
Elisabeth Noubissi Youmbi
Department of Biological Sciences, Laboratory of Zoology, Higher Teachers Training College, University of Yaounde I, P.O. Box 47, Yaounde, Cameroon
Wenceslas Yana
Faculty of Science, University of Bamenda, P.O. Box 39, Bamenda, Cameroon
Joseph Lebel Tamesse
Department of Biological Sciences, Laboratory of Zoology, Higher Teachers Training College, University of Yaounde I, P.O. Box 47, Yaounde, Cameroon
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 4 | Page No.: 182-197







ABSTRACT
Triplochiton scleroxylon is a plant species of tropical Africa. It’s known as "Ayous" in Cameroon. It’s soft, light wood and white cream; it can be used industrially and has an economic importance. In Cameroon, the national reforestation programs of T. scleroxylon stumble on the proliferation of psyllids pest of this plant. Two psyllids feed on T. scleroxylon in Cameroon, Diclidophlebia eastopi and D. harrisoni. D. eastopi larvae caused leaf discoloration and stunting of seedlings. D. harrisoni larvae caused leaf curl. Damages are important on young plants. The population dynamics of these two psyllids species was conducted in a natural secondary forest from February 2009 to January 2011. We collected 570 males and 597 females of D. eastopi and 295 males and 269 females of D. harrisoni. The numerical variation of psyllids population in 2009 shown six main generations for D. eastopi and five main generations for D. harrisoni. In 2010, six generations were obtained for D. eastopi against seven generations for D. harrisoni. Some climatic factors influenced the numerical variation of psyllids on T. scleroxylon in Cameroon. The phenology of the host plant seems to be the main biotic factor influencing the numerical variations of psyllids on T. scleroxylon. A better understanding of the population dynamic of psyllids pest of commercial timbers could be very important for an integrated pest management in the tropical forest of Africa.
PDF Abstract XML References Citation
Received: November 15, 2013;
Accepted: February 18, 2014;
Published: April 18, 2014
How to cite this article
Elisabeth Noubissi Youmbi, Wenceslas Yana and Joseph Lebel Tamesse, 2014. Population Dynamic of Diclidophlebia eastopi Vondracek and Diclidophlebia
harrisoni Osisanya, Psyllids Pest of Triplochiton scleroxylon (K.
Schum) (Urticales: Sterculiaceae) in Yaounde, Cameroon. Journal of Entomology, 11: 182-197.
DOI: 10.3923/je.2014.182.197
URL: https://scialert.net/abstract/?doi=je.2014.182.197
DOI: 10.3923/je.2014.182.197
URL: https://scialert.net/abstract/?doi=je.2014.182.197
INTRODUCTION
Triplochiton scleroxylon is a large deciduous forest tree commonly attaining 45 m in height and 1.5 m in diameter. Triplochiton scleroxylon is known under various and common names according to each country of origin: "Ayous" or "Ayos" from Cameroon, "Wawa" from Ghana, "Samba" from Cost Ivory, "Obéché" or "Arere" from Nigeria, "Babo" from Central Africa Republic; this plant is also knew as "Maple African" or "African white wood" (Halle, 1961; Marche-Marchand, 1965; Vivien and Faure, 1985). The tree is found in the semi-deciduous moist forest, in the transition zone between forest and moist savannah and in scattered outliers where local topography favors a closed forest community. Trees occur naturally from Guinea to Democratic Republic of Congo and from Gabon to Nigeria. T. scleroxylon is referred to as a pioneer species and it has been suggested that shifting cultivation in West Africa has influenced the natural distribution. Trees normally occur in clusters of 10 or more and isolated trees are very rare. Obeche is recognized as a very important source of timber for export. It is one of the three timber species (others are Entandrophragma cylindricum and Lophira alata) that have traditionally accounted for more than half of timber supply in Cameroon (Wunder, 2003). These different uses of T. scleroxylon give an undeniable economic importance (Halle, 1961; Vivien and Faure, 1985). Among the pests of this plant are psyllids of Diclidophlebia genus.
