
M. Shankara Murthy
Department of Entomology, College of Agriculture, Bheemarayanagudi, 585 287, India
V. T. Sannaveerappanavar
Department of Entomology, University of Agricultural Sciences, GKVK, Bangalore, 560 065, India
K. S. Shankarappa
Department of Horticultural Plant Pathology, K.R.C. College of Horticulture, Arabhavi, 591 218, University of Horticultural Sciences, Bagalkot, Karnataka, India
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 2 | Page No.: 95-101







ABSTRACT
Diamondback Moth (DBM), Plutella xylostella L. is one of the key insect pests of crucifer crops in India and elsewhere, causing huge crop losses. Despite its economic importance, population structure of this pest is not known from India. Therefore, genetic diversity of DBM collected from different cruciferous growing regions of Karnataka comprising the part of southern India (Bangalore, Belgaum, Hassan and Shimoga), Delhi and Ludhiana comprising northern parts of India was assessed using random amplified polymorphic DNA (RAPD) markers. Eighteen random primers were screened to investigate the genetic variability among the seven population of DBM. Five Operan (Op) primers viz., OpC 6, OpC 9, OpC 10, OpC 4 and OpB 20 out of eighteen produced unique banding patterns that could differentiate all the seven population and produce a total of 183 amplicons. Maximum number of amplicons obtained with primer OpC 6 and the least number of amplicons noticed with OpB 20 with the average amplicons of 36.60 per primer and all of them were polymorphic, indicating high genetic variability among DBM population. The cluster analysis of RAPD data clearly separated these populations into two distinct groups with first group consisting of only Delhi population and rest of the population falling into second group. The maximum dissimilarity (43%) was observed between the populations from Delhi and Belgaum. Whereas, the minimum dissimilarity (24%) was found in population from Ludhiana and Hassan. The present investigations have shown the existence of wide genetic diversity among the DBM populations within India.
PDF Abstract XML References Citation
Received: August 02, 2013;
Accepted: October 04, 2013;
Published: March 08, 2014
How to cite this article
M. Shankara Murthy, V. T. Sannaveerappanavar and K. S. Shankarappa, 2014. Genetic Diversity of Diamondback Moth, Plutella xylostella L. (Yponomeutidae:
Lepidoptera) Populations in India Using RAPD Markers. Journal of Entomology, 11: 95-101.
DOI: 10.3923/je.2014.95.101
URL: https://scialert.net/abstract/?doi=je.2014.95.101
DOI: 10.3923/je.2014.95.101
URL: https://scialert.net/abstract/?doi=je.2014.95.101
INTRODUCTION
The Diamondback Moth (DBM), Plutella xylostella (Linnaeus) (Yponomeutidae: Lepidoptera) is known for developing resistance to almost all insecticides used for its control (Sannaveerappanavar and Viraktamath, 2006). It causes significant economic damage to vegetable crops belonged to cruciferous family in tropical and sub-tropical countries grown throughout the year. The crop losses due to DBM may often go up to 100% (Calden and Hare, 1986). The world production of cruciferous crops is estimated to be around 42.2 million tonnes and the cost of control of DBM alone is about one billion US dollars per annum (Talekar and Shelton, 1993). In India, cabbage and cauliflower are grown in an area of 0.438 million hectares with a production of 6.34 million tonnes per annum. The annual loss due to the menace of DBM in India was estimated to be 16 million US dollars (Mohan and Gujar, 2003). Despite its economic importance, population structure of this pest is not known from India.
Different types of molecular approaches to characterize variability/diversity of species are available now. Development of molecular markers provides an excellent knowledge/information on the genetic diversity of DBM and helps to identify pesticide resistant populations in order to evolve appropriate management strategies (Roehrdanz and Degragillier, 2000). Apart from management aspects, these also provide the frame work to understand the taxonomy, population structure, races/biotypes etc. Among the different molecular tools, RAPD technique has been widely used in the determination of population structure of an insect, where prior knowledge of DNA sequences are not known and aids in fingerprinting of the genotypes and to develop DNA markers (RAPD markers) specific to races and biotypes (Harry et al., 1998). This technique has several advantages when compared to other techniques with respect to handling, simplicity and cost effectiveness. With this existing scenario, in the present study an attempt was made to study the genetic variability of P. xylostella population from six geographical regions representing the south and north Indian cruciferous ecosystem using RAPD markers.
