
Jing-Ee Yii
Department of Crop Science, Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia Bintulu Sarawak Campus, Nyabau Road, 97000, Bintulu, Sarawak, Malaysia
Choon-Fah J. Bong
Department of Crop Science, Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia Bintulu Sarawak Campus, Nyabau Road, 97000, Bintulu, Sarawak, Malaysia
Jie-Hung P. King
Department of Crop Science, Faculty of Agriculture and Food Sciences, Universiti Putra Malaysia Bintulu Sarawak Campus, Nyabau Road, 97000, Bintulu, Sarawak, Malaysia
Kadir Jugah
Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
Journal of Entomology
Year: 2016 | Volume: 13 | Issue: 1-2 | Page No.: 1-10
DOI: 10.3923/je.2016.1.10







ABSTRACT
Present study was conducted to investigate the feeding preferences of the pestiferous termite species in oil palm plantation, Coptotermes curvignathus and to develop an effective combination of toxicants and phagostimulant with concepts of a cost-effective and environmental friendly termite baiting technology. In choice feeding bioassays, size of bait and nutrient were investigated and found to significantly influence termite feeding preference. The results showed rubber wood sawdust was consumed 4-6 times more than rubber wood in granular and block form by C. curvignathus. It signified that sawdust was palatable and in easier consumable form than solid wood block and granules, which needed to be chewed by termite workers. Whereas for the nutrient preference test, glucose was most preferred by termites with the greater consumption of paper discs compared to other carbon and nitrogen sources. Formulated bait with combinations of 0.001 mg a.i L–1 fipronil, 108 conidia g–1 bait Metarhizium anisopliae and 1% glucose gave the greatest synergistic effect (χ2 = 17.889), followed by 108 conidia g–1 bait with glucose. The results demonstrated that glucose might be a key additive in development of an effective bait system for subterranean termite management.
PDF Abstract XML References Citation
Received: August 12, 2015;
Accepted: October 22, 2015;
Published: February 15, 2016
How to cite this article
Jing-Ee Yii, Choon-Fah J. Bong, Jie-Hung P. King and Kadir Jugah, 2016. Feeding Preferences of Oil Palm Pest Subterranean Termite Coptotermes curvignathus (Isoptera: Rhinotermitidae). Journal of Entomology, 13: 1-10.
DOI: 10.3923/je.2016.1.10
URL: https://scialert.net/abstract/?doi=je.2016.1.10
DOI: 10.3923/je.2016.1.10
URL: https://scialert.net/abstract/?doi=je.2016.1.10
INTRODUCTION
Termite control in oil palm plantations in peatland is a never-ending story in the industry. It is estimated that around 20% of the oil palms (≈2.4 million ha) are cultivated on peat in South East Asia region (Lim et al., 2001). To date, Malaysia has 666,000 ha of oil palm planted on peat soil and the total losses due to termite infestation reached hundreds of million dollars each year with estimation of $87.2 ha–1 year–1 at a density of 160 palms ha–1 (Zulkefli et al., 2012). The oil palm plantations are forced to incur enormous cost yearly on prophylactic termite treatments, mainly using chemical spraying. However, the termite colonies are never eliminated, whereby re-infestation of termites occurs frequently after 3-6 months.
Hence, eco-friendly control methods like termite bait was introduced for managing subterranean termites since 20 years ago after commercial release in developed country as a result of growing environmental awareness (Evans and Iqbal, 2015). The main purpose of implement baiting is to kill the specific target by using a smidgen of active ingredient into food matrix to eliminate the colony without affecting non-target organisms. Termite baiting faces the greatest challenge in that the owners of those oil palm plantations are still doubtful on termite baiting because the effect of the control is slow compared to synthetic chemical pesticides. Besides, the higher price of bait in current market hinders termite baiting to forge ahead in the oil palm industry. Researchers are now exploring a more attractive and palatable bait by adding phagostimulants, additives and attractants.
A palm can be killed when the termites consume up the living meristematic tissue in less than 2 months and further spread to neighbouring palms (Lim et al., 2012). Hence, an effective bait ought to be discovered quickly by a termite colony, accepted and also be preferred over the numerous food sources in natural environment. Once they accept the bait as a food source, the chance is higher that the colony population will be eliminated when adequate bait toxin is taken up. Food selection of termites is influenced by many factors (Traniello and Leuthold, 2000). Apparently, termite colony caste dynamics require nutrients for growth and reproduction. This dietary requirement for termites may change seasonally (Botch et al., 2010). According to Reinhard and Kaib (2001), phagostimulants may influence termite feeding preferences. Numerous studies also have investigated on wood rot fungi (Cornelius et al., 2002; Su, 2005; Ding et al., 2015), sugars (Reinhard and Kaib, 2001; Waller and Curtis, 2003; Saran and Rust, 2005; Haifig et al., 2010; Castillo et al., 2013), amino acids (Chen and Henderson, 1996), carbon dioxide (Bernklau et al., 2005; Broadbent et al., 2006), pheromone (Reinhard et al., 2002a, b) and sport drinks (Getty et al., 2007; Cornelius and Osbrink, 2008) in order to discover and improve on the efficacy, acceptance and enhancement of bait uptake by termites in the bait station.