Psyllids are sap-sucking insects of the plants. They are very specific on their host plants and can be monophagous or oligophagous. During the nutrition, psyllids fit their mandibular and maxillary stylets in the plant tissues and they suck the sap. The saliva injected could contain bacterial like germs (Aubert et al., 1988; Marutani-Hert et al., 2011) or virus (Marutani-Hert et al., 2009). According to Li (2011), about 3850 psyllids species are described worldwide. They belong to the order of Hemiptera and are mainly associated with perennial plants (Hodkinson, 1974; Burckhardt, 1994; Hollis, 2004). However, some species are associated with monocotyledons (Burckhardt, 1987) and other species are recorded on conifers (Burckhardt, 2005). Psyllids can become serious pests that hinder the growth and development of crops (Burckhardt, 1994; Tamesse et al., 1999; Brennan et al., 2001; Burckhardt et al., 2006; Dzokou et al., 2009; Yana et al., 2009). Trioza erytreae Del Guercio and Diaphorina citri Kuwayama are the vectors of huanglongbing or “greening” which is a citrus disease in Africa in general and Cameroon in particular (Wagner et al., 1991) for the first citrus psyllid species and in Asia and America for the second citrus psyllid species (Bonani et al., 2010; Leon et al., 2011; Tiwari et al., 2011). In Cameroon, the following authors, Burckhardt (1987) and Burckhardt et al. (2006) studied the biodiversity of Psyllids and they included the genus Diclidophlebia in Paurocephalinae subfamily. But the recent taxonomic revision of Psyllidae by Burckhardt and Ouvrard (2012) included the same genus in the Psyllinae subfamily. Burckhardt and Mifsud (2003) described 25 species in Diclidophlebia genus with 13 neotropical species, 9 indoaustralian species and 3 afrotropical species including D. eastopi and D. harrisoni. These two species feed on T. scleroxylon in Cameroon. In Nigeria, under the conditions of nursery seedling plants growth is slowed sharply and many seedlings die due to repeated attacks by psyllids (Osisanya, 1974a). The control of the insect pests is based on a thorough understanding of their population dynamics (Riba and Silvy, 1989). According to Milaire (1987), studies on population dynamics help to keep pest populations below the threshold of economic damage by insecticides. It allows finding strategies to optimize the use of chemicals in integrated program of control (Webb, 1977) and provides useful information for taking preventive measures against the resurgence of infestations (Nyeko et al., 2002).
The aim of this study was to study the population dynamics of D. eastopi and D. harrisoni, which feed on T. scleroxylon, in Cameroon. Abiotic and biotic factors that regulated natural populations of the pest are investigated.
MATERIALS AND METHODS
Site and period of study: The study was conducted for two consecutive years from February 2009 to January 2011 in a natural secondary forest at Nkolbisson, VII th district of Yaounde. The altitude and geographical coordinates are: 740 m, 03°52'290'' N 11°25'420'' E. Host plants were planted three years ago and they are about 2 m height. The total number of plants is twenty. Ten plants were chosen randomly for psyllids collection. These plants received no pesticide treatment before and during the study period.
Sampling method: Sampling was done once a week from February 2009 to January 2011 (24 months). Adult psyllids were captured with a sweep net of 0.5 mm mesh size and with an aspirator. Larvae were collected directly from buds and leaves of the host plant. All specimens were preserved in 70% ethanol and kept in the laboratory and were examined under a stereomicroscope. In the laboratory, adults were counted and grouped to male and female. Larval stages are identified and their number counted.
Meteorological data were obtained from the Institute of Agricultural Research for Development (IRAD), Nkolbisson. The selected parameters are temperature, relative humidity, rainfall, wind speed and insolation.
Data analysis: SPSS statistical program was used to compare mean with nonparametric Wilcoxon and Mann-Whitney tests (p<0.05). This program was also used to calculate the Spearman correlations between abiotic and biotic factors regulation population dynamic of Diclidophlebia spp. psyllids of T. scleroxylon.
