
Eko Kuswanto
School of Life Sciences and Technology, Institut Teknologi Bandung, Jalan Ganesha 10, Bandung, 40132, Indonesia
Intan Ahmad
School of Life Sciences and Technology, Institut Teknologi Bandung, Jalan Ganesha 10, Bandung, 40132, Indonesia
Ramadhani E. Putra
School of Life Sciences and Technology, Institut Teknologi Bandung, Jalan Ganesha 10, Bandung, 40132, Indonesia
Idham S. Harahap
Bogor Agricultural University, Jalan Dramaga 1 Bogor 16610, Indonesia
Journal of Entomology
Year: 2015 | Volume: 12 | Issue: 2 | Page No.: 87-94







ABSTRACT
Factors that cause individuals to recognize nestmates and non-nestmates in the same termite species require ongoing research. Chemical communication between individuals is predicted to be a key factor in the recognition behavior. This study aims to determine the level of aggression intraspecies Macrotermes gilvus and identify chemical components that play a role in influencing the behavior. Agonistic test intra- and intercolony M. gilvus that separated distance far away and a GC-MS analysis to determine of volatile compounds which role communication among individuals M. gilvus have been done. The research results showed that no aggression behavior intraspecies M. gilvus both intra- and intercolony. Agonistic test on three termite colonies M. gilvus from Bandung, Bogor and Bandar Lampung within maximum distance 216 km showed no agonistic. Agonistic level in intracolony only happens antennation or examination, while in intercolony occurs antennation, examination, alarm behavior and avoidance one another. Aggressive behavior (as a marker of agonistic behavior) does not appear both inter- and intracolony. Identification of the volatile compounds that assumed to play a role as, a key of intraspecific colony recognition has been carried out using GC-MS analysis, the results are 22 volatile compounds of caste workers and soldiers (both minor- and major soldier). Two novel volatile compounds has been identified i.e., (Z)-6-octadecenoic acid and (E)-9-octadecenoic acid, respectively, as the key of individuals for recognizing both non- and nestmembers of M. gilvus. Those compounds are species-specific on M. gilvus.
PDF Abstract XML References Citation
Received: June 29, 2015;
Accepted: August 18, 2015;
Published: August 25, 2015
How to cite this article
Eko Kuswanto, Intan Ahmad, Ramadhani E. Putra and Idham S. Harahap, 2015. Two Novel Volatile Compounds as the Key for Intraspecific Colony Recognition in Macrotermes gilvus (Isoptera: Termitidae). Journal of Entomology, 12: 87-94.
DOI: 10.3923/je.2015.87.94
URL: https://scialert.net/abstract/?doi=je.2015.87.94
DOI: 10.3923/je.2015.87.94
URL: https://scialert.net/abstract/?doi=je.2015.87.94
INTRODUCTION
Profile of hydrocarbons has been stated as the species-specific and may explain the difference of intercolony and can be used to express a genetic relationships between individuals (Haverty et al., 1988; Bagine et al., 1994; Howard et al., 1988; Haverty et al., 1999, 2005; Batista-Pereira et al., 2004). Cuticular hydrocarbons are prime candidates for recognition of the colony in social insects than morphological, genetic and geographic distances (Kaib et al., 2004).
Cuticular hydrocarbon variation analysis, according to Kaib et al. (2004), in species Macrotermes subhyalinus in West Africa (Comoe National Park) on a short distance (<1 km) showed four chemically different phenotypes, also found a significant correlation between the differences hydrocarbons composition and intercolony agression. The other side, Neoh et al. (2012) stated that agonistic behavior does not occur in major workers individuals of M. gilvus despite coming from different colonies and distance apart (sites ranged from 6-18 km).
It is necessary to further research on the chemical components involved to explain the link between the different colonies (spatially) and a low agression nature (even no agression) intraspecies M. gilvus in the range colonies more distances (156-210 km). This research is done to determine the level of agression behaviors both intra- and intercolony M. gilvus and identify the volatile components, which is the key recognition individuals intraspecies M. gilvus.