Among the termite management tools, integrated fungus and chemical pesticides are seldom being discussed especially in termite baiting. Evans and Iqbal (2015) had summarized neurotoxin as the fastest active ingredient to cause colony elimination, followed by chitin synthesis inhibitors and metabolic inhibitors. Nevertheless, it was too fast acting for baiting, whereby higher doses of active ingredient may kill the termites in situ or induced avoidance behaviour among the termites. However, using chemical pesticide at low doses was found successful in some baiting studies (Henderson and Forschler, 1996; Huang et al., 2006). Among all, non repellent and slow-acting fipronil is well known as termiticide in plantations. Incorporating fungus Metarhizium anisopliae spores and chemical insecticide fipronil in rubber wood sawdust as bait was proven to give synergistic effect on termite mortalities in preliminary study. Meanwhile, incorporated massive amount of fungus or virulent fungal strains were more likely to be recognized by termite and triggered avoidance behaviour (Rath, 2000) in termite colonies. To minimize the defensive behaviour of termites, adding attractant with pathogens might be the way to disguise or cover the odour of the spores from termites. A successful case was implemented among subterranean termite Odontotermes obesus, whereby the mixing of conidia with attractants was able to enhance the dissemination of pathogens among the population and caused an epizootic (Balachander et al., 2013).
Different species of termites vary greatly in their feeding preferences (Ngee et al., 2004) because of their difference in ecology and feeding habit. The variations are even present within the same genus of termite. For instance, Coptotermes gestroi, R. flavipes and R. virginicus preferred sucrose with yeast solutions (Waller et al., 1999; Haifig et al., 2010), C. formosanus was attracted preferentially to glutamic acid and aspartic acid (Chen and Henderson, 1996) while arbutin was the favourite supplement for R. santonensis (Reinhard and Kaib, 2001). Thus, this study was conducted to investigate the feeding preferences of the pestiferous termite species in oil palm plantation, C. curvignathus in terms of nutrient and size of bait. Besides, the present study also attempted to develop an effective combination of toxicants and phagostimulant with concepts of a cost-effective and environmental-friendly termite baiting technology through synergistic bioassay of formulated bait.
MATERIALS AND METHODS
Experimental site: Studies were conducted in Laboratory of Entomology, Universiti Putra Malaysia Bintulu Sarawak Campus from year 2012-2013 to determine the feeding preferences of C. curvignathus and synergism of formulated bait against termites.
Collection of termites: Termites were collected from an oil palm estate, located approximately 70 km west of Bintulu, Sarawak, Malaysia. The plantation was developed on peat and three sites were set up as termites baiting stations located at Block 9 (N 03°00'47.1") with 9.93 ha, Block 13 (N 03°01'17.6") with 22.01 ha and Block 16 (N 03°01'00.6", E 112°52'51.7") with 39.41 ha. The rubber logs (length 25 cm×diameter 10 cm) were buried around the infested palm to attract the foraging termites to the bait stations. The collected termites were kept in termite culture room (28±1°C) inside dark plastic container with rubber logs as food source until used.
Feeding preference/Dietary pattern of termite: Feeding preference is crucial in formulating bait in order to increase the palatability of bait and phagostimulatory of termites. Different species of termites have their own feeding preference. The size of food and the nutrients preferred by C. curvignathus were investigated in multiple choice feeding tests.
Food size preference: Bait size preference of C. curvignathus was tested in laboratory. One hundred mature termites (90 workers+10 soldiers) were pre-incubated in 15 cm petri dish with moist sand in total darkness at 28±1°C. Rubber wood was used as food source and made into three different sizes of each bait, i.e., sawdust (0.05 cm), granular (0.5 cm) and block (0.5×0.5×2 cm3). The dry weight of bait was recorded pre and post-treatment to termites. The feeding trials were conducted for 7 days. The experiment was repeated twice with 5 replicates each. The amount of baits removed (presumably consumed) by termites was calculated to test the significant difference of size preferred by the termites.
Nutrient preference: Nutrient preferred by C. curvignathus was tested in the laboratory. Groups of termites each comprising 90 workers and 10 soldiers were pre-incubated for 1 day in 15 cm petri dish with moist sand in total darkness (28±1°C). Nine different nutrient solutions (1.0% v/v) were prepared and these included two monosaccharides (glucose, fructose), three disaccharides (lactose, maltose and sucrose), a polysaccharide (dextrin), three nitrogen sources (urea, yeast extract and peptone) and sterilized distilled water served as control. Filter paper (Whatman) discs with 2×2 cm2 dimension were cut and soak in the nutrient solutions for 15 min. The nutrient-treated filter paper discs were then dried in the oven (60°C) for 3 days. The dry weight of filter paper discs were recorded before and after treatments with termites for 7 days. Five repetitions were carried out with 5 replicates each. The consumption of filter papers by termites were analyzed to obtain the significant differences between treatments.