RESULTS
Numerical variations of Diclidophlebia eastopi population: The numbers of psyllids counted on T. scleroxylon varied constantly from a week to another.In 2009, the numerical variations showed five main peaks for the first larval stage respectively in March, May, July, September and January and in 2010, the six peaks were obtained in March, May, July, September, November and January 2011 (Fig. 1). In 2009, the numerical variations for the second larval stage showed six peaks in February, May, July, September, November and January and in 2010, the peaks were obtained in March, May, July, September, November and January 2011 (Fig. 1). For the third larval stage, the numerical variations, in 2009, showed six main peaks in March, May, July, October, November and January and in 2010, the five peaks were obtained in May, July, September, November and January (Fig. 1). The numerical variation of the fourth larval stage in 2009 showed six main peaks in February, April, July, September, November and December and in 2010, the five peaks were observed in February, May, July, October and December (Fig. 2). Lastly for the larval stages, the numerical variations of fifth instars larvae showed in 2009 six peaks in February, May, July, October, November and January and in 2010, five main peaks were obtained in February, April, June, October and January (Fig. 2).
The numerical variations of adults showed in 2009 and 2010 six main peaks. But those peaks were not in the same month during the 2 years (Fig. 3). The number of generations of D. eastopi in our region was 6 in 2009 and 2010. In total, 1167 adults were collected including 570 males and 597 females. The sex ratio was 0.96 in favor of female. Concerning the larvae stages, we obtained 1972 larvae of first stage, 1361 of second larvae stage, 972 of third larval stage, 713 of fourth larval stage and 540 larvae of fifth stage.
The statistical analysis showed that they was no significant difference between the average numbers of psyllids collected in 2009 and 2010 (Z = -3.45; P = 0.73 for the first larval stage; Z = -4.07; P = 0.68 for the second larval stage; Z = -4.60; P = 0.64 for the third larval stage; Z = -6.82; P = 0.49 for the fourth larval stage; Z = -8.56; P = 0.39 for the fifth larval stage; Z = -1.44; P = 0.14 for the males and Z = -1.89; P = 0.05 for the females). D. eastopi population seems to be the same from 1 year to another in our region.
Numerical variations of Diclidophlebia harrisoni population: The number of psyllids counted on T. scleroxylon varied constantly from one week to another.
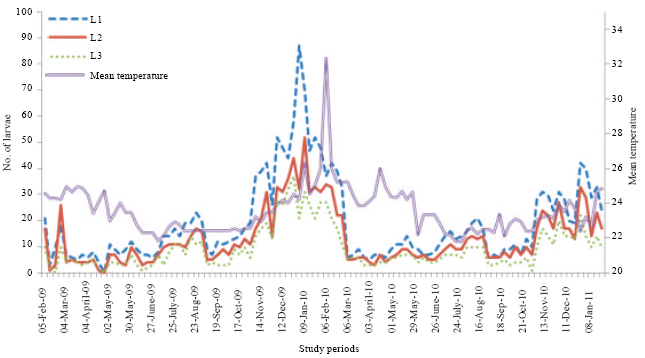 | |
| Fig. 1: | Numerical variations of larvae of early stages of D. eastopi on T. scleroxylon in an experimental plantation depending on the temperature variation in Yaounde from February 2009 to January 2011 |
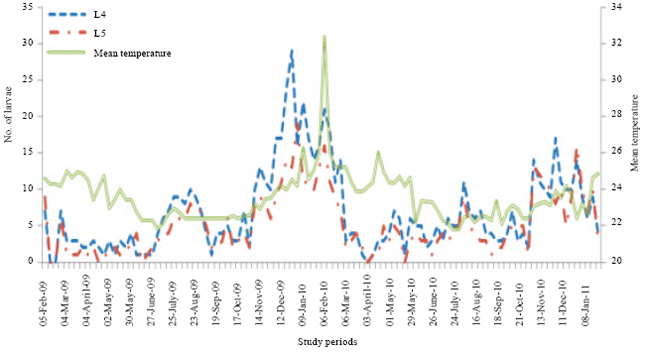 | |
| Fig. 2: | Numerical variations of larvae of advanced stages of D. eastopi on T. scleroxylon in an experimental plantation depending on the temperature variation in Yaounde from February 2009 to January 2011 |
In 2009, the numerical variations of the first larval stage showed seven peaks obtained in February, March, July, September, November, December and January; in 2010, eight peaks were obtained, respectively in February, March, May, July, September, November, December and January (Fig. 4).