MATERIALS AND METHODS
Insect populations: Larvae and pupae of DBM were collected from six locations of the south and north Indian cruciferous ecosystem viz., Bangalore, Hassan, Shimoga and Belgaum (Karnataka); Delhi and Ludhiana (Punjab) and a laboratory reared susceptible population. These were reared separately in the laboratory on mustard seedlings by adopting the mass rearing technique developed by Liu and Sun (1984) with suitable modifications to get the progeny.
Chemicals: All of the chemicals and reagents were of molecular grade. Primers, Taq polymerase, dNTPs, Magnesium chloride, 10x genie buffer, Agarose, Ethidium bromide were from Bangalore Genie Ltd, Bangalore, India.
DNA extraction: DNA isolations were performed as in the method described by Saghai-Maroof et al. (1984). In brief, 20 larvae of first generation progeny from each location were randomly picked, dissected and the gut contents were completely removed to avoid any contamination of plant DNA. Resulting skin and legs were used to prepare genomic DNA. These samples were stored at -20°C until further use. The larval sample was quickly frozen in liquid nitrogen and ground into a fine powder using mortar and pestle. The fine powder was transferred to a sterile tube containing 750 μL of lysis buffer (1% Cetyl Trimethyl Ammonium Bromide (C-TAB), 5% Polyvinyl Pyrrolidone (PVP), 1.4 M NaCl, 20 mM 2-mercaptoethanol) and mixed gently. To above mixture an equal volume (750 μL) of phenol: chloroform: isoamyl alcohol (25:24:1) was added and mixed well by inversion. The mixture was centrifuged at 8000 rpm for 10 min at 4°C and the upper aqueous layer containing the DNA was carefully transferred to another 1.5 mL centrifuge tube and equal volume of ice cold isopropanol was added. The mixture was gently shaken by inversion to precipitate the DNA. The precipitated DNA was again centrifuged at 8000 rpm for 5 min and the resultant supernatant/upper layer was discarded. The pellet was washed with 500 μL of 70% ethanol, dried and re-suspended in appropriate quantity of 10xTE buffer (10 mM Tris HCl (pH 8.0)+1mM EDTA).
| Table 1: | Primer sequences used for RAPD-PCR analysis in P. xylostella L. |
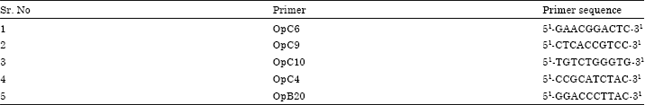 | |
The isolated DNA was purified and quantified according to the protocol described by Maniatis et al. (1982). The concentration of DNA was assessed by a spectrophotometer method using UV visible spectrophotometer. Finally, the DNA was stored at -20°C for further use.
PCR amplification: The genomic DNA from P. xylostella collected from six different places was subjected to Polymerase Chain Reaction (PCR) using five selected decamer random primers (Table 1). PCR was carried out for each primer in 25 μL standard reaction mixture consisting of 40 ng of template DNA, 0.2 mM primer, 3.5 mM magnesium chloride, 10x Bangalore Genie buffer, 3 U Taq polymerase and 10 mM each dATP, dTTP, dGTP and dCTP. PCR amplification was carried out using thermocycler (Eppendorf AG, Germany) in 0.2 mL micro-centrifuge tubes. Cycle conditions were 1 min initial denaturation at 94°C followed by 40 cycles of 1 min at 94°C for denaturation, 40 sec at 35°C for annealing, 1 min 30 sec at 72°C for extension and ended with 10 min final extension at 72°C.
Electrophoresis of PCR products: PCR products were separated on 1.0% agarose gels by using electrophoresis at 5 V cm-1 for 2.0 h. The banding pattern was visualized using ethidium bromide staining. All the 18 primers were tested at least twice for the reproducibility of banding pattern. A set of 5 primers producing reproducible banding patterns were selected for the present study. The molecular size of the amplified products was estimated using 1 kb DNA marker (MBI Fermentas). The gel pictures acquired through a gel documentation system into a computer were processed and scored to get binary data.