MATERIALS AND METHODS
Source colonies and maintenance in the laboratory: Three colonies of termites M. gilvus obtained from three different locations: Bandung, Bogor and Bandar Lampung, in July, 2014. Distance Bogor-Bandar Lampung is 156 km, Bandung-Bogor 164 km and Bandung-Bandar Lampung 210 km. Three nests of three colonies were moved and maintained to the laboratory for one week before bioassay. Nest consists some of combs and the royal chambers in which there are both caste workers, soldiers (minor soldier and major soldier) and reproductive (queen, king and alates). Maintenance termites using sealed plastic containers (volume 5.6 L), which filled moistened sand on the bottom and tissue (Tessa TP-01) as, a food source for termites. Maintenance is performed in the incubation chamber at temperature 28±2°C and humidity >85%.
Intra and intercolony agonistic test: Individuals that are used to test agonistic are major workers, minor soldiers and major soldiers from those three colonies. The testing arena, modified by Wong and Lee (2010) and Neoh et al. (2012), using a filter paper disc (diameter 5 cm) moistened with distilled water was placed in a 5-cm-diameter plastic dish. The arena was equally divided into two areas by a plastic card. Termites in the ratio 1: 1 (five individuals from each colony) was tested with a combination of workers versus workers, workers versus minor soldiers, workers versus major soldiers, minor soldiers versus minor soldiers and minor soldiers versus major soldiers. Termites from one colony in a pair of workers versus workers, minor soldiers versus minor soldiers and major soldiers versus major soldiers were marked with a dot of white paint on the head capsule to distinguish them from the same individuals of the other colony. Termites were acclimatized in the test arena for 5 min before the separating plastic card was removed. The behaviors were video recorded during encounters (Ricoh WG-4) for 5 min. The videos were reviewed and analyzed for agonistic reactions. The agression behaviors were leveled from 1-3 based on the definition described by Jmhasly and Leuthold (1999) with slight modifications of Neoh et al. (2012): (1) examination or antennation: contact between antenna, head or body (it means no agonistic behavior), (2) alarm reaction or avoidance, jerking, chasing and/or being chased (it means no agonistic behavior) and (3) agression: mandibles open, seizing or biting (it means no agonistic behavior). The agression behaviors that occur during interactions for each 10 sec intervals were recorded and a total of 30 observations for each test was made to scale and averaged from five replicates were there.
Chemical analysis of volatile compounds: For volatile sampling used live animals, such as; the method of Matsuura et al. (2010) and Himuro et al. (2011). A total of 20 individuals of major workers each colonies from Bogor (code WB), Bandung (code WD) and Bandar Lampung (code WL), then 20 individuals of minor soldiers of Bandung's colony (code MSD) and Bandar Lampung's colony (code MSL), also 10 individuals of major soldiers from Bogor (code JSB) and Bandar Lampung (code JSL), those seven of the samples were washed for 15 min in 100 mL of n-hexane. Then 1 μL sample leaching results were analyzed using gas chromatography. Gas Chromatography-Mass Spectrophotometer (GC-MS) analyzes of volatiles were performed on Hewlett Packard Agilent 5890, equipped with a capillary column HP-1 column (30 m long×250 μm phase thickness×0.25 μm diameter column). Column temperature was raised from 40°C (5 min hold) at 10°C min–1 to 280°C. The injection was splitless, with helium as the carrier gas (1.5 mL min–1). The MS data was obtained under the following conditions: ionization current, 100 μA; ionization energy, 70 eV; accelerating voltage, 1470.6V; scan range of 40-450 m/z. The gas chromatographic and mass spectrophotometer system were both controlled with a data processing system. Compounds were identified from comparisons of mass spectra with a National Institute of Standards and Technology (NIST) data library system.