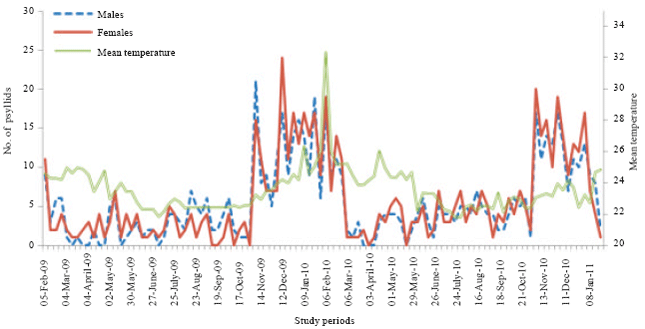 | |
| Fig. 3: | Numerical variations of D. eastopi adults depending on the temperature variation in an experimental plantation in Yaounde from February 2009 to January 2011 |
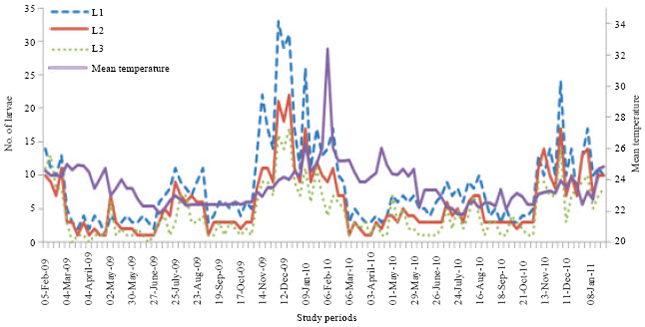 | |
| Fig. 4: | Numerical variations of larvae of early stages of D. harrisoni on T. scleroxylon in an experimental plantation depending on the temperature variation in Yaounde from February 2009 to January 2011 |
Concerning the second larval stage, in 2009, the numerical variations showed six peaks in March, May, July, September, November and January and in 2010, the peaks were obtained in February,March, July, November and January 2011 (Fig. 4).
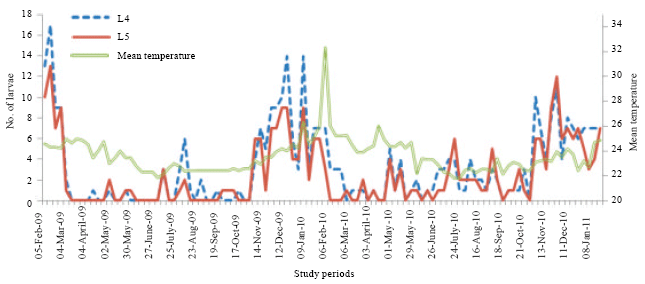 | |
| Fig. 5: | Numerical variations of larvae of advanced stages of D. harrisoni on T. scleroxylon in an experimental plantation depending on the temperature variation in Yaounde from February 2009 to January 2011 |
In 2009, for the third larval stage, the numerical variation showed seven peaks in February, March, May, July, August, November and January; in 2010, the six peaks were obtained in February, March, May, July, November and January 2011 (Fig. 4). In 2009, for the fourth larval stage, the numerical variations showed seven peaks in February, April, May, August, September, November and January; in 2010, the six peaks were observed in March, May, June, July, October and December (Fig. 5). In 2009, the numerical variations of the fifth instars larvae showed six peaks in February, May, July, September, November and January; in 2010, seven peaks were obtained in March, May, June, July, October, December and January 2011 (Fig. 5).
