
J.A. Ugwu
Department of Agricultural Technology, Federal College of Forestry Ibadan, Forestry Research Institute of Nigeria, Nigeria
A.A. Omoloye
Department of Crop Protection and Environmental Biology, University of Ibadan, Nigeria
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 1 | Page No.: 34-41







ABSTRACT
Iroko gall bug, Phytolyma lata (Homoptera; Psylidae) is a major insect pests militating against cultivation of Milicia exceslsa (Iroko) in many parts of West Africa. The reproductive and developmental biology was studied to give an insight on the management approach that could be adopted for it. The study was conducted on 6 month-old iroko seedlings in the laboratory following standard procedure at 27-32±0.3°C and 75-80% relative humidity and 12:12 photoperiod. Developmental period, sex ratio, fecundity, oviposition period and adult longevity were assessed. Adult females Phytolyma lata were oviparous under laboratory condition with; pre-oviposition period (1.40±0.54 days), oviposition periods (4.00±1.22 days), fecundity (41.20±5.35/female), incubation period (5.8±0.54 days) and 95% egg viability. There were five nymph instar stages. The body width and head width of female were significantly larger than male (p>0.05).The average growth rate for the five instars was 1.48. The average life cycle of Phytolyma lata was 17.55±0.97 days and the sex ratio was 1:1.45 in favour of females. There was significant (p<0.05) correlation (r = 0.973) between the stages of nymphal development and the head width. Mated and unmated male lived significantly (p<0.05) shorter than mated female unmated female under laboratory condition.
PDF Abstract XML References Citation
Received: June 24, 2013;
Accepted: November 18, 2013;
Published: March 08, 2014
How to cite this article
J.A. Ugwu and A.A. Omoloye, 2014. Biology of Iroko Gall Bug, Phytolyma lata Scott. (Homoptera; Psylidae)
on Milicia excelsa (Welw) C.C. Berg. Journal of Entomology, 11: 34-41.
DOI: 10.3923/je.2014.34.41
URL: https://scialert.net/abstract/?doi=je.2014.34.41
DOI: 10.3923/je.2014.34.41
URL: https://scialert.net/abstract/?doi=je.2014.34.41
INTRODUCTION
Milicia excelsa (Welw) C. C. Berg commonly known as Iroko is an important timber trees species in Africa. It belongs to the family Moraceae and distributed across the entire breadth of Africa (Keay, 1989). M. excelsa wood is the most valuable timber species from all of West, Central and East Africa. The wood is extensively used due to its high durability and good working properties. There is high demand of M. excelsa timber and other products from Milicia wood such as sliced veneer, rotary veneer and profile boards for decorative and structural uses. Cultivation of Milicia species is being constrained widely across West Africa by Iroko gall bug, Phytolyma lata (Cobbinah and Wagner, 1992; Cobbinah and Appiah-Kwarteng, 1996). The psyllid (iroko gall bug) attack the buds and young leaves of Milicia excelsa plants especially the seedling which later leads to formation of galls on the site of attack. The galls afterward burst to release adults when they psyllid completes their life cycle inside the gall. The bug lays eggs on young stems, leaves and shoots of the host tree in high numbers (Nichols et al., 1998). When the first instar nymphs hatches, it punctures plant surfaces and induces gall formation which later develops to enclose the nymph inside the gall.
Generally, gall formation is followed by saprophytic fungi attack on the apical region of the infested plant after the gall has ruptured to release adult leading to foliage die-back down to the wood tissue (Cobbinah and Wagner, 1995). However, the attacks decrease with age, as the old trees tend to be more resistant. Phytolyma attack stunts its growth and affects stem shape and size and also reduces the regeneration rate of the species.
Over the years, attention of researchers has been on natural regeneration (Nichols et al., 1998) and evaluation of progeny for resistant lines of M. excelsa (Cobbinah and Wagner, 1995) but a great deal of success has not been recorded. Also several chemicals have been evaluated in both Ghana and Nigeria for control of Phytolyma species but prove ineffective due to the cryptic nature of the pest. The knowledge of the biology of a pest gives an insight on the management approach that could be adopted for its Integrated Pests Management (IPM). However, currently there is dearth information on the biology of Phytolyma lata in Nigeria. Therefore this work reports the reproductive and developmental biology of Phytolyma lata on Milicia excelsa under laboratory condition.
MATERIALS AND METHODS
Experimental site: The experiment was conducted at the Entomology Laboratory of Department of Crop Protection and Environmental Biology, University of Ibadan under ambient temperature, 24-27°C, 88-92% relative humidity and 12:12 day photoperiod. Treatments were arranged Completely Randomized Design (CRD) and all measurement were made under microscope fitted with an ocular micrometer eye-piece (x10).