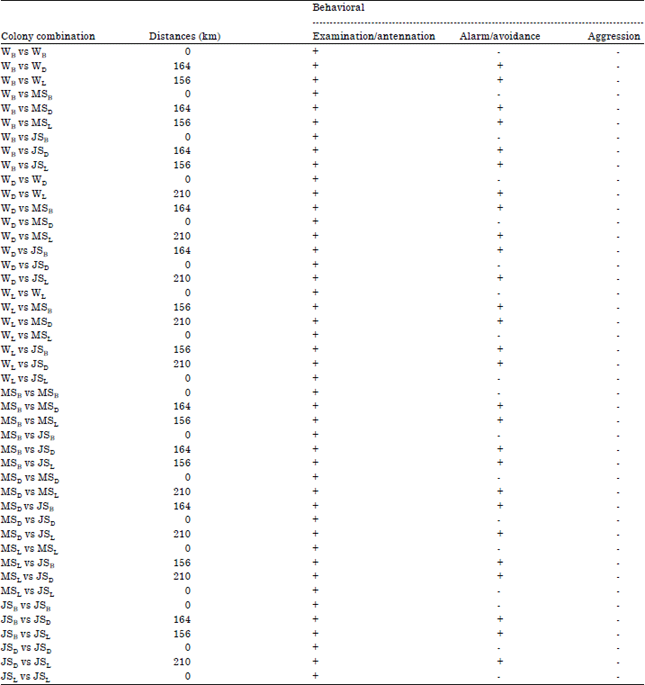
Data analyses: Quantitative behavioral test with total 45 combination, i.e., 18 intracolony combinations and 27 intercolony combinations (Table 1), was executed in July, 2014. The agression behaviors that occur during interactions, such as; examination and antennation (level 1), alarm reaction and avoidance (level 2) and agression (level 3) were observed. If the behavior on each level occurred ≥80% in any combination then it is expressed as, + means existing and if, this is only ≤20% in any combination then expressed as means no existing.
RESULTS AND DISCUSSION
In general, no agonistic happened intraspecies M. gilvus. Caste differences both inter- and intracolony not indicated agonistic behavior, as well as the distance (ranged from 156-210 km) was not significantly to describe about intraspecific on M. gilvus. Combination treatment between workers versus workers or workers versus minor soldiers, or workers versus major soldiers, or minor soldier versus minor soldiers and minor soldier versus major soldier did not show any agonistic behavior in the species M. gilvus. M. gilvus termite colony from Bandung, Bogor, Bandar Lampung with the closest distance 0 km (intracolony) and the longest 210 km (intercolony) did not show any agonistic behavior. No agonistic behavior in M. gilvus presented in Table 1.
The scale of agonistic behavior only at the level of alarm/avoidance in the different colonies distance and at the level of examination/antennation on intracolony while the level of agression, which is a sign of agonistic never occurred in all treatment combinations both intra- and intercolony in intraspecies M. gilvus. This is consistent with the results of research Neoh et al. (2012) that a combination of worker versus worker intra- and intercolony showed very low intraspecific aggression on M. gilvus, whereas in M. carbonarius showed little agression towards non-nestmates. According Kaib et al. (2004), colony recognition in Macrotermes intraspecies not influenced by morphology, genetics, geography or distance significantly. This is also corroborated by Jmhasly and Leuthold (1999) that aggressive behavior and an examination on major worker M. subhyalinus and M. bellicosus not correlated with spatial distance intercolony, also not correlated with differences in the volume of the nest and colony age involved. Shelton and Grace (1997) also stated, the same thing that the geographic separation of termite colonies was not a factor in predicting agonism on Coptotermes formosanus. Low agonistic response not only happened in intraspecies Macrotermes spp. but also occurs on Reticulitermes spp. (Grace, 1996; Delphia et al., 2003; Harahap et al., 2005), on Microcerotermes crassus (Wong and Lee, 2010) and on Coptotermes spp. (Uchima and Grace, 2009; Shelton and Grace, 1997; Cornelius and Osbrink, 2003).
| Table 1: | Behavioral existence of major workers resulting from different colony combination in Macrotermes gilvus |
 | |
Of 22 volatile components were identified, it can be specified that the six main volatile components derived from termites workers M. gilvus, 11 main volatile components of minor soldier and 15 main volatile components of major soldier. Compounds detected by GC-MS analysis presented in Table 2.