In 2009, the numerical variations of adults showed five peaks in February, May, November, December and January, but in 2010, seven peaks were obtained in February, March, May, July, September, October and December (Fig. 6). The number of generations was 5 in 2009 and 7 in 2010. In total, 564 adults were collected including 295 males and 269 females. The sex ratio is 1.09 and it is lightly in favor of males. For larval stages, we collected 878 larvae of first stage, 608 of second larvae stage, 456 of third larval stage, 332 of fourth larval stage and 262 larvae of fifth stage.
The statistical analysis showed that they was no significant difference between the average numbers of psyllids collected in 2009 and 2010 (Z = -0.36; P = 0.97 for the first larval stage; Z = -3.82; P = 0.70 for the second larval stage; Z = -2.44; P = 0.80 for the third larval stage; Z = -1.66; P = 0.09 for the fourth larval stage; Z = -1.37; P= 0.16 for the fifth larval stage, for adults, Z = -2.180; P = 0.03). Nevertheless statistical analysis showed a significant difference between the numerical variation of females in 2009 and 2010 (Z = -2.04; P = 0.04).
Considering the total average number of the five larvae of D. eastopi, the numerical variations showed in 2009 six peaks respectively in March, May, July, October, December and January; in 2010, six peaks were obtained in March, May, July, September, November and January 2011 (Fig. 7).
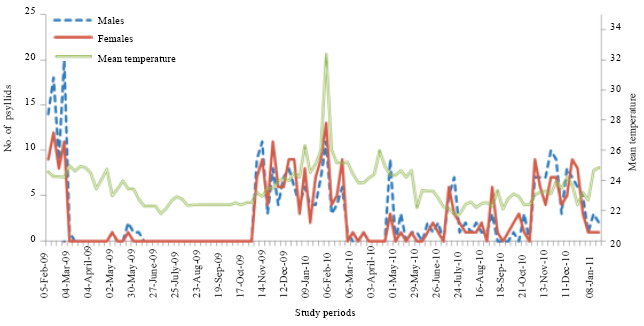 | |
| Fig. 6: | Numerical variations of D. harrisoni adults depending on the temperature variation in an experimental plantation in Yaounde from February 2009 to January 2011 |
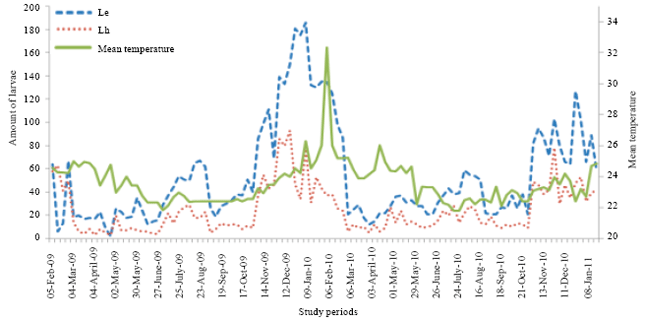 | |
| Fig. 7: | Comparison of numerical variations of different larval stages of D. eastopi (Le) and D. harrisonni (Lh) depending on the temperature variation in an experimental plantation in Yaounde from February 2009 to January 2011 |
Also, considering the total average number of the five larvae stage of D. harrisoni, the numerical variations in 2009 showed seven peaks, respectively in February, March, May, July, September, November and January; in 2010, seven peaks were obtained in March, May, July September, November, December and January 2011 (Fig. 7). Nymphs of D. eastopi were more abundant than those of D. harrisoni.
