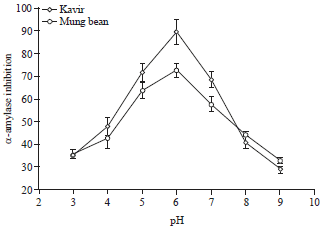 | |
| Fig. 7: | Effect of pH values on the inhibitory activity of kavir and mung bean α-amylase inhibitors towards larvae α-amylase |
 | |
| Fig. 8: | In gel assay of the effect of kavir extract on the adult α-amylase activity, a: First column of the left hand side shows control. With increasing the inhibitor concentrations the amount of the enzyme activity decreases, f: 10 μg protein extract, e: 5 μg protein extract, d: 2.5 μg protein extract, d: 1.25 μg protein extract and b: 0.625 μg protein extract |
 | |
| Fig. 9: | In gel assay of the effect of mung bean extract on the adult α-amylase activity, a: First column of the right hand side shows control. With increasing the inhibitor concentrations the amount of the enzyme activity decreases, f: 10 μg protein extract, e: 5 μg protein extract, d: 2.5 μg protein extract, c: 1.25 μg protein extract and b: 0.625 μg protein extract |
 | |
| Fig. 10: | In gel assay of the effect of kavir extract on the larvae α-amylase activity, a: First column of the left hand side shows control. With increasing the inhibitor concentrations the amount of the enzyme activity decreases, f: 10 μg protein extract, e: 5 μg protein extract, d: 2.5 μg protein extract, c: 1.25 μg protein extract and b: 0.625 μg protein extract |
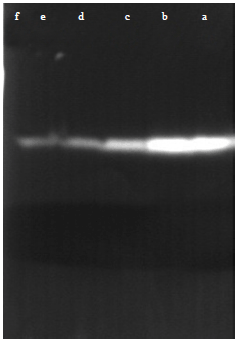 | |
| Fig. 11: | In gel assay of the effect of mung bean extract on the larvae α-amylase activity, a: First column of the right hand side shows control. With increasing the inhibitor concentrations the amount of the enzyme activity decreases, f: 10 μg protein extract, e: 5 μg protein extract, d: 2.5 μg protein extract, c: 1.25 μg protein extract and b: 0.625 μg protein extract |
DISCUSSION
It has been generally suggested that physio-chemical midgut conditions, especially pH of midgut contents are a most important factor that affects digestive enzymes28. It is speculated that the midgut pH in larvae and adult of X. luteola were acidic that are corresponding with α-amylase activity of this insect, so, it has been reported optimum pH of α-amylase activity of this pest is 6.0 Tatli et al.17.
In the present study, it was found that seed proteinaceous extracts of seven plant species including mung bean, pea, broad bean, barley, triticale and two wheat cultivars (Alborz and kavir) affected larvae and adult α-amylase activity of the elm leaf beetle. However, the effect of seed extracts on the α-amylase activity was different. So that, kavir and mung bean proteinaceous extracts created most inhibition of α-amylase in both larvae and adults. Different inhibitions indicated that different plant species produce different metabolite with different specificity in order to protect themselves.
As mentioned above it has been many studies about the specificity of inhibitory effect of plant proteinaceous extracts on the α-amylase activity of insects14,16.
The tests completed with greatest inhibition of α-amylase activity of larvae and adult made by kavir and mung bean proteinaceous extracts at pH 6 which is the optimum pH for the activity of this enzyme in vitro condition. Similar investigation to the study was conducted by Valencia-Jimenez et al.29 showed that Phaseolus coccineus L. and Phaseolus vulgaris L., seed proteinaceous extracts inhibited the α-amylase activity of Tecia solanivora Povolny (Lepidoptera: Gelechiidae) at pH 6.0 by 70 and 87%, respectively. However, the α-amylase inhibitor from amaranth seeds inhibited 80% of the insect amylase activity at pH 9.0 that was optimum pH of this pest in vitro condition. So, since amaranth seed inhibitor inhibited α-amylase activity at optimum pH, they concluded this inhibitor is more promising candidates for potato transformation than inhibitors from two beans.