Based on the results of previous experiment, the highest consumption of carbon and nitrogen source nutrient was mixed and reevaluated against termites in choice feeding test. The preparation of nutrient-treated filter paper discs was same as above. The test was repeated thrice with 15 replicates each. After that, the most preferred nutrient was tested against termites using different concentrations (0.1, 1.0 and 3.0%). Filter paper disc soaked in sterilized distilled water served as control. Termite preferences were evaluated by calculating the consumption between treated and untreated filter paper discs.
Toxicity effect of formulated cellulose bait against C. curvignathus: Rubber wood sawdust (0.05 cm) was used as the substrate to develop three types of baits, i.e., fipronil bait (fb), Metarhizium bait (mb) and fipronil+Metarhizium bait (fmb) that incorporated both fipronil and Metarhizium with and without addition of glucose into the bait. This experiment was carried out to determine the interaction of glucose in bait against termites. Sawdust was immersed in several low concentrations of fipronil working solution (0.001, 0.01, 0.05, 0.1 mg a.i L–1 in acetone) and air dried for 2 day. Glucose solution (1.0% v/v) was added into the sawdust, mixed thoroughly and oven dried at 60°C for 3 days. Dry harvested pure M. anisopliae var. anisopliae isolate TA (Hoe et al., 2009) conidia from broken maize grain were mixed into fipronil treated rubber wood sawdust and adjusted to three concentrations of 107, 108 and 109 conidia g–1 of formulated bait. A total of 0.01 g bait was applied to twenty pre-conditioned termites (18 workers and 2 soldiers) per replicate. During the bioassay, cadavers of dead termites were transferred onto moist blotter to observe mycosis of fungus. Termite mortalities were recorded daily over a period of two weeks and mortality at 8 Days Post Treatment (DPT) were corrected using Abbott’s formula before subjecting to data analysis. The experiment was repeated thrice with five replications each.
Statistical analysis: All statistical analysis was performed using SAS 9.0. The size preference test was normalized using log transformation while the data of nutrient test and mortality percentage for each treatment were arcsine square root transformed prior to analysis using ANOVA. The means of consumption and termite mortality were separated using Duncan New Multiple Range Test (DNMRT) at p≤0.05. A factorial arrangement with completely randomized design was performed to test differences in the treatment combinations. The significant results indicated there was an interaction between bait and glucose and vice versa. Synergy between bait+glucose was analysed by comparing mortality rate induced by combinations of both agents (observed) with the sum of mortalities induced by each agent separately (expected). The expected mortality (ME) was obtained using formula: ME = Mb+Mg (1-Mb/100), where, Mb and Mg were the observed percent mortality caused by baits (fb, mb and fmb) without glucose and glucose alone, respectively. Chi square test (χ2) was performed by calculating the calculated χ2 value using formula: χ2 = (Mfmg-ME)2/ME, where, the Mfmg represents the observed mortality for the treatment combinations (fb, mb or fmb+glucose) and then compared to the table value for 1 df (>3.84). If the calculated χ2 value exceeds the tabulated value, it indicates non-additive effect (either synergistic or antagonistic) of the combinations. Significant interaction of the combination was determined through the difference between Mfmg-ME (positive = synergistic, negative = antagonistic). In contrast, if the tabulated value exceeds calculated χ2 value, it represents an additive effect at p≤0.05.
RESULTS
Size preference: The termite feeding preference was significantly affected by the sizes of bait (F = 79.39, df = 2, p<0.0001). The consumption of rubber wood sawdust (207.390 mg) was 6 times higher than those of the granular (33.000 mg) and 4 times higher than the block (52.570 mg) forms (Fig. 1).
Nutrient preference: In nutrient preference test, different nutrients significantly affected the consumption of filter paper disc by C. curvignathus (F = 7.74, df = 9, p<0.0001). Among all the nutrients, sugars such as glucose, lactose, fructose and sucrose comprised the carbon source were more palatable for C. curvignathus. However, among all the carbon sources, only glucose showed significant difference from the control and achieved the highest consumption (13.340 mg). There were not statistically significant differences in consumption of lactose, fructose, sucrose, dextrin and maltose when compared with control. In the choice for nitrogen source, termites consumed more of urea (10.368 mg) as compared to yeast extract (6.364 mg) and peptone (1.904 mg). Termites showed repellence to peptone with the least consumption of filter paper (Fig. 2). While in the test of glucose and urea combination, the result showed significant difference between treatments (F = 4.83, df = 3, p = 0.0032). A total of 12.271 mg filter paper was taken up by termites in glucose and urea combination and it was significantly different from the control. However, the combination treatment was not statistically significant from the single treatment of glucose (14.140 mg) and urea (13.769 mg) (Fig. 3). Considering the most cost-effective way to formulate the bait, glucose was chosen as the only nutrient for bait development and the termites most preferred glucose concentration was checked (F = 7.08, df = 3, p = 0.0003). The results showed C. curvignathus fed preferentially on filter paper treated with 1.0 and 3.0% glucose solution when compared to control but there was no significant difference between both concentrations of glucose. Meanwhile, 0.1% glucose solution was too little to induce the phagostimulatory of termites as the consumption was not different from that of control treatment (Fig. 4).