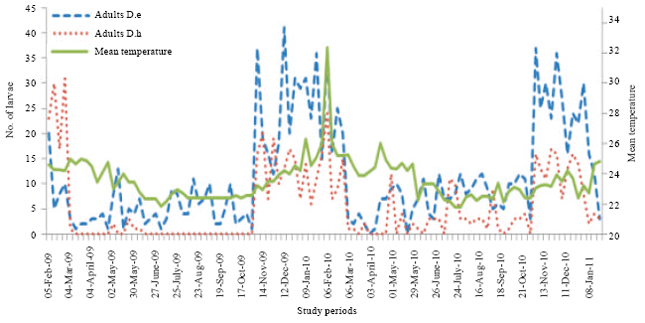 | |
| Fig. 8: | Comparison of numerical variations of D. eastopi (D.e) and D. harrisoni (D.h) adults depending on the temperature variation in an experimental plantation in Yaounde from February 2009 to January 2011 |
The numerical variation of adults shows six peaks of D. eastopi and five peaks of D. harrisoni. Diclidophlebia eastopi adults are more abundant than adults of D. harrisoni (Fig. 8). The cohabitation between the two species is in the advantage of D. eastopi.
The Wicolxon test comparing the adult average of D. eastopi and D. harrisoni showed in 2009 a significant difference (Z = -4.992; p<0.001) between those two species. In 2010, we also noticed a significant difference between D. eastopi and D. harrisoni (Z = -3.811; p<0.001).
Impact of abiotic factors on the numerical variation of D. eastopi and D. harrisoni: The ombrothermic diagram of Yaounde (Fig. 9) shows four seasons with two rainy seasons and two dry seasons: Rainy season: August-November and March-July; dry season: November-March and July-August. The two years of study (2009, 2010) we recorded on the first week of February 7 days which the temperature was more than 32°C.
Effects of temperature on the numerical variations of D. eastopi and D. harrisoni: The Spearman correlations between the numerical variation of individuals of Diclidophlebia eastopi and temperature is: R = 0.070; P = 0.480 for the first larval stage; R = 0.090; P = 0.360 for the second larval stage; R = 0.225; P = 0.020 for the third larval stage; R = 0.140; P = 0.140 for the fourth larval stage; R = 0.090; P = 0.370 for the fifth larval stage; R = 0.110; P = 0.260 for the males and R = 0.211; P = 0.030 for the females. Considering these values of Spearman correlation test (Table 1), we noticed that the Spearman correlation is positive but not significant except for the third larval stage and female. Then, the numerical variation of the third larval stage and female is correlated with annual temperature variation.
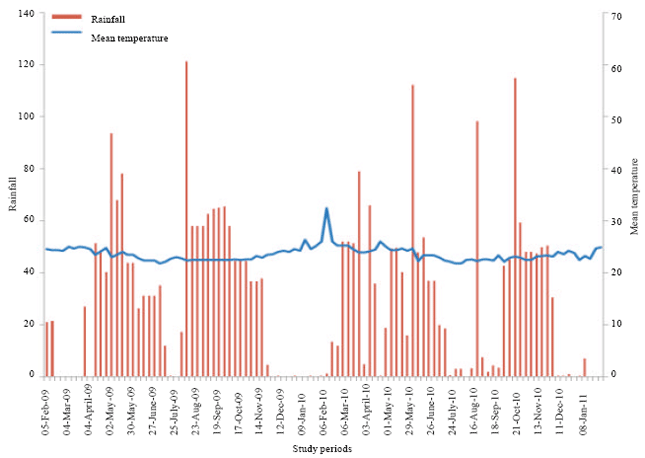 | |
| Fig. 9: | Rainfall and mean temperature of the study region from February 2009 to January 2011 (source: Weather Station of study site) |
| Table 1: | Spearman correlation between populations of Diclidophlebia eastopi and some climatic parameters of the study site from 2009 to 2011 |
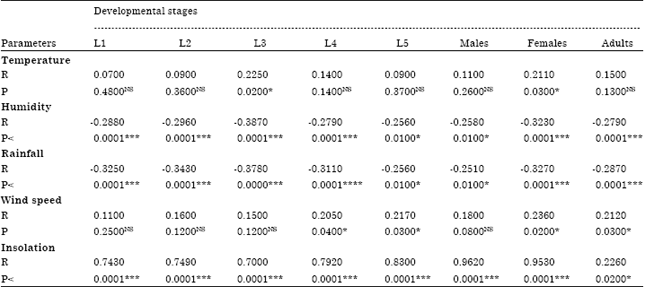 | |
| R: Spearman correlation coefficient, P: Probability value, NS: Not significant, *Significant correlation, **Highly significant correlation, ***Very highly significant correlation | |
| Table 2: | Spearman correlation between populations of Diclidophlebia harrisoni and some climatic parameters of the study site from 2009 to 2011 |
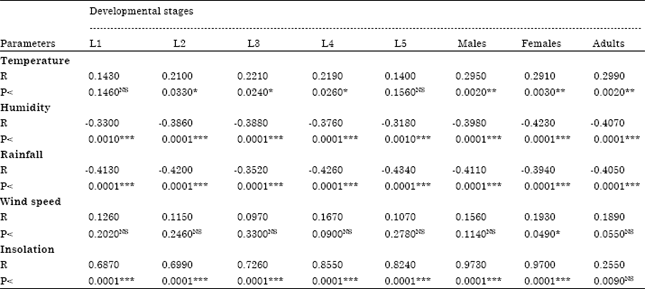 | |