Insect culture: Six month old Milicia seedlings were placed inside wooden cage (170 cm heightx30 cm diameter) and a matured gall collected from the wild were introduced inside the cage. The galls were allowed to release adult P. lata inside the cage and the seedlings were infested. The plants were watered daily and monitored for gall formation. The P. lata culture was maintained in the cage until the end of the study (Fig. 1).
Life cycle study
Egg: Teneral adults were picked with the aid of a camel hair brush and paired in oviposition cage (13 cm heightx6 cm diameter). The sexes of the insects were determined by comparing their external genitalia with the aid of hand lens. Fresh Milicia leaves and stem were introduced into separate cages daily and sugar solution (25%) soaked in cotton wool was introduced to supplement their food. The experiment was replicated four times. The paired adult P. lata was monitored for oviposition, the female laid on the fresh leaves and stem which was replaced daily until the female stopped ovipositing. Eggs on the leaves and stem were removed, counted and placed in another cage for incubation. The incubation cage was kept moist to prevent desiccation. The pre-oviposition, oviposition period, fecundity, incubation period and egg viability were recorded. The egg viability was determined by counting the number of first instar nymph that hatched out from the incubated eggs after two days. A sample of 20 eggs was collected with the aid of camel hair brush and measured for egg length and width.
Nymphal stages: Twenty (20) first instar nymph were transferred with aid of camel hair brush from the incubation cage to the wooden cage (170 cm heightx30 cm width) which enclosed a six-month Milicia seedling and were replicated four times.
 | |
| Fig. 1: | Culturing of Phytolyma lata under laboratory condition |
The nymphs were allowed to feed on the young leaf tissues of the shoot apex and thereafter monitored for gall formation. At the initiation of gall formation, starting from the second day of gall formation, each gall was split open every day to search for exuviae, which clearly indicated molting (Ewete and Olagbaju, 1990). Twenty samples of each nyphal stage were collected and preserved in 70% alcohol for measurement of the body length, body width, head width, the developmental period of each nymphal stage and records of characteristics of each nymphal stages. The observed head width was tested for conformity with Dyar,s rule (Dyar, 1890) by comparing the observed and calculated average head width. The calculated average head width of an instar was obtained as a product of the mean head width of the succeeding instar and its mean growth ratio. The growth ratio was calculated as the quotient of the observed mean head width of the succeeding instar and the observed mean head width of the previous instar.
Adult: Thirty male and female newly emerged adults were collected and preserved in 70% alcohol for the measurement of body length, body width and head width. The sex ratio was determined by introducing a pair of teneral adult to a six-month’s old M. excelsa seedling inside a wooden cage (170 cm heightx30 cm diameter) where they fed, mated and oviposited on the potted plant. Galls formed were monitored until maturity and adult were released under natural condition. The emerged adults were counted and the sex ratio was determined per each cycle. The longevity of mated and unmated adults was also determined.
Data analysis: Data on the morphometric of the nymphal and adult stages were subjected to Analyses of Variance and significant means were separated using Duncan Multiple Range Test (DMRT). The relationship between the head width and developmental period were subjected to Correlation analysis.
RESULTS
Phytolyma lata have seven life stages: Egg, five instars and the adult. The female gall bug oviposited 24 h after emergence and oviposition lasted 3-6 days with an average of 4.00 days. The average number of eggs laid per female was 41.2±5.35 with range from 32-45 eggs (Table 1). The eggs were laid either in rows or sometimes scattered singly on the leaves or stem of M. excelsa. About 35-66% of the eggs were laid on the first day and within 2-3 days over 80% of the total number of eggs had been laid. The pre-oviposition period of mated female P. lata under laboratory condition on M. excelsa leaves was 1.4±0.54 and 2.00±0.50 on the stem with a range of 1-2 days (Table 1). Eggs are oval in shape, creamy white when just laid and anteriorly pointed (Fig. 2a). The egg was 0.4±0.50 mm in length and 0.2±0.50 mm in width (Table 2). The average incubation period of the eggs was 5.80 days. The newly emerged first instar nymph is cream colour, head quadrangle with a pair of compound eyes, two antennae, the thorax and abdomen are not segmented (Fig. 2b). The first instar nymph was 3.2±0.92 mm in length and 1.90±0.74 mm in width (Table 2). Second instar nymph is creamy white in colour, with two prominent antennae, the two eye spot were dark brown in colour (Fig. 2c). The mean body length was 9.50±0.50 mm, width 7.20±1.77 mm and the head width 4.45±0.60 (Table 2). Third instar is creamy in colour; eye spot dark brown, the thorax and the abdomen were clearly defined, segmented and with wing pad (Fig. 2d). The mean body length was 16.63±3.55 mm, body width 10.64±1.36 mm and head width was 6.59±0.91 mm (Table 2). Fourth instar nymph are greenish; the abdominal segment distinct and clearly demarcated from the thorax, eye spot dark and reduced (Fig. 2e). The mean body length was 28.0±3.21 mm, mean body width 12.93±1.67 mm and head width 7.50±0.76 mm (Table 2). The fifth instar is greenish, eye spot dark and reduced, wings are developed though very tender and transparent, the abdominal segment and the genital organ appeared distinct. The general body feature presented an appearance of miniature adult (Fig. 2f). The mean body length was 30.00±0.67 mm, body width 12.20±1.40 mm and head width 7.40±0.70 mm (Table 2). Adult is dark brown in colour with fully developed pair of wing adapted for short flight.
