
Hamdy K. Abou- Taleb
Plant Protection Research Institute, Agricultural Research Center, Sabahia, Alexandria, Egypt
Hossam El-Din M. Zahran
Department of Entomology, Faculty of Agriculture (El-Shatby), Alexandria University, Egypt
Abir A. Gad
Department of Entomology, Faculty of Agriculture (El-Shatby), Alexandria University, Egypt Department of Entomology, Faculty of Agriculture (El-Shatby), Alexandria University, Egypt
Journal of Entomology
Year: 2015 | Volume: 12 | Issue: 2 | Page No.: 77-86







ABSTRACT
Toxicity and effects of lufenuron and chlorfluazuron on some biochemical and physiological parameters of Spodoptera littoralis (Boisd.) larvae were assessed. Results showed no significant differences in toxicity of lufenuron and chlorfluazuron against 4th instar larvae with LC50 values 1.7 and 2.2 ppm, respectively. While, activity level of alanine aminotransferase (ALT), aspartate aminotransferase (AST) and alkaline phosphatase (ALP) increased, activity level of glutathione S-transferase (GST) showed adverse results after exposure to lufenuron and chlorfluazuron compared with control. Lufenuron at 0.62 ppm and chlorfluazuron at 0.76 ppm caused the highest increase in the AST activity (from 1.06 in control to 4.13 and 4.26 OD/mg protein/min, respectively) and ALT activity (from 1.02 in control to 5.22 and 5.50 OD/mg protein/min, respectively), after 72 h of exposure. In the case of GST, the highest enzyme activity inhibition was occurred by lufenuron at 0.28 and 0.62 ppm, where the inhibition percentage was 38.6 and 45.6, respectively. Results also revealed that, chlorfluazuron caused the highest decrease in total haemocyte count 49.26 and 53.89% at 0.31 and 0.76 ppm, respectively. Changes in the differential haemocyte population during lufenuron and chlorfluazuron treatments have been assessed. The percentage of Prohaemocytes (Pr) markedly decreased after treatment with both insecticides when compared with control but no significant difference was observed between different concentrations of tested insecticides. Moreover, the percentage of Granulocytes (Gr) and Plasmatocytes (Pl) decreased markedly after treatments. In contrast, Oenocytes (Oe) and Spherulocytes (Sph) increased after 72 h of lufenuron and chlorfluazuron treatment. Also, the Corpora Allata (CA) activity has been observed in S. littoralis treated larvae. Both lufenuron and chlorfluazuron decreased the CA surface area after 72 h of treatment compared to control.
PDF Abstract XML References Citation
Received: February 06, 2015;
Accepted: April 07, 2015;
Published: May 04, 2019
How to cite this article
Hamdy K. Abou- Taleb, Hossam El-Din M. Zahran and Abir A. Gad, 2015. Biochemical and Physiological Effects of Lufenuron and Chlorfluazuron on Spodoptera littoralis (Boisd.) (Lepidoptera: Noctuidae). Journal of Entomology, 12: 77-86.
DOI: 10.3923/je.2015.77.86
URL: https://scialert.net/abstract/?doi=je.2015.77.86
DOI: 10.3923/je.2015.77.86
URL: https://scialert.net/abstract/?doi=je.2015.77.86
INTRODUCTION
Cotton leafworm, Spodoptera littoralis (Boisd.) is one of the most destructive agricultural lepidopterous pests within its subtropical and tropical range. In Egypt, it can attack numerous economically important crops throughout the year (Hatem et al., 2009). The chemical control of S. littoralis has been extensively reported in relation especially to cotton in Egypt (Abo-El-Ghar et al., 1986). Extensive use of insecticides, multiple generations of CLW per annum and the availability of host crops, around the year, have contributed to the development of resistance in this pest to many insecticide groups (Abo Elghar et al., 2005; Abou-Taleb, 2010). Selective insecticides, such as Insect Growth Regulators (IGRs), are highly desirable in integrated and resistance pest management programs. They are advantageous because they do not persist long in the environment due to their rapid biodegradation and exhibit low toxicity for non-target organisms especially vertebrates (Zibaee et al., 2011).