| R: Spearman correlation coefficient, P: Probability value, NS: Not significant, *Significant correlation, **Highly significant correlation, ***Very highly significant correlation | |
The Spearman correlations between the numerical variation of each developmental stages of Diclidophlebia harrisoni and temperature is: R = 0.143; P = 0.146 for the first larval stage; R = 0.210; P = 0.033 for the second larval stage; R = 0.221; P = 0.024 for the third larval stage; R = 0.219; P = 0.026 for the fourth larval stage; R = 0.140; P = 0.156 for the fifth larval stage; R = 0.295; P = 0.002 for the males and R = 0.291; P = 0.003 for the females. Considering these values of Spearman correlation test (Table 2), we noticed that the Spearman correlation is positive but not significant with the first and fifth larval stages. It is significant with the others larval stages and adults. Then, the numerical variation of the others developmental stages of psyllid depends on the variation of temperature. Developmental stages of D. harrisoni are most susceptible to temperature variation than developmental stages of D. eastopi. Larval stages of D. eastopi could resist more than D. harrisoni larval stages.
Effects of relative humidity on the numerical variations of D. eastopi and D. harrisoni: The Spearman correlations between the numerical variation of individuals of Diclidophlebia eastopi and relative humidity is: R = -0.288; p<0.001 for the first larval stage; R = -0.296; p<0.001 for the second larval stage; R = -0.387; p<0. 001 for the third larval stage; R = -0.279; p<0.001 for the fourth larval stage; R = -0.256; p<0.010 for the fifth larval stage; R = -0.258; p<0.010 for the males and R = -0.323; p<0.001 for the females. According to Spearman correlation test (Table 1), we noticed that the correlation coefficient is negative and very highly significant with all larval stages and adults. Then the numerical variation of D. eastopi depends on the level of relative humidity; when the average number of relative humidity is higher, the number of psyllids could be low.
The Spearman correlations between the numerical variation of individuals of developmental stages of Diclidophlebia harrisoni and relative humidity is : R = -0.330; p<0.001 for the first larval stage; R = -0.386; p<0.001 for the second larval stage; R = -0.388; p<0.001 for the third larval stage; R = -0.376; p<0.001 for the fourth larval stage; R = -0.318; p<0.001 for the fifth larval stage; R = -0.398; p<0.001 for the males and R = -0.423; p<0.001 for the females. According to Spearman correlation test (Table 2), we noticed that the correlation coefficient is negative and very highly significant with all larval stages and adults. Then, the numerical variation of D. harrisoni depends on the level of relative humidity; when the average number of relative humidity is higher, the number of psyllids could be low.