| Table 1: | Pre-oviposition, oviposition periods, fecundity and incubation periods of P. lata on M. excelsa |
 | |
| Table 2: | Mean developmental periods and body morphometrics of developmental stages of Phytolyma lata on Milicia excelsa |
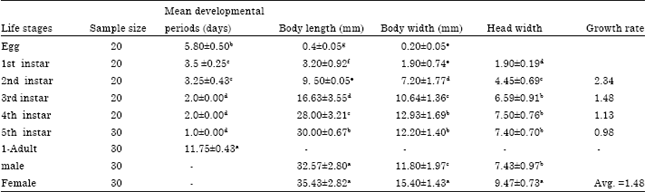 | |
| Growth rate: The mean head width of a preceding instar divided by the mean head width of a succeeding instar | |
 | |
| Fig. 2(a-h): | Life stages of Phytolyma lata, (a) Eggs x100, (b) 1st instar x100, (c) 2nd instar x100, (d) 3rd instar x50, (e) 4th instar x50, (f) 5th instar x25, (g) Adult male x25 and (h) Adult female x25 |
| Table 3: | Head width for nymphal instars and t-test for conformity to Dyar’s rule |
 | |
| Growth ratio: Observed mean head width of nymphal instar divided by observed mean head of succeeding nymphal instar, Calculated mean = observed mean head capsule (mm) of a succeeding instar multiplied by the mean of the growth ratio, Mean of the growth ratio = 0.75 | |
There are distinct light yellow markings that separate each abdominal segment, the colour of the markings of the female are more distinct and deeper in colour than that of the male (Fig. 2g and h). The female has ovipositor at the last segment which has dark spots at the tip while the male has aedeagus which when pulled out during sex are bifurcated. The mean body length of the male was 32.57±2.80 mm, female 35.43±2.82 mm; body width of the male was 11.80±1.97, female 15. 40±1.43; the head width of the male was 7.43±0.97 mm, female 9.47±0.73 mm (Table 2). The mean duration of developmental period of the nymphal instars were: 1st instar 3.50±0.25 days, 2nd instar 3.25±0.43 days, 3rd instar 2.00±0.00 days, 4th instar 2.00±0.00 days and 5th instar 1.00±0.00day. The total developmental period of the nymphal instars was 11.75 days (Table 2).
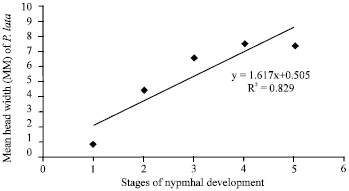 | |
| Fig. 3: | Relationship between the nymphal stages of P. lata and their head width |
| Table 4: | Mean longevity (days) of adult P. lata fed with sugar solution under laboratory condition |
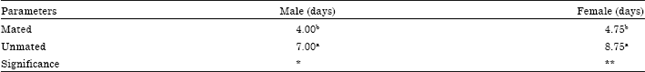 | |
| Means followed by same letters in a column are not significantly different at 5% level using duncan multiple range test *: 5% level of significance; **: 1% level of significance | |
There was significant (p<0.05) correlation (r = 0.973) between the stages of nymphal development and the head width (Fig. 3). Ninety-five percent of eggs laid hatched under ambient temperature 24-27°C, 88-92% relative humidity and 12:12 h day photoperiod.