There are two main volatile components are believed to play an important role in the non-agonistic behavior on M. gilvus, namely (Z)-6-octadecenoic acid and (E)-9-octadecenoic acid. Component (Z)-6-octadecenoic acid is contained in the worker caste of termites M. gilvus from all three colonies and also at major soldiers of Bogor.
| Table 2: | Compounds detected by GC-MS analysis from worker, minor soldier, major soldier of M. gilvus |
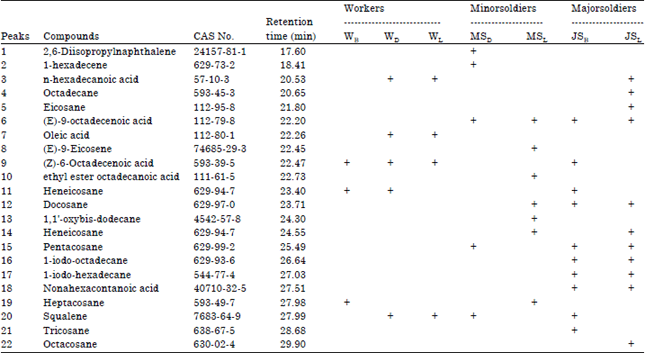 | |
| WB, JSB: Colony from Bogor, WD, MSD: Colony from Bandung, WL, MSL, JSL: Colony from bandar lampung, +: Identified, -: Not identified | |
While (E)-9-octadecenoic acid specifically found on the soldier caste both minor- and major soldiers in all colonies.
This is a new novelty and can explain about the components that play a role in nestmate and non-nestmate recognition intraspecies M. gilvus. A similar study of volatile compounds ever done Matsuura et al. (2010) and Himuro et al. (2011) on the species Reticulitermes speratus and Nasutitermes takasagoensis. Volatile compounds produced specifically queen on Reticulitermes speratus and Nasutitermes takasagoensis influence the behavior and physiology of colony members, these compounds have been identified by them i.e., n-butyl-n-butyrate and 2-methyl-1-butanol (in R. speratus) and phenylethanol (in N. takasagoensis), respectively.
Compounds (Z)-6-octadecenoic acid and (E)-9-octadecenoic acid has a formula C18H34O2 with molecular weight 282.46 (Fig. 1 and 2). Both chemicals are thought to be a component of chemical messages in communication intra- and intercolony on termites M. gilvus. As explained Richard and Hunt (2013), also Trhlin and Rajchard (2011) that the chemical message is the primary mode of communication for the majority of intra-social insect species. Previous research as did by Stuart (1961), Smythe and Coppel (1966), Matsumura et al. (1968) and Tokoro et al. (1994) also began to assert that the pheromone trail markers on the various types of termites have been identified and played a major role in the introduction of the colony and kin recognition that influence the behavior of the colony in general in terms of sourcing food and defend the colony from predators. Pheromone trail marker or recruitment has been discovered in the 1960’s, when three substances has been identified successfully. The first is the (E)-6-cembrene A of Nasutitermes exitiosus (Moore, 1966). The substance is not specific to the species because that is also found in N. walker and N. graveolus (Moore, 1966) and in Trinervitermes bettonianus (McDowell and Oloo, 1984). Then (3Z,6Z,8E)-3,6,8-dodecatrienol first identified as a pheromone trail marker of Reticulitermes virginicus (Matsumura et al., 1968; Tai et al., 1969).
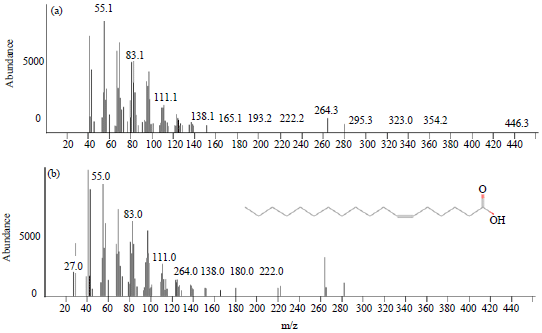 | |
| Fig. 1(a-b): | Mass spectra of (Z)-6-Octadecenoic acid from workers and soldiers of M. gilvus (top) and from a commercial sources (bottom) |
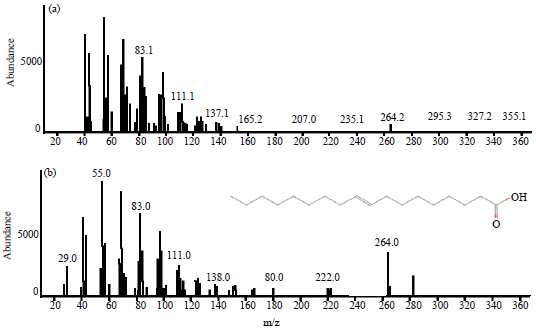 | |
| Fig. 2(a-b): | Mass spectra of (E)-9-Octadecenoic acid from soldiers of M. gilvus (top) and from a commercial sources (bottom) |
A novelty of volatile compounds in M. gilvus as an important chemical component in a chemical message intraspecies M. gilvus expected to expand further research on how to reset the definition of a colony in M. gilvus or may be in termite generally. Colony, which is known as a unique organization that does not apply to M. gilvus and perhaps more appropriately categorized as, a separate colony as supracolony, where a great distance but still occurs recognition colony members as well as, in intracolony.