The IGRs are compounds that interfere with growth of insects (Kai et al., 2009). They disrupt the normal activity of the endocrine system of insects, affecting development, reproduction or metamorphosis of the target insects. The IGRs include Juvenile Hormone (JH) mimics and Chitin Synthesis Inhibitors (CSIs) (Tunaz and Uygun, 2004). The CSIs, such as chlorfluazuron and lufenuron inhibit the production of chitin, a major component of the insect exoskeleton. By consideration of this information, the aim of this study is to investigate the interaction of such IGR compounds (lufenuron and chlorfluazuron) with some biochemical enzymatic components. Efficacy of these IGR compounds on the total hemocyte count, different hemocyte count and corpora allata activity were also carried out. These parameters have a strong relation with growth, development and ecdysis process of insect larvae. The biochemical basis of buffering in the haemolymph of insects has received little attention and the chemical complexity of insect haemolymph. Peel and Akam (2007) suggested that the relative importance of various compounds to buffering may be quite different from that in other arthropods or vertebrates. Gad and Abdel-Megeed (2006) studied the effects of spinosad and emamectin benzoate on the blood picture and DNA structure of the cotton leaf worm S. littoralis. However, this haemocyte population is not only affected by changes during the development process, but also by various forms of stress such as insecticides, parasitization and starvation (Essawy, 1991).
MATERIALS AND METHODS
Experimental insect: Laboratory strain cotton leafworm larvae used for testing program was reared for several years on castor bean leaves at 25±2°C and 65±5% RH as mentioned by Eldefrawi et al. (1964).
Tested insecticides: Lufenuron (Match®5%EC) and chlorfluazuron (Atabron®5%EC) were supplied by Syngenta.
Bioassay: Homogenous pieces of the castor oil leaves were dipped in a series of the insecticides concentrations for 10 sec held vertically to allow excess solution to drip off and dried at room temperature. Treated castor oil leaf pieces were transferred to a plastic cups and the appropriate number of (10 larvae in each cup) starved 4th instar (45±2 mg larva–1) larvae were added. Each concentration was replicated five times. Castor oil leaves were dipped in water for control treatment. Mortality percentages were recorded after 72 h of treatment, corrected according to Abbott equation (Abbott, 1925) and subjected to probit analysis. The median lethal concentrations, confidence limits and the slope were calculated (Finney, 1971).
Biochemical assays
Sample preparation for enzymatic assay: Starved laboratory strain 4th instar larvae of S. littoralis were fed on castor oil leaves dipped in LC10 and LC25 equivalent concentrations of lufenuron and chlorfluazuron solutions plus untreated larvae as a control. Suitable numbers of total larvae were collected after 24, 48 and 72 h of treatment. These larvae were homogenized (1: 10 w/v) in glass distilled water pH 7, using glass homogenizer. The homogenate was centrifuged at 5000 rpm for 30 min at 4°C using IEC-CRU 5000 cooling centrifuge. The supernatant was used for biochemical assays.
Assay of aspartate (AST) and alanine aminotransferases (ALT): Activity of AST and ALT was measured according to the method of Reitman and Frankel (1957) using Diamond Diagnostic kit (Diamond Co., Egypt). In this method 100 μL of enzyme source was added to 500 μL of 100 mM phosphate buffer pH 7.2 containing 80 mM L-aspartate as a substrate for AST or 80 mM D-L-alanine as a substrate for ALT and 4 mM α-ketoglutarate. This mixture was incubated for 30 min at 37°C. After that, 500 μL of developing color reagent, 4 mM 2,4-dinitrophenylhydrazine, was added and the solution was incubated for 20 min at room temperature. Lastly, 5 mL of 0.4 N NaOH was added then mixed and left at room temperature for 5 min. An assay mixture without enzyme source was used as the blank and the absorption was measured at 546 nm using spectrophotometer (Milton Roy Spectronic 601). The AST and ALT specific activities were determined as OD546/mg protein/min.