The interaction between α-amylase and inhibitors occurs in the insect gut. As mentioned X. luteola larvae and adult have an acidic midgut pH range from 5.5-6.5 and 6.0-7, respectively and it has been reported that it has a slightly acidic optimum pH 6 for protease activity17. Therefore, it can be expected that α-amylase activity would be high under such conditions and for it to be inhibited if mung bean and kavir proteinaceous extracts are present at a high enough concentration. The accordance between gut lumen pH, amylase optimal pH and pH optimum for amylase inhibition by plant amylase inhibitors has been described in other insect studies30,31.
CONCLUSION
In conclusion, based on the results, it can be concluded that extracts from different species had different effects on the insect alpha amylase because of their unique metabolite/s. Also, it should be indicated that mung bean and kavir seed inhibitors have a good potential to inhibition of the elm leaf beetle amylase activity. So, it could be included that inhibitors from mung bean and kavir seeds are more promising candidates for elm transformation than other inhibitors.
SIGNIFICANCE STATEMENT
This study was the first research on effectiveness of plant inhibitor on elm leaf beetle that data can be used on control of the pest and decrease the application of chemical insecticide and helps to public health.
ACKNOWLEDGMENTS
The authors would like to acknowledge the financial support of the University of Tehran for this research under grant number 7110023/6/15.
REFERENCES
- Huerta, A., I. Chiffelle, K. Puga, F. Azua and J.E. Araya, 2010. Toxicity and repellence of aqueous and ethanolic extracts from Schinus molle on elm leaf beetle Xanthogaleruca luteola. Crop Protect., 29: 1118-1123.
CrossRefDirect Link - Isman, M.B., 2006. Botanical insecticides, deterrents and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol., 51: 45-66.
CrossRefPubMedDirect Link - Franco, O.L., D.J. Rigden, F.R. Melo, C. Bloch Jr., C.P. Silva and M.F. Grossi de Sa, 2000. Activity of wheat α-amylase inhibitors towards bruchid α-amylases and structural explanation of observed specificities. Eur. J. Biochem., 267: 2166-2173.
CrossRefPubMedDirect Link - Franco, O.L., D.J. Rigden, F.R. Melo and M.F. Grossi-de-Sa, 2002. Plant α-amylase inhibitors and their interaction with insect α-amylases: Structure, function and potential for crop protection. Eur. J. Biochem., 269: 397-412.
CrossRefDirect Link - Svensson, B., K. Fukuda, P.K. Nielsen and B.C. Bonsager, 2004. Proteinaceous α-amylase inhibitors. Biochim. Biophys. Acta, 1696: 145-156.
CrossRefDirect Link - Payan, F., 2004. Structural basis for the inhibition of mammalian and insect α-amylases by plant protein inhibitors. Biochimica et Biophysica Acta (BBA)-Proteins Prot., 1696: 171-180.
CrossRefDirect Link - Sivakumar, S., M. Mohan, O.L. Franco and B. Thayumanavan, 2006. Inhibition of insect pest α-amylases by little and finger millet inhibitors. Pesticide Biochem. Physiol., 85: 155-160.
CrossRefDirect Link - Powers, J.R. and J.R. Whitaker, 1978. Purification and some physical and chemical properties of red kidney bean (Phaseolus vulgaris) α-amylase Inhibitor. J. Food Biochem., 1: 217-238.
CrossRefDirect Link - Abe, J., U. Sidenius and B. Svensson, 1993. Arginine is essential for the α-amylase inhibitory activity of the α-amylase/subtilisin inhibitor (BASI) from barley seeds. Biochem. J., 293: 151-155.