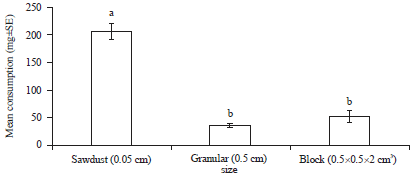 | |
| Fig. 1: | Mean consumption (mg±SE) of different sizes of rubber wood bait by Coptotermes curvignathus after 1 week in choice feeding test. Bars with different letters are significantly different at p≤0.05 (DNMRT) |
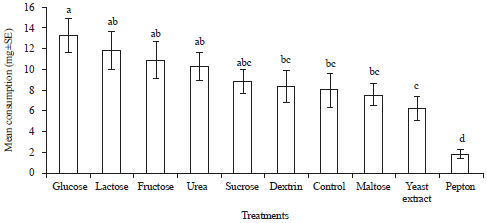 | |
| Fig. 2: | Mean consumption (mg±SE) of nutrient-treated filter paper by Coptotermes curvignathus after 1 week in multiple choices feeding test. Bars with different letters are significantly different at p≤0.05 (DNMRT) |
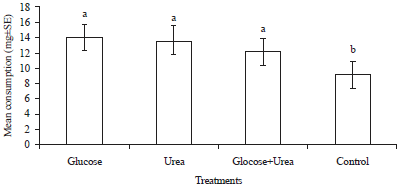 | |
| Fig. 3: | Mean consumption (mg±SE) of combination of nutrient-treated filter paper by Coptotermes curvignathus after 1 week in choice feeding test. Bar with different letters is significantly different at p≤0.05 (DNMRT) |
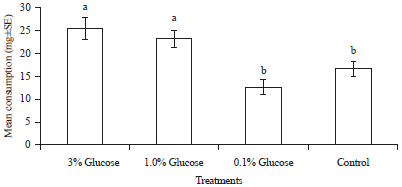 | |
| Fig. 4: | Mean consumption (mg±SE) of filter paper by Coptotermes curvignathus on different concentrations of glucose after 1 week in choice feeding test. Bar with different letters is significantly different at p≤0.05 (DNMRT) |
Toxicity effect of formulated bait: The results showed that termite mortality was significantly affected by different types of baits (F = 126.31, df = 19, p<0.0001) and interaction between bait and glucose (F = 2.85, df = 19, p<0.0001). Among all the treatments, the chi-square results showed synergistic interactions when incorporation with glucose in 108 and 0.001+108 combinations, wherein the greatest synergistic effect occurred when 0.001 mg a.i L–1 fipronil and 108 conidia g–1 bait acted synergistically with 1% glucose in formulated bait (χ2 = 17.889). Meanwhile, treatment 0.05+107 bait caused an antagonistic effect when glucose was incorporated into the bait. Most of the baits additively interacted with glucose in killing termites (Table 1).
| Table 1: | Synergy bioassay of different combination of fipronil and Metarhizium anisopliae in termite mortality at 8 DPT |
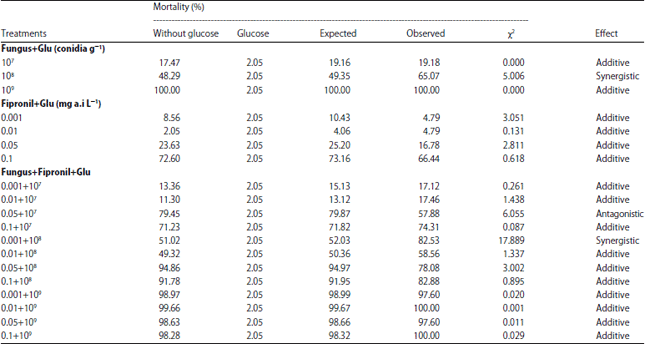 | |
| A chi-square comparison that exceeds 3.84 with df = 1 and α = 0.05 is considered synergistic or antagonistic interaction tab | |
DISCUSSION
To develop an economic agricultural product for termite baiting, it is fundamental to study the termite behaviour primarily on feeding preference of pestiferous termite species C. curvignathus in oil palm plantation. Food consumption is the best parameter to detect the phagostimulant of termites (Haifig et al., 2010). The results implied that C. curvignathus feeding preference varied according to food size. The rubber wood sawdust was consumed about 4-6 times more than wood block and granular form. The moisture content for three types of bait was examined and the initial moisture of rubber sawdust (2.31%) was significantly higher than granular (0.70%) and block (0.35%) form. However, there was no difference in moisture content of sawdust and granular after 1 day exposure to termites, whereby the moisture was increased to 29.88 and 29.34%, respectively. Notably, the moisture content of baits might not be the main factor affecting their choices. In the prospect of energy saving in termites, sawdust was easier to consume than solid wood, which needed to be chewed by termite workers. It reduces the energy used for mechanical break up of the rubber wood surface. This energy saving behaviour was also observed in termites when they attacked mature oil palms, whereby lesser energy was required when they attacked through the basal region of palm instead of climbing up >1 m in height up the palm trunk and then infested through the spear region, such as termite infestation on young palms (Chan et al., 2011). Hence, it is possible that termites tend to reduce energy requirement even in their daily feeding. Results of the current study was in accordance with Kasseney et al. (2011), whereby the termite Odontotermes formosanus also consumed more bait in flour type than solid wood.