Assay of alkaline phosphatases (ALP): Activity of ALP was determined according to the method of DGKC (1972), using Diamond Diagnostic kit (Diamond Co., Egypt). In this method, 20 μL of the enzyme source was added to 1 mL diethanolamine buffer (0.9 M, pH 9.8) containing 0.6 mM magnesium ions and 1 mM p-nitrophenyl phosphate. Then, the mixture was mixed in the cuvette, incubated for 30 sec in the spectrophotometer (Milton Roy Spectronic 601), start stopwatch simultaneously and read again after exactly 1, 2 and 3 min at 405 nm. The ALP specific activity was determined as OD405/mg protein/min.
Assay of glutathione S-transferase (GST): The GST was determined by using CDNB as a substrate (Kao et al., 1989). The assay mixture consisted of 50 mM CDNB in 95% ethanol, 50 mM GSH and 10-20 mg of enzyme sample in 3 mL of 50 mM phosphate buffer (pH 7.5). The change in absorbance was measured at 340 nm for up to 3 min and the enzyme activity in terms of mmol of CDNB conjugated min–1 mg protein–1 was calculated using the extinction coefficient of 9.6 mM–1 cm–1.
Protein measurements: Protein estimation has been carried out according to Lowry et al. (1951) using folin reagent as a coloring reagent. The developed color was measured at 750 nm using Sequoia-Turner Model 340 spectrophotometer against the blank. The standard curve of protein was established using different concentrations of Bovine Serum Albumin (BSA) (10-100 μg μL–1).
Studies of haemocytes: The haemolymph samples were taken after 72 h of treatment with lufenuron and chlorfluazuron (at LC10 and LC25 equivalent values). The Total Haemocyte Count (THC) was carried out using the haemocytometer as reported according to method described by Jones (1962) and Essawy (1991). The Differential Haemocyte Count (DHC) was calculated using method of Akai and Sato (1973). To calculate the DHC, 100 cells were identified to their typical haemocyte type after staining a smear of haemolymph with Wright's stain (Essawy, 1990).
Corpora allata activity: Corpora Allata (CA) volume was used as an indicator of the Juvenile Hormone (JH) level (Pflugefelder, 1948). Larvae were dissected after 72 h of treatment. The method of Armstrong and Carr (1964) was used to calculate the CA surface area.
Statistical analysis: Data were analyzed statistically using analysis of variance (ANOVA) and differences among the means were determined for significance at 0.01 using LSD test (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
Toxicity of lufenuron and chlorfluazuron against 4th instar S. littoralis larvae: According to the LC50 values (Table 1), there is no significant difference between the toxicity of lufenuron (LC50 = 1.7 ppm) and chlorfluazuron (LC50 = 2.2 ppm) against the 4th instar larvae of S. littoralis after 72 h of exposure. The concentrations of 0.28 and 0.62 ppm were LC10 and LC25 for lufenuron and the concentrations of 0.31 and 0.76 ppm were LC10 and LC25 for chlorfluazuron. These concentrations were used to find their effects on enzymatic and non enzymatic components. These results are in agreement with Abdel-Rahman and Abou-Taleb (2007). They reported a comparable toxicity of lufenuron and chlorfluazuron against the 2nd instar larvae of S. littoralis.