CONCLUSION
Agonistic behavior is closely related to the individual chemical components issued in the colony to recognize members of intra- and intercolony intraspecific in Macrotermes gilvus. The absence of agonistic in M. gilvus both intra- and intercolony associated with volatile compounds, which are owned by the workers and soldiers. Both the main volatile compound is 6-octadecenoic acid, (Z) -and 9-octadecenoic acid, (E), respectively, which are owned by individual workers and soldiers of M. gilvus be key intra- and intercolony recognition in the same species.
ACKNOWLEDGMENT
The authors would like to thanks to the Academy of Analytical Chemistry (Politeknik AKA), Bogor. for facilitating equipments for GC-MS, also thanks to the Ministry of Religious Affairs, Republic of Indonesia for their financial support by providing the scholarship study No. Dj.I/Dt.I.IV/3/HN.01/1182/2010.
REFERENCES
- Bagine, R.K.N., R. Brandl and M. Kaib, 1994. Species delimitation in Macrotermes (Isoptera: Macrotermitidae): Evidence from epicuticular hydrocarbons, morphology and ecology. Ann. Entomol. Soc. Am., 87: 498-506.
CrossRefDirect Link - Batista-Pereira, L.G., M.G. dos Santos, A.G. Correa, J.B. Fernandes and A. Arab et al., 2004. Cuticular hydrocarbons of Heterotermes tenuis (Isoptera: Rhinotermitidae): Analyses and electrophysiological studies. Zeitsschrift Naturforschung, 59: 135-139.
Direct Link - Cornelius, M.L. and W.L.A. Osbrink, 2003. Agonistic interactions between colonies of the formosan subterranean termite (Isoptera: Rhinotermitidae) in New Orleans, Louisiana. Environ. Entomol., 32: 1002-1009.
Direct Link - Delphia, C.M., K.A. Copren and M.I. Haverty, 2003. Agonistic behavior between individual worker termites from three cuticular hydrocarbon phenotypes of Reticulitermes (Isoptera: Rhinotermitidae) from Northern California. Ann. Entomol. Soc. Am., 96: 585-593.
Direct Link - Grace, J.K., 1996. Absence of overt agonistic behavior in a northern population of Reticulitermes flavipes (Isoptera: Rhinotermitidae). Sociobiology, 28: 103-110.
Direct Link - Harahap, I.S., E.P. Benson, P.A. Zungoli, P.H. Adler and H.S. Hill, 2005. Inter- and intra-colony agonistic behavior of native subterranean termites, Reticulitermes flavipes and Reticulitermes virginicus (Isoptera: Rhinotermitidae). Sociobiology, 46: 305-316.
Direct Link - Haverty, M.I., K.A. Copren, M.G. Getty and V.R. Lewis, 1999. Agonistic behavior and cuticular hydrocarbon phenotypes of colonies of Reticulitermes (Isoptera: Rhinotermitidae) from Northern California. Ann. Entomol. Soc. Am., 92: 269-277.
CrossRefDirect Link - Haverty, M.I., M. Page, L.J. Nelson and G.J. Blomquist, 1988. Cuticular hydrocarbons of dampwood termites, Zootermopsis: Intra- and intercolony variation and potential as taxonomic characters. J. Chem. Ecol., 14: 1035-1058.
CrossRefDirect Link - Haverty, M.I., R.J. Woodrow, L.J. Nelson and J.K. Grace, 2005. Identification of termite species by the hydrocarbons in their feces. J. Chem. Ecol., 31: 2119-2151.