Scoring of bands and statistical analysis: PCR amplification products of the seven samples were scored as presence (1) or absence (0) of bands. Genetic similarity matrix was computed for each individual population using Jacquard similarity co-efficient (Sneath and Sokal, 1973). The resultant similarity matrix was used to generate/construct dendogram using unweighted pair group method with arithmetic average (UPGMA) statistical software.
RESULTS AND DISCUSSION
The genetic variability of P. xylostella population collected from different regions was investigated by RAPD-PCR analysis. Eighteen random primers were screened in the present study to investigate the genetic variability among the seven populations of DBM. The result revealed that, five out of the eighteen primers screened produced clear amplicons and were used for analysis. All the five primers viz., OpC 6, OpC 9, OpC 10, OpC 4 and OpB 20 resulted in a total of 183 amplicons. The total number of clear and scorable bands obtained from each primer ranged from 20 to 40, with an average of 36.60 bands per primer and all of them were polymorphic, indicating of high variability among P. xylostella populations (Table 2).
| Table 2: | Summary of statistics of random primers analysis for genetic diversity in P. xylostella |
 | |
| Table 3: | Dissimilarity coefficients of geographic populations of P. xylostella obtained by RAPD-PCR analysis |
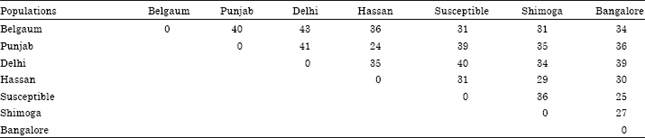 | |
Previously, Roux et al. (2007) reported similar results wherein a very high variability between populations of DBM with total polymorphism of 100%. Similarly, occurrence of a high level of genetic diversity among spatially isolated populations was observed in other insect pests like Helicoverpa in China and India (Zhou et al., 2000; Lei et al., 1995, 1996; Fakrudin et al., 2004) and Spodoptera litura F. (Janarthanan et al., 2002; Monnerat et al., 2006). Maximum number of bands (49) were obtained with primer OpC 6, followed by OpC 9 (46 bands), OpC 10 (38 bands) and OpC 4 (30 bands). The least number of bands (20) were noticed with OpB 20 primer. The size of the amplified fragments obtained varied from 375 bp to >2000bp. The representative gel showing the RAPD amplification patterns is shown in Fig. 1.
The dendrogram constructed using cluster analysis clearly showed that, existence of 2 principle clusters, with Delhi population alone is forming a single cluster and the rest of the populations forming the second cluster (Fig. 2). The second cluster was subdivided into two sub-clusters with Punjab and Hassan populations forming a single cluster while the populations of Bangalore, Shimoga and Belgaum along with the susceptible laboratory population formed second sub-cluster. The second sub-cluster was further divided into two smaller clusters with Bangalore and susceptible cultures forming one cluster and the populations of Shimoga and Belgaum forming another. Further, matrix of genetic distances among the seven DBM populations indicated that Delhi and Belgaum populations with 43% genetic distance were the most diverse populations (Table 3). While, Punjab and Hassan with 24% dissimilarities were the closest populations, followed by susceptible and Bangalore with 25%. The population collected from different locations formed into groups according to their geographical origin except Hassan population indicating RAPD polymorphism partly corresponded to the distance or clustering of the DBM population was not in line with the geographical demarcation. This could be attributed to biological characteristics of DBM. Secondly, independent evolution of these populations generated genetic divergence. Roux et al. (2007) who reported genetic differentiation among the populations did not reflect geographical distances between them. A similar result was also reported by Endersby et al. (2006) found no differentiation with microsatellites.