Results of biochemical assays: Significant effects on alanine (ALT) and aspartate (AST) aminotransferases caused by lufenuron and chlorfluazuron were observed (Table 2). Both concentrations of lufenuron and chlorfluazuron increased the AST and ALT activity compared to control after 48 and 72 h of exposure. Lufenuron at 0.76 ppm and chlorfluazuron at 0.62 ppm caused the highest increase in the AST activity, where the activity was 4.13 and 4.26 (OD/mg protein/min) compared to 1.06 (OD/mg protein/min) in control after 72 h of treatment, respectively.
| Table 1: | Toxicity of lufenuron and chlorfluazuron against 4th instar larvae of S. littoralis after 72 h of exposure |
 | |
| Table 2: | In vivo effect of lufenuron and chlorfluazuron on AST and ALT activity in 4th instar larvae of S. littoralis after different exposure times |
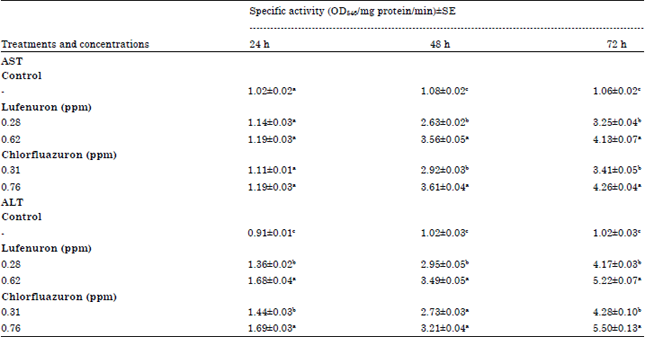 | |
| Numbers within the same column with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.05) | |
| Table 3: | In vivo effect of lufenuron and chlorfluazuron on ALP activity in 4th instar larvae of S. littoralis after different exposure times |
 | |
| Numbers within the same column with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.05) | |
In the case of ALT, the activity of the enzyme was 5.22 and 5.50 (OD/mg protein/min) compared to 1.02 (OD/mg protein/min) in control, at the same concentrations of lufenuron and chlorfluazuron, respectively, after 72 h of treatment (Table 2). The changes of AST and ALT activity in the S. littoralis larvae following lufenuron and chlorfluazuron exposures suggested that S. littoralis showed adaptive elevation in the activity of the two aminotransferase enzymes, thereby, probably aiding gluconeogenesis through transamination of glucogenic amino acids to meet the energy demand under lufenuron and chlorfluazuron toxicity. These data are in accordance with many results of other authors. Zibaee et al. (2011) mentioned that AST and ALT activity significantly increased in Eurygaster integriceps after exposure to pyriproxyfen. They concluded that possible damages of this insecticide to haemocytes and fat bodies are the reason in elevation of their activity. This correlation could be attributed to regeneration of haemocytes by hematopoietic organs and fat bodies that definitely needs to different amino acids prepared by transamination process. Abou-Taleb et al. (2009) recorded an increase in the AST and ALT activity of S. littoralis larvae as a result of emamectin benzoate treatment. In another study, Ramaswamy et al. (1999) reported that the activity of these enzymes was elevated when Sarotherodon mossambicus (Peters) had been exposed to carbaryl.
Alkaline phosphatases (APs) are classically described as homodimeric nonspecific metalloenzymes which catalyze phosphomonoesterase reactions (Trowsdale et al., 1990). They are responsible for removing phosphate groups from many types of molecules, including nucleotides, proteins and alkaloids in alkaline conditions under name of dephosphorylation (Zibaee and Bandani, 2010). Also, they have an additional role in the final steps of lipid digestion in insects (Klowden, 2007). In this study, treatment of S. littoralis 4th instar larvae by lufenuron and chlorfluazuron increased the activity of ALP after 48 and 72 h of exposure (Table 3). The highest increase in the ALP activity was achieved by chlorfluazuron at 0.76 ppm after 72 h of exposure, where the ALP activity was 199.5% of control (Table 3). While, Abou-Taleb et al. (2009) reported an inhibition of ALP activity of S. littoralis larvae after exposure to emamectin benzoate, Zibaee et al. (2011) reported a significant increase in E. integriceps ALP activity after exposure to pyriproxyfen. They mentioned that, Changes in ALP and ACP activities after treatment with pyriproxyfen in comparison with control indicate altering the physiological balance in nutritional and intermediate metabolism.