Termite workers do not have the ability to choose the food visually or physically as they have no eyes. Thus, the possibility is that termites accomplished the wood size assessment by using vibroacoustic signals (Evans et al., 2005, 2007; Inta et al., 2007). As in Inta et al. (2007), different species of termites prefers a particular food size. Life history of termites is plausible as the main reason that make drywood termite Cryptotermes domesticus and C. secundus varied in their food size preference, whereby the former fed preferentially on smaller blocks of wood, while the latter preferred larger wood (Evans et al., 2005, 2007; Inta et al., 2007). However, there was paucity in food size study on other termites especially in genus Coptotermes. It is difficult to correlate their preference with their ecological behaviour. In nature, C. curvignathus prefers to attack variable sizes of crops and structural buildings, such as oil palm, coconut, mango, rubber trees, etc. (Wood, 1968; Harris, 1969) which are usually larger in size. While under experimental condition, they tend to feed upon the smaller size food rather than the larger, which is contradictory to their natural feeding habit.
Apart from the bait size, nutrients are important in the dietary of termites. Termites acquire these compounds from their food or via metabolism with the aid of symbionts (La Fage and Nutting, 1978). A multiple choices test of carbon and nitrogen food sources was designed to test the nutrient preference of C. curvignathus to ensure the selected nutrient was the most preferred and attractive over numerous food sources. Sugars are phagostimulatory for a wide range of insects, including termites. The sugars usually occur in wood and fungi, which are the primary food sources of subterranean termites. Current study proved that C. curvignathus prefer sugars such as glucose, lactose, fructose and sucrose. Sugar treatments appeared to have no effect on termite vitality (Waller and Curtis, 2003). Among all, glucose was the most favoured nutrient for termites in comparison to control. In nature, termites prefer cellulose which is derived from D-glucose units. The plants that are usually attacked by C. curvignathus such as oil palms and rubber trees both contain high amount of soluble sugars and starch (Henson et al., 1999; Chantuma et al., 2009). To compete with the abundant food sources in natural environment, inclusion of glucose in bait as attractant is necessary to attract more termites towards formulated bait.
The process of feeding and communal food exploitation in termites is strongly connected with their labial glands (Reinhard and Kaib, 2001). The secretion from the labial gland (saliva) is known to have diverse species-specific functions, including intraspecific communication, nest construction and regulation of nest climate as a fungicide, social nutrient, source of digestive enzymes, food-marking pheromone and also in defensive role (Noirot, 1969; Hewitt et al., 1971; Martin and Martin, 1978; Veivers et al., 1982; Hogan et al., 1988; Kaib and Ziesmann, 1992; Sillam-Dusses et al., 2012; Cheng, 2013). Glucose was identified as one of the compounds in the labial gland on thorax of 15 termites species from all over the world apart from inositol, arbutin and hydroquinone (Reinhard et al., 2002a). It proved that glucose is desired as a multifunctional nutrient in dietary of termite besides acting as energy sources in termite body. The positive feeding response to glucose in this study indicated that glucose showed some potential as a phagostimulant for the development of palatable baits for termites, notably C. curvignathus.
According to Higashi et al. (1992) and Ding et al. (2015), termite foods are lack of nitrogen and that termites depend on intestinal bacteria for nitrogen fixation for survival (Slaytor and Chappell, 1994). The percentage of nitrogen relative to carbon in termites is around 10-20% whereas the woody and herbaceous tissues are 0.2-4% (La Fage and Nutting, 1978). Nitrogen source is crucial for production of proteins and influences termite behaviour such as food choice (Prestwich et al., 1980; Pearce, 1997) and nesting preference (Shellman-Reeve, 1994) in several termite species. Due to limitation of nitrogen content in natural food resources, nitrogen additives are believed able to increase food consumption of termites. In current study, urea is the most preferable nitrogen sources by C. curvignathus compared to yeast extract and peptone. Urea solutions was also proven palatable to termites in choice feeding tests of Reticulitermes flavipes and R. virginicus but high dietary N in termite food can cause defaunation, whereby higher concentration (5.0% w/v) of urea would adversely affect the protozoan populations as well and further lowering the survivor (Waller, 1996). Cannibalism and entombment will also occur when termites are nutritionally stressed (Pearce, 1997).