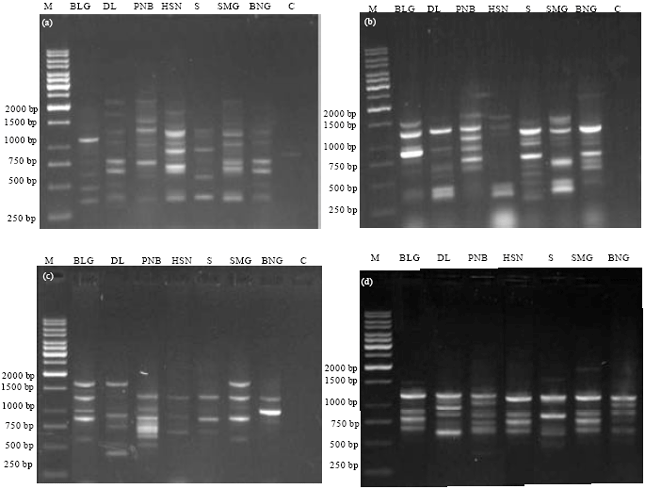 | |
| Fig. 1(a-d): | Agarose gel showing RAPD banding patterns of diamond back moth from different geographical regions of India using operan primer (a) OpC-9, (b) OpC-6, (c) OpC-4 and OpC-10. Lanes M = 1 kb molecular weight marker (MBI Fermentas Life Sciences, Germany) BLG = Belgaum, DL = New Delhi, PNB = Punjab HSN = Hassan, S = Susceptible SMG = Shimoga, BNG = Bangalore, C = Water control |
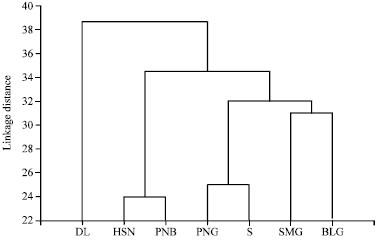 | |
| Fig. 2: | Dendogram showing genetic diversity of P. xylostella populations based on RAPD markers |
CONCLUSION
The present investigations have shown the existence of wide genetic diversity among the DBM populations within India. Detailed studies are needed to identify genetic markers linked to insecticide resistance mechanisms in these populations. This may intern help in evolving appropriate management strategies.
ACKNOWLEDGMENT
We are thankful to Dr. K.T. Rangaswamy, Professor, Department of Plant Pathology and Dr. Shailaja Hitalamani, Professor, Department of Genetics and Plant breeding, University of Agricultural Sciences, Bangalore for providing the laboratory facilities for carrying out the study.
REFERENCES
- Endersby, N.M., S.W. McKechnie, P.M. Ridland and A.R. Weeks, 2006. Microsatellites reveal a lack of structure in Australian populations of the diamondback moth, Plutella xylostella (L.). Mol. Ecol., 15: 107-118.
CrossRef - Fakrudin, B., S.H. Prakash, K.B. Krishnareddy, Vijaykumar, P.R. Badariprasad, B.V. Patil and M.S. Kuruvinashetti, 2004. Genetic variation of cotton bollworm, Helicoverpa armigera (Hubner) of South Indian cotton ecosystem using RAPD markers. Curr. Sci., 87: 1654-1657.
Direct Link - Janarthanan, S., S. Seshadri, K. Kathiravan and S. Ignacimuthu, 2002. Use of RAPD in assessing the genetic variability in Spodoptera litura. J. Exp. Biol., 40: 839-841.
Direct Link - Lei, Z., L. Li, Y. Guo, W. Wang and K. Yu, 1995. RAPD analysis of two sibling species of Helicoverpa hardwick from China. Plant Prot., 21: 10-12.
Direct Link - Liu, M.Y. and C.N. Sun, 1984. Rearing diamondback moth (Lepidoptera: Yponomeutidae) on rape seedlings by a modification of the Koshihara and Yamada method. J. Econ. Entomol., 77: 1608-1609.
Direct Link - Monnerat, R., E. Martins, P. Queiroz, S. Orduz and G. Jaramillo et al., 2006. Genetic variability of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis cry toxins. Appl. Environ. Microbiol., 72: 7029-7035.
CrossRef - Mohan, M. and G.T. Gujar, 2003. Local variation in susceptibility of the diamondback moth, Plutella xylostella (Linnaeus) to insecticides and role of detoxification enzymes. Crop Prot., 22: 495-504.
CrossRef - Roux, O., M. Gevrey, L. Arvanitakis, C. Gers, D. Bordat and L. Legal, 2007. ISSR-PCR: Tool for discrimination and genetic structure analysis of Plutella xylostella populations native to different geographical areas. Mol. Phylogenet. Evol., 43: 240-250.
CrossRefDirect Link - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Talekar, N.S. and A.M. Shelton, 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol., 38: 275-301.
CrossRefDirect Link - Zhou, X., S.W. Applebaum and M. Coll, 2000. Overwintering and spring migration in the bollworm Helicoverpa armigera (Lepidoptera: Noctuidae) in Israel. Environ Entomol., 29: 1289-1294.
CrossRefDirect Link