Glutathione S-transferases in insects play an important role in detoxification of insecticides (Clark, 1989). In this study, S. littoralis GST activity (μmol min–1 mg protein–1) was significantly decreased as a result of exposure to lufenuron and chlorfluazuron compared to control (Table 4).
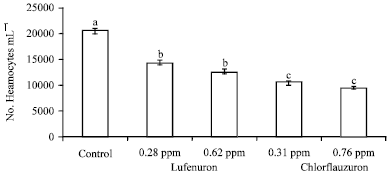 | |
| Fig. 1: | Efficacy of lufenuron and chlorfluazuron on the total haemocyte counts of S. littoralis 4th instar larvae. Error bars represent standard error of five replications. Columns within a group with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.01) |
| Table 4: | In vivo effect of lufenuron and chlorfluazuron on GST activity in 4th instar larvae of S. littoralis after different exposure times |
 | |
| Numbers within the same column with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.05) | |
The highest enzyme activity inhibition was occurred by lufenuron at 0.28 and 0.62 ppm, where the inhibition percentage was 38.6 and 45.6, respectively. These results are in agreement with Badawy et al. (2013), they reported a decrease in the GST activity of earthworms Aporrectodea caliginosa as a result of its exposure to lufenuron. In a contrast with these results, it has also been shown that certain insecticides induce GST activity (Clark, 1989).
Effect of the tested insecticides on Total Haemocyte Counts (THCs): The effects of lufenuron and chlorfluazuron in the haemolymph parameters after 72 h of treatments of treatments were evaluated. Figure 1 shows a fluctuation in mean number of haemocytes in the haemolymph of the 4th instar larvae of S. littoralis as a result of insecticide treatments after 72 h. The Total Haemocyte Counts (THC) were clearly affected by the tested insecticides. After treatment with lufenuron concentrations at 0.28 and 0.62 ppm THC count significantly decreased by 30.13 and 38.51%, respectively when compared with control but there are no significant different between two lufenuron concentrations. Moreover, treatment with chlorfluazuron at 0.31 and 0.76 ppm significantly decrease the THC by 49.26 and 53.89%, respectively, compared with control but also, no significant different in THC was observed between chlorfluazuron concentrations.
Six types of hemocytes were found in the hemolymph of S. littoralis and they were identified as Prohaemocytes, Granulocytes, Plasmatocytes, Oenocytoides and Spherule cells. With regard to lufenuron treatment, a reduction in the percentage of the Prohaemocyte was observed about 18.2 and 12%, at concentration 0.28 and 0.62 ppm, respectively but no significant different was observed between the tested concentrations of lufenuron.
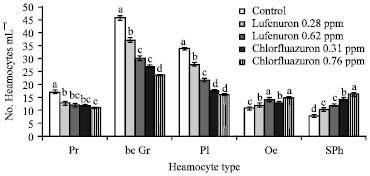 | |
| Fig. 2: | Efficacy of lufenuron and chlorfluazuron on the differential haemocyte counts of S. littoralis 4th instar larvae. Error bars represent standard error of five replications. Columns within a group with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.01) |
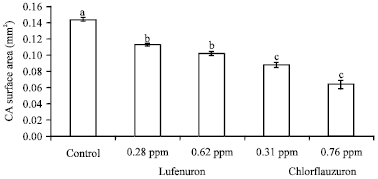 | |
| Fig. 3: | Efficacy of lufenuron and chlorfluazuron on the corpora allata activity of S. littoralis 4th instar larvae. Error bars represent standard error of five replications. Columns within a group with a letter in common are not significantly different according to analysis of variance (ANOVA) test (LSD at p<0.01) |
An obvious increase appeared in the number of the Granulocyte of the treated larvae (37.5 and 30.2%) compared with the total haemocyte population but still less than control (45.8%) (Fig. 2). The same trend was observed in the percentage of Plasmatocyte at 0.28 and 0.62 ppm were 28 and 21.6%, respectively.