CrossRefDirect Link - Himuro, C., T. Yokoi and K. Matsuura, 2011. Queen-specific volatile in a higher termite Nasutitermes takasagoensis (Isoptera: Termitidae). J. Insect Physiol., 57: 962-965.
CrossRefDirect Link - Howard, R.W., B.L. Thorne, S.C. Levings and C.A. McDaniel, 1988. Cuticular hydrocarbons as chemotaxonomic characters for Nasutitermes corniger (Motschulsky) and N. ephratae (Holmgren) (Isoptera: Termitidae). Ann. Entomol. Soc. Am., 81: 395-399.
CrossRefDirect Link - Jmhasly, P. and R.H. Leuthold, 1999. Intraspecific colony recognition in the termites Macrotermes subhyalinus and Macrotermes bellicosus (Isoptera, Termitidae). Insectes Sociaux, 46: 164-170.
CrossRefDirect Link - Kaib, M., P. Jmhasly, L. Wilfert, W. Durka and S. Franke et al., 2004. Cuticular hydrocarbons and aggression in the termite Macrotermes subhyalinus. J. Chem. Ecol., 30: 365-385.
CrossRefDirect Link - Matsumura, F., H.C. Coppel and A. Tai, 1968. Isolation and identification of termite trail-following pheromone. Nature, 219: 963-964.
CrossRefDirect Link - Matsuura, K., C. Himuro, T. Yokoi, Y. Yamamoto, E.L. Vargo and L. Keller, 2010. Identification of a pheromone regulating caste differentiation in termites. Proc. Natl. Acad. Sci., 107: 12963-12968.
CrossRefDirect Link - McDowell, P.G. and G.W. Oloo, 1984. Isolation, identification and biological activity of trail-following pheromone of termite Trinervitermes bettonianus (Sjostedt) (Termitidae:Nasutitermitinae). J. Chem. Ecol., 10: 838-851.
CrossRefDirect Link - Moore, B.P., 1966. Isolation of the scent-trail pheromone of an Australian termite. Nature, 211: 746-747.
CrossRefDirect Link - Neoh, K.B., Y. Indiran, M. Lenz and C.Y. Lee, 2012. Does lack of intraspecific aggression or absence of nymphs determine acceptance of foreign reproductives in Macrotermes? Insectes Sociaux, 59: 223-230.
CrossRefDirect Link - Richard, F.J. and J.H. Hunt, 2013. Intracolony chemical communication in social insects. Insectes Sociaux, 60: 275-291.
CrossRefDirect Link - Wong, N. and C.Y. Lee, 2010. Intra- and interspecific agonistic behavior of the subterranean termite Microcerotermes crassus (Isoptera: Termitidae). J. Econ. Entomol., 103: 1754-1760.
Direct Link - Shelton, T.G. and J.K. Grace, 1997. Suggestion of an environmental influence on intercolony agonism of formosan subterranean termites (Isoptera: Rhinotermitidae). Environ. Entomol., 26: 632-637.
CrossRefDirect Link - Smythe, R.V. and H.C. Coppel, 1966. A preliminary study of the sternal gland of Reticulitermes flavipes (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am., 59: 1008-1010.
CrossRefDirect Link - Stuart, A.M., 1961. Mechanism of trail-laying in two species of termites. Nature, 189: 419-419.
CrossRefDirect Link - Tai, A., F. Matsumura and H.C. Coppel, 1969. Chemical identification of the trail-following pheromone for a southern subterranean termite. J. Organ. Chem., 34: 2180-2182.
CrossRefDirect Link - Trhlin, M. and J. Rajchard, 2011. Chemical communication in the honeybee (Apis mellifera L.): A review. Vet. Med., 56: 265-273.
Direct Link - Tokoro, M., M. Takahashi and R. Yamaoka, 1994. (Z,E,E)-dodecatrien-1-ol: A minor component of trail pheromone of termite, Coptotermes formosanus Shiraki. J. Chem. Ecol., 20: 199-215.
CrossRefDirect Link - Uchima, S.Y. and J.K. Grace, 2009. Interspecific agonism and foraging competition between Coptotermes formosanus and Coptotermes gestroi (Blattodea: Rhinotermitidae). Sociobiology, 54: 765-776.
Direct Link