However, there was no significant difference in the combination of urea and glucose which might be due to both being additive having the same overwhelming phagostimulating effect (Reinhard and Kaib, 2001). It might be that these nutrients have to be added at higher concentrations to observe the effect. In the glucose percentage test, different concentrations of glucose were tested because termite feeding behaviour may alter even in a small change in concentration (Haifig et al., 2010). Waller and Curtis (2003) found that termites preferred a weak sugar concentration, whereby R. flavipes and R. virginicus consumed significantly more filter paper treated with 1 or 3% (wt:vol) sugar solutions (glucose, xylose and sucrose) than water-treated paper in choice tests. In addition, the study of Castillo et al. (2013) had indicated that C. curvignathus consumed more bait matrices comprising of cellulose, 3% glucose and casava. Current study proved again with similar result that C. curvignathus showed preference on filter paper discs supplemented with 1 and 3% glucose. Since both concentrations showed no statistically significant difference in termite consumption, this suggested that 1% of glucose is more applicable to use as additive in termite bait in order to minimize the cost.
The additive and synergistic effect of incorporation of glucose in rubber wood sawdust bait indicated that the formulated baits have the potential to become subterranean termite management tool in agriculture. The mixing of rubber wood sawdust with Metarhizium spores which was in powder form had increased the possibility of termites body contact with fungus spores when foraging and consuming at the treated area. The forager termite workers were contaminated most likely around the mouth part, abdomen and legs. The grooming behaviour of termites caused further pathogen infection transmission among the nestmates. Addition of glucose into the formulated bait had disguise or make them non-repellent against termites. Glucose probably serve as a masking agent for concealing the presence of fungal volatiles that are repellents to termites from detection by antennal olfactory of termite Balachander et al. (2013) and increases the palatability of the bait. The lower sensitivity of termites on foreign organisms favours the fungal spores for sufficient penetration sites and time to penetrate into termite cuticle which makes the termites susceptible to disease attack. The antagonistic effect of combination of glucose with 0.05 mg a.i L–1 fipronil and 107 conidia g–1 bait is difficult to hypothesize as combination of 0.05 mg a.i L–1 fipronil and 107 conidia g–1 bait without glucose had obtained the highest synergistic effect in the preliminary study. One of the possibilities may be that the addition of glucose could not overwhelm the synergy effect of 0.05 mg a.i L–1 fipronil and 107 conidia g–1 bait without glucose which caused higher termite mortalities in the bioassay.
Generally, three species termite of genus Coptotermes were found in oil palm plantations on peatland, i.e., C. curvignathus, C. borneensis and C. sepangensis (Kon et al., 2012). Currently, only the fresh wood eater of species C. curvignathus was identified as the main culprit that caused damage and palm death by feeding living plant tissue (Chan et al., 2011). There was no study yet regarding C. borneensis and C. sepangensis as pest to oil palm trees even both termite species were also discovered on the oil palm tree (Kon et al., 2012). Several studies had proved that termites have species-species preferences (Haifig et al., 2010). Thus, the bait was formulated based on preference of the target species C. curvignathus in this study i.e., glucose. However, glucose is a very common nutrient that can be found in natural food resources for termites such as wood and decay plant materials but vary in concentration. Glucose is presumably preferred by other termite species as well which are not pest in monoculture oil palm plantations. In this case, C. curvignathus is the largest in size over all oriental Coptotermes species (Thapa, 1981) and very aggressive (Lim and Silek, 2001), whereby C. curvignathus were observed to be able to occupy the food sources from other termites as their possession. Hence, application of glucose formulated bait on C. curvignathus infested palms or bait stations would be the best way to avoid indiscriminate killing of non-pest termite species.
CONCLUSION
In conclusion, knowledge of termite behaviour is essential before formulating a bait. The feeding preference test demonstrated that bait size and nutrients supplemented in bait play a role in food selection of termites. Rubber wood in sawdust form is effective in increasing termite feeding consumption. Besides, glucose might be a key additive in development of an effective bait system for subterranean termite management. Inclusion of low concentration of glucose into rubber sawdust bait with chemical and biological control agents can improve the attractiveness and palatability of bait matrices among termites and is believed to eliminate the colonies ultimately. Further studies on field trial are necessary to confirm the efficacy of bait in natural environment.
ACKNOWLEDGMENTS
This publication was funded by The Ministry of Higher Education (MOHE) Malaysia and also supported by Research University Grant Scheme (RUGS) UPM Project No. 01-01-11-1128RU.
REFERENCES
- Balachander, M., O.K. Remadevi and T.O. Sasidharan, 2013. Dissemination of Metarhizium anisopliae infection among the population of Odontotermes obesus (Isoptera: Termitidae) by augmenting the fungal conidia with attractants. J. Asia-Pac. Entomol., 16: 199-208.