The chlorfluazuron treatments was characterized its decrease in the number of plasmatocyte at 0.31 and 0.76 ppm treatments estimated by 17.5 and 16%, respectively, after 72 h of the treatment (Fig. 2). An increase in the activities of both oenocytoide and spherule cells characterized the treated larvae compared to the control. These results were similar to those obtained in previous studies (Arnold and Hinks, 1976) when they noted a high mitotic and rapid turnover of spherule cells, possibly as a mechanism of releasing products of their metabolism into the haemolymph. In addition, spherule cells were noted to play a role in recreation of some haemolymph proteins (Akai and Sato, 1973). Also, agreed to the results of Zibaee et al. (2011), suggested that treatment with pyriproxyfen reduces circulating hemocytes, including those involved in phagocytosis and encapsulation such as plasmatocytes and granulocytes and significantly impaired nodule formation as well as sluggish activity of PO system. Gad and Abdel-Megeed (2006), observed that spinosad and emamectin benzoate decreased the total and differential haemocyte counts in the S. littoralis.
Corpora allata activity: Results reported in Fig. 3 depict that treatment of S. littoralis 4th larval instar with both insecticides lufenuron and chlorfluazuron caused markedly decrease in the CA surface area after 72 h of treatment. Lufenuron at concentrations 0.28 and 0.62 ppm markedly decreased the CA surface area by 21.60 and 29.12%, respectively. Furthermore, chlorfluazuron at concentrations 0.31 and 0.76 ppm caused a sharp decrease in the CA surface area about 38.95 and 55.9%, respectively, compared to control. These results are in agreement with Gadot et al. (1991) suggests that 20E causes atrophy of CA cells linked with a low rate of JH synthesis in B. germanica. Also, Rankin and Stay (1985) reported that 20E provokes a reduction of JH titre in D. punctata. Furthermore, azadirachtin (AZA) causes a depletion of JH titres. Thus, the commonly reported decrease of JH titre by AZA could be imputed to the degeneration of CA cells (Malczewska et al., 1988). Also, Sayah (2002) reported that injection of AZA as well as 20-hydroxyecdysone (20E) into vitellogenic females of Labidura riparia caused ultrastructural damage in CA cells. However, some ultrastructural modifications are specific to each molecule.
CONCLUSION
Cotton leafworm is one of the most destructive agricultural lepidopterous pests. Chlorfluazuron and lufenuron (CSIs) are used for cotton leafworm control and they have many biochemical and physiological effects. While, activity level of ALT, AST and ALP increased, activity level of GST showed adverse results after exposure to lufenuron and chlorfluazuron. Lufenuron and chlorfluazuron reduced circulating hemocytes, including plasmatocytes and granulocytes and significantly inhabited the corpora allata activity after 72 h of lufenuron and chlorfluazuron treatment. The current study was performed and it was observed significant discrepancies in common physiological processes by changing the biochemical compositions in hemolymph and haemocytes population in Spodoptera littoralis larvae.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Abo Elghar, G.E., Z.A. Elbermawy, A.G. Yousef and H.K. Abd Elhady, 2005. Monitoring and characterization of insecticide resistance in the cotton leafworm, Spodoptera littoral is (Boisd.) (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol., 8: 397-410.
CrossRefDirect Link - Abou-Taleb, H.K., A.S.A. Saad, H.A. Mesbah, S.M. Abdel Rahman and D.A. El-Deeb, 2009. Toxicity of emamectin benzoate against laboratory and field strains of cotton leafworm with reference to its effects on the AST, ALT and ALP activity. Proceedings of the 6th International Symposium of Mediterranean Group on Pesticide Research, October 27-29, 2009, Cairo, Egypt.
- Arnold, J.W. and C.F. Hinks, 1976. Haemopoiesis in lepidoptera. I. The multiplication of circulating haemocytes. Can. J. Zool., 54: 1003-1012.