The observed values of head width of various larval instars when compared with calculated values showed conformity to Dyar’s rule (Table 3). The mean longevity of mated adult males was 4.00 days, unmated 7.00 days while for mated female it was 4-75 days, unmated 8.75. Mated male lived significantly (p<0.05) shorter than the mated female under laboratory condition. Moreover, unmated male lived significantly (p<0.01) shorter than unmated female (Table 4).
DISCUSSION
The female Phytolyma lata were oviparous under laboratory condition. Eggs were laid in batches of 3-15 on the leaves and stem of young Milicia seedlings, a higher batches of eggs were reported by Wagner et al. (1991).
The mean incubation period of P. lata eggs in this study under laboratory condition was 5.8 days ranged (4-8 days). This was in line with the earlier report of Wagner et al. (1991) that the incubation period of P. lata is about 8 days. White (1966), Browne (1968) and Cobbinah (1986) also reported approximately 8 days of incubation. However, when eggs were kept in the cages and covered with leaves, the incubation period was shorter. This indicates that temperature is a major factor in the incubation of the egg. It also suggests that the incubation period of P. lata may vary from one location to another depending on the prevailing environmental conditions. The little variation in the incubation period of P. lata in this study and that reported by Wagner et al. (1991) could be attributed to the variation in the environmental conditions of the different study areas. Ninety five percent of egg viability observed in this study indicates the successful reproductive ability of the insects. This also attributes to the high population of the insects that account for the constant infestation on M. excelsa throughout the year. The viability of the eggs enhances their chances of multiple generations that were earlier reported. Cobbinah (1986) reported that Phytolyma is multivoltine and ten or more generations can be recorded in a year. It was observed that P. lata completes it life cycle on M. excelsa between 24-26 days (3-4) weeks. This was in the same range with the earlier report by Cobbinah (1986), that it takes 2-3 weeks for P. lata to completes it life cycle. Female P. lata are moderately fecund, with an average of 41.2 eggs being laid by a female under laboratory condition. In contrast, Wagner et al. (1991) reported that approximately 9-546 eggs are laid by female. The environmental factors could be responsible for the variation in the fecundity of the female P. lata. Higher number of eggs were laid by the female at the beginning of the rainy season (March-June) and the sizes of adult P. lata were also bigger. The variations are probably due to variation in the environmental conditions. This suggested that environment was more favourable for the insect during the rainy season. There were five nymphal instars as was previous reported by Cobbinah (1986). The ratio of the previous instar’s nymphal head width to the succeeding one obeyed Dyar’s rule (Dyar, 1890). The head size increased at each molt by an average of 1.48 approximating the expected constant ratio of 1.4 for lepidopterous (Wigglesworth, 1974). A linear relationship was obtained between the head width and with a high correlation coefficient of 0.91. This indicates that variation in the developmental period for each nymphal instar was not strong enough to vacate Dyar’s rule.
CONCLUSION
The problem of P. lata on Milicia excelsa can be curtailed with identification of appropiate Integrated Pest Management (IPM). The proper understanding of the biology of a pests provides the pedestal for a more sustainable control method. Therefore, this work has provided a base for the researcher to design a sustainable Integrated Pest Management(IPM) for Phytolyma lata.
ACKNOWLEDGMENTS
This study was part of the first author’s postgraduate work in the Department of Crop Protection and Environmental Biology, University of Ibadan. We are grateful to ETF for sponsoring part of this study and to the Forestry Research Institute of Nigeria for granting the enabling environment towards the success of this study.
REFERENCES
- Cobbinah, J.R., 1986. Factors affecting the distribution and abundance of Phytolyma lata (Homoptera: Psyllidae). Int. J. Insect Sci., 7: 111-115.
CrossRef - Cobbinah, J.R. and M.R. Wagner, 1995. Phenotypic variation in Milicia excelsa to attack by Phytolyma lata (Psyllidae). For. Ecol. Manage., 75: 147-153.
CrossRefDirect Link - Dyar, H.G., 1890. The number of molts of lepidopterous larvae. Psyche, 5: 420-422.
CrossRefDirect Link - Ewete, F.K. and R.A. Olagbaju, 1990. The development of Aspavia armigera Fabricius (Hemiptera: Pentatomidae) and its status as a pest of cowpea and rice. Int. J. Trop. Insect Sci., 11: 171-177.
CrossRef - Nichols, J.D., M.R. Wagner, V.K. Agyeman, P. Bosu and J.R. Cobbinah, 1998. Influence of artificial gaps in tropical forest on survival, growth and Phytolyma lata attack on Milicia excelsa. For. Ecol. Manage., 110: 353-362.
CrossRefDirect Link