CrossRefDirect Link - Bernklau, E.J., E.A. Fromm, T.M. Judd and L.B. Bjostad, 2005. Attraction of subterranean termites (Isoptera) to carbon dioxide. J. Econ. Entomol., 98: 476-484.
CrossRefDirect Link - Broadbent, S., M. Farr, E.J. Bernklau, M.S. Siderhurst, D.M. James and L.B. Bjostad, 2006. Field attraction of termites to a carbon dioxide-generating bait in Australia (Isoptera). Sociobiology, 48: 1-9.
Direct Link - Castillo, V.P., A.S. Sajap and M.H. Sahri, 2013. Feeding response of subterranean termites Coptotermes curvignathus and Coptotermes gestroi (Blattodea: Rhinotermitidae) to baits supplemented with sugars, amino acids and cassava. J. Econ. Entomol., 106: 1794-1801.
CrossRefPubMedDirect Link - Chan, S.P., C.F.J. Bong and W.H. Lau, 2011. Damage pattern and nesting characteristic of Coptotermes curvignathus (Isoptera: Rhinotermitidae) in oil palm on peat. Am. J. Applied Sci., 8: 420-427.
CrossRefDirect Link - Chantuma, P., A. Lacointe, P. Kasemsap, S. Thanisawanyangkura and E. Gohet et al., 2009. Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping. Tree Physiol., 29: 1021-1031.
CrossRefDirect Link - Chen, J. and G. Henderson, 1996. Determination of feeding preference of Formosan subterranean termite (Coptotermes formosanus Shiraki) for some amino acid additives. J. Chem. Ecol., 22: 2359-2369.
CrossRefDirect Link - Cheng, D.B., 2013. Progress in termite pheromones. Acta Entomologica Sinica, 56: 419-426.
Direct Link - Cornelius, M.L. and W.L. Osbrink, 2008. Effect of bait supplements on the feeding and tunneling behavior of the Formosan subterranean termite (Isoptera: Rhinotermitidae). Sociobiology, 51: 497-511.
Direct Link - Cornelius, M.L., D.J. Daigle, W.J. Connick Jr., A. Parker and K. Wunch, 2002. Responses of Coptotermes formosanus and Reticulitermes flavipes (Isoptera: Rhinotermitidae) to three types of wood rot fungi cultured on different substrates. J. Econ. Entomol., 95: 121-128.
CrossRefDirect Link - Evans, T.A. and N. Iqbal, 2015. Termite (order Blattodea, infraorder Isoptera) baiting 20 years after commercial release. Pest Manage. Sci., 71: 897-906.
CrossRefDirect Link - Evans, T.A., J.C.S. Lai, E. Toledano, L. McDowall, S. Rakotonarivo and M. Lenz, 2005. Termites assess wood size by using vibration signals. Proc. Natl. Acad. Sci. USA., 102: 3732-3737.
CrossRefDirect Link - Evans, T.A., R. Inta, J.C.S. Lai and M. Lenz, 2007. Foraging vibration signals attract foragers and identify food size in the Drywood termite, Cryptotermes secundus. Insectes Sociaux, 54: 374-382.
CrossRefDirect Link - Getty, G.M., C.W. Solek, R.J. Sbragia, M.I. Haverty and V.R. Lewis, 2007. Large-scale suppression of a subterranean termite community using the Sentricon® Termite Colony Elimination System: A case study in Chatsworth, California, USA. Sociobiology, 50: 1041-1050.
Direct Link - Haifig, I., F.F. Marchetti and A.M. Costa-Leonardo, 2010. Nutrients affecting food choice by the pest subterranean termite Coptotermes gestroi (Isoptera: Rhinotermitidae). Int. J. Pest Manage., 56 : 371-375.
CrossRefDirect Link - Henson, I.E., K.C. Chang, M.S.N. Aishah, S.H. Chai, M.Y. Hasnuddin and A. Zakaria, 1999. The oil palm trunk as a carbohydrate reserve. J. Oil Palm Res., 11: 98-113.
Direct Link - Hewitt, P.H., J.J.C. Nel and I. Schoeman, 1971. Influence of group size on water imbibition by Hodotermes mossambicus alate termites. J. Insect Physiol., 17: 587-600.
CrossRefDirect Link - Higashi, M., T. Abe and T.P. Burns, 1992. Carbon-nitrogen balance and termite ecology. Proc. R. Soc. London B: Biol. Sci., 249: 303-308.
CrossRefDirect Link - Hoe, P.K., C.F.J. Bong, K. Jugah and A. Rajan, 2009. Evaluation of Metarhizium anisopliae var. anisopliae (Deuteromycotina: Hyphomycete) isolates and their effects on subterranean termite Coptotermes curvignathus (Isoptera: Rhinotermitidae). Am. J. Agric. Biol. Sci., 4: 289-297.