CrossRefDirect Link - Badawy, M.E.I., A. Kenawy and A.F. El-Aswad, 2013. Toxicity assessment of buprofezin, lufenuron and triflumuron to the earthworm Aporrectodea caliginosa. Int. J. Zool.
CrossRefDirect Link - Clark, A.G., 1989. The comparative enzymology of the glutathione S-transferases from non-vertebrate organisms. Comp. Biochem. Physiol. Part B: Comp. Biochem., 92: 419-446.
CrossRefDirect Link - DGKC., 1972. Empfehlungen der deutschen gesellschaft fur klinische chemie. Z. Klin. Chem. U. Klin. Biochem., 10: 182-192.
Direct Link - Eldefrawi, M.E., A. Toppozada, N. Mansour and M. Zeid, 1964. Toxicological studies on the Egyptian cotton leafworm, Prodenia litura. I. Susceptibility of different larval instars of Prodenia to insecticides. J. Econ. Entomol., 57: 591-593.
CrossRefDirect Link - Gadot, M., A.S. Chiang, E.L. Burns and C. Schal, 1991. Cyclic juvenile hormone biosynthesis in the cockroach, Blattella germanica: Effects of ovariectomy and corpus allatum denervation. Gen. Comp. Endocrinol., 82: 163-171.
CrossRefDirect Link - Hatem, A.E., H.B. Homam, R.A. Amer, S.A. Samad, H.A. Saleh and A.I. Hussien, 2009. Synergistic activity of several acids in binary mixtures with synthetic insecticides on Spodoptera littoralis (Boisduval). Bol. San. Veg. Plagas, 35: 533-542.
Direct Link - Jones, J.C., 1962. Current concepts concerning insect hemocytes. Am. Zool., 2: 209-246.
CrossRefDirect Link - Kai, Z.P., J. Huang, S.S. Tobe and X.L. Yang, 2009. A potential insect growth regulator: Synthesis and bioactivity of an allatostatin mimic. Peptides, 30: 1249-1253.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Malczewska, M., D.B. Gelman and B. Cymborowski, 1988. Effect of azadirachtin on development, juvenile hormone and ecdysteroid titres in chilled Galleria mellonella larvae. J. Insect Physiol., 34: 725-732.
CrossRefDirect Link - Peel, A.D. and M. Akam, 2007. The dynamics of yolk deposition in the desert locust Schistocerca gregaria. J. Insect Physiol., 53: 436-443.
CrossRefDirect Link - Ramaswamy, M., P. Thangavel and N.P. Selvam, 1999. Glutamic Oxaloacetic Transaminase (GOT) and Glutamic Pyruvic Transaminase (GPT) enzyme activities in different tissues of Sarotherodon mossambicus (Peters) exposed to a carbamate pesticide, carbaryl. Pestic. Sci., 55: 1217-1221.
CrossRefDirect Link - Rankin, S.M. and B. Stay, 1985. Regulation of juvenile hormone synthesis during pregnancy in the cockroach, Diploptera punctata. J. Insect Physiol., 31: 145-157.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Sayah, F., 2002. Ultrastructural changes in the corpus allatum after azadirachtin and 20-hydroxyecdysone treatment in adult females of Labidura riparia (Dermaptera). Tissue Cell, 34: 53-62.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Trowsdale, J., D. Martin, D. Bicknell and I. Campbell, 1990. Alkaline phosphatases. Biochem. Soc. Trans., 18: 178-180.
PubMed - Tunaz, H. and N. Uygun, 2004. Insect growth regulators for insect pest control. Turk. J. Agric. For., 28: 377-387.
Direct Link - Zibaee, A., I. Zibaee and J.J. Sendi, 2011. A juvenile hormone analog, pyriproxifen, affects some biochemical components in the hemolymph and fat bodies of Eurygaster integriceps Puton (Hemiptera: Scutelleridae). Pestic. Biochem. Physiol., 100: 289-298.
CrossRefDirect Link