CrossRefPubMedDirect Link - Chagolla-Lopez, A., A. Blanco-Labra, A. Patthy, R. Sanchez and S. Pongor, 1994. A novel α-amylase inhibitor from amaranth (Amaranthus hypocondriacus) seeds. J. Biol. Chem., 269: 23675-23680.
PubMedDirect Link - Harrison, R.L. and B.C. Bonning, 2010. Proteases as insecticidal agents. Toxins, 2: 935-953.
CrossRefDirect Link - Esmaeily, M. and A.R. Bandani, 2015. Interaction between larval α-amylase of the tomato leaf miner, Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) and proteinaceous extracts from plant seeds. J. Plant Protect. Res., 55: 278-286.
CrossRefDirect Link - Bannakan, I., P. Hormchan, A. Wongpiyasatid and A. Engkakul, 2007. Effects of α-amylase inhibitor on mungbean weevil, Callosobruchus maculatus, in vivo and in vitro and on barley malt α-amylase in vitro. Kasetsart J. (Nat. Sci.), 41: 451-460.
Direct Link - Rahimi, V. and A.R. Bandani, 2014. Comparison of the effects of cereal and legume proteinaceous seed extracts on α-amylase activity and development of the Sunn pest. J. Asia-Pacific Entomol., 17: 7-11.
CrossRefDirect Link - Tatli, I., A.R. Bandani and A. Moslemi, 2013. The elm leaf beetle α-amylase and its activity relationship with insect feeding. Arch. Phytopathol. Plant Protect., 46: 917-926.
CrossRefDirect Link - Bignell, D.E. and J.M. Anderson, 1980. Determination of pH and oxygen status in the guts of lower and higher termites. J. Insect Physiol., 26: 183-188.
CrossRefDirect Link - Cohen, A.C., 1993. Organization of digestion and preliminary characterization of salivary trypsin-like enzymes in a predaceous heteropteran, Zelus renardii. J. Insect Physiol., 39: 823-829.
CrossRefDirect Link - Bandani, A.R., M. Kazzazi and M. Mehrabadi, 2009. Purification and characterization of midgut α‐amylases of Eurygaster integriceps. Entomol. Sci., 12: 25-32.
CrossRefDirect Link - Baker, J.E., 1987. Purification of isoamylases from the rice weevil, Sitophilus oryzae (L.) (Coleoptera: Curculionidae), by high-performance liquid chromatography and their interaction with partially-purified amylase inhibitors from wheat. Insect Biochem., 17: 37-44.
CrossRefDirect Link - Mehrabadi, M., A.R. Bandani and F. Saadati, 2010. Inhibition of Sunn pest, Eurygaster integriceps, α-amylases by α-amylase inhibitors (T-αAI) from Triticale. J. Insect Sci., Vol. 10, No. 1.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Terra, W.R. and C. Ferreira, 1994. Insect digestive enzymes: Properties, compartmentalization and function. Comp. Biochem. Physiol., 109: 1-62.
CrossRefDirect Link - Valencia-Jimenez, A., V.J.W. Arboleda, A.L. Avila and M.F. Grossi-de-Sa, 2008. Digestive α-amylases from Tecia solanivora larvae (Lepidoptera: Gelechiidae): Response to pH, temperature and plant amylase inhibitors. Bull. Entomol. Res., 98: 575-579.
CrossRefDirect Link - Biggs, D.R. and P.G. McGregor, 1996. Gut pH and amylase and protease activity in larvae of the New Zealand grass grub (Costelytra zealandica; Coleoptera: Scarabaeidae) as a basis for selecting inhibitors. Insect Biochem. Mol. Biol., 26: 69-75.
CrossRefDirect Link - Valencia, A., A.E. Bustillo, G.E. Ossa and M.J. Chrispeels, 2000. Alpha-amylases of the coffee berry borer (Hypothenemus hampei) and their inhibition by two plant amylase inhibitors. Insect Biochem. Mol. Biol., 30: 207-213.
CrossRefPubMedDirect Link