CrossRefDirect Link - Hogan, M., P.C. Veivers, M. Slaytor and R.T. Czolij, 1988. The site of cellulose breakdown in higher termites (Nasutitermes walkeri and Nasutitermes exitiosus). J. Insect Physiol., 34: 891-899.
CrossRefDirect Link - Huang, Q.Y., C.L. Lei and D. Xue, 2006. Field evaluation of a fipronil bait against subterranean termite Odontotermes formosanus (Isoptera: Termitidae). J. Econ. Entomol., 99: 455-461.
CrossRefPubMedDirect Link - Inta, R., T.A. Evans, J.C.S. Lai and M. Lenz, 2007. What do vibrations have to do with termites' food choice? Acoustics Aust., 35: 73-77.
Direct Link - Kaib, M. and J. Ziesmann, 1992. The labial gland in the termite Schedorhinotermes lamanianus (Isoptera: Rhinotermitidae): Morphology and function during communal food exploitation. Insectes Sociaux, 39: 373-384.
CrossRefDirect Link - Kasseney, B.D., T. Deng and J. Mo, 2011. Effect of wood hardness and secondary compounds on feeding preference of Odontotermes formosanus (Isoptera: Termitidae). J. Econ. Entomol., 104: 862-867.
CrossRefPubMedDirect Link - Kon, T.W., C.F. Bong, J.H. King and C.T.S. Leong, 2012. Biodiversity of termite (Insecta: Isoptera) in tropical peat land cultivated with oil palms. Pak. J. Biol. Sci., 15: 108-120.
CrossRefPubMedDirect Link - Martin, M.M. and J.S. Martin, 1978. Cellulose digestion in the midgut of the fungus-growing termite Macrotermes natalensis: The role of acquired digestive enzymes. Science, 199: 1453-1455.
CrossRefDirect Link - Ngee, P.S., A. Tashiro, T. Yoshimura, Z. Jaal and C.Y. Lee, 2004. Wood preference of selected Malaysian subterranean termites (Isoptera: Rhinotermitidae, Termitidae). Sociobiology, 43: 535-550.
Direct Link - Prestwich, G.D., B.L. Bentley and E.J. Carpenter, 1980. Nitrogen sources for Neotropical nasute termites: Fixation and selective foraging. Oecologia, 46: 397-401.
CrossRefDirect Link - Rath, A.C., 2000. The use of entomopathogenic fungi for control of termites. Biocontrol Sci. Technol., 10: 563-581.
CrossRefDirect Link - Reinhard, J. and M. Kaib, 2001. Thin-layer chromatography assessing feeding stimulation by labial gland secretion compared to synthetic chemicals in the subterranean termite Reticulitermes santonensis. J. Chem. Ecol., 27: 175-187.
CrossRefDirect Link - Reinhard, J., M.J. Lacey, F. Ibarra, F.C. Schroeder, M. Kaib and M. Lenz, 2002. Hydroquinone: A general phagostimulating pheromone in termites. J. Chem. Ecol., 28: 1-14.
CrossRefPubMedDirect Link - Saran, R.K. and M.K. Rust, 2005. Feeding, uptake and utilization of carbohydrates by western subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol., 98: 1284-1293.
CrossRefPubMedDirect Link - Shellman-Reeve, J.S., 1994. Limited nutrients in a dampwood termite: Nest preference, competition and cooperative nest defence. J. Anim. Ecol., 63: 921-932.
CrossRefDirect Link - Sillam-Dusses, D., J. Krasulova, V. Vrkoslav, J. Pytelkova and J. Cvacka et al., 2012. Comparative study of the labial gland secretion in termites (Isoptera). PLoS ONE, Vol. 7.
CrossRefDirect Link - Slaytor, M. and D.J. Chappell, 1994. Nitrogen metabolism in termites. Comp. Biochem. Physiol. Part B: Comp. Biochem., 107: 1-10.
CrossRefDirect Link - Su, N.Y., 2005. Directional change in tunneling of subterranean termites (Isoptera: Rhinotermitidae) in response to decayed wood attractants. J. Econ. Entomol., 98: 471 -475.
CrossRefPubMedDirect Link - Veivers, P.C., A.M. Musca, R.W. O'Brien and M. Slaytor, 1982. Digestive enzymes of the salivary glands and gut of Mastotermes darwiniensis. Insect Biochem., 12: 35-40.
CrossRefDirect Link - Waller, D.A., 1996. Ampicillin, tetracycline and urea as protozoicides for symbionts of Reticulitermes flavipes and R. virginicus (Isoptera: Rhinotermitidae). Bull. Entomol. Res., 86: 77-81.
CrossRefDirect Link - Waller, D.A. and A.D. Curtis, 2003. Effects of sugar-treated foods on preference and nitrogen fixation in Reticulitermes flavipes (Kollar) and Reticulitermes virginicus (Banks) (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am., 96: 81-85.
CrossRefDirect Link