Effects of rainfall on the numerical variations of D. eastopi and D. harrisoni: The Spearman correlations between the numerical variation of individuals of developmental stages of Diclidophlebia eastopi and rainfall is: R = -0.325; p<0.001 for the first larval stage; R = -0.343; p<0.001 for the second larval stage; R = -0.378; p<0.001 for the third larval stage; R = -0.311; p<0.001 for the fourth larval stage; R = -0.256; p<0.010 for the fifth larval stage; R = -0.327; p<0.001 for the females and R = -0.251; p<0.010 for the males. According to Spearman correlation test (Table 1), we noticed that the correlation coefficient is negative and highly significant with all larval stages and adults. Then, the numerical variation of D. eastopi depends on the level of the quantity of rainfall; when the quantity of rainfall is important, the number of psyllids could be low.
The spearman correlations between the numerical variation of individuals of developmental stages of Diclidophlebia harrisoni and rainfall is: R = -0.413; p<0.001 for the first larval stage; R = -0.420; p<0.001 for the second larval stage; R = -0.352; p<0.001 for the third larval stage; R = -0.426; p<0.001 for the fourth larval stage; R = -0.434; p<0.001 for the fifth larval stage and R = -0.405; p<0.001 for the adults. According to these values of Spearman correlation test (Table 2), we noticed that the correlation coefficient is negative and very highly significant with all larval stages and adults. Then, the numerical variation of D. harrisoni depends on the level of the quantity of rainfall; the number of psyllids is low when the rainfall is very high.
Effects of wind speed on the numerical variations of D. eastopi and D. harrisoni: The Spearman correlations between the numerical variation of individuals of adults stages and wind speed is: R = 0.212; P = 0.030 (for Diclidophlebia eastopi) (Table 1) and R = 0.189; p<0.05 (for D. harrisoni) (Table 2). According to these values of Spearman correlation test, we noticed that the correlation coefficient is positive and significant for D. eastopi and D. harrisoni adults. This suggests that when the wind speed is higher, their number counted on host plant were also higher. The wind speed influenced the numerical variation of adults of D. eastopi and D. harrisoni.
Effects of insolation on the numerical variations of D. eastopi and D. harrisoni: The spearman correlations between the numerical variation of individuals of developmental stages of D. eastopi and insolation is: R = 0.743; p<0.001 for the first larval stage; R = 0.749; p<0.001 for the second larval stage; R = 0.700; p<0.001 for the third larval stage; R = 0.792; p<0.001 for the fourth larval stage; R = 0.830; p<0.001 for the fifth larval stage; R = 0.226; p<0.020 for the adults. According to these values of Spearman correlation test (Table 1), we noticed that the correlation coefficient is positive and very highly significant with all the larval stages and adults. Then, the numerical variation of larvae and adults of D. eastopi depends on the variation of insolation.
The Spearman correlations between the numerical variation of individuals of developmental stages of D. harrisoni and insolation is: R = 0.687; p<0.001 for the first larval stage; R = 0.699; p<0.001 for the second larval stage; R = 0.726; p<0.001 for the third larval stage; R = 0.855; p<0.001 for the fourth larval stage; R = 0.824; p<0.001 for the fifth larval stage; R = 0.973; p<0.001 for males and R = 0.970; p<0.001 for females. According to these values of Spearman correlation test (Table 2), we noticed that the correlation coefficient is positive and very highly significant with all the larval stages and adults. Then, the numerical variation of larvae and adults of D. harrisoni depends on the variation of insolation.
 | |
| Fig. 10(a-b): | Leaves of T. scleroxylon with damages caused by D. eastopi, (a) Dorsal view showing larvae and floccular wax and (b) Turn to yellow |
 | |
| Fig. 11(a-b): | Leaves of T. scleroxylon with damages caused by D. harrisoni, (a) Initial coiling and (b) Fully wound |
Damages: Although, the two species lives together in the same plant, they cause different damages to their host. D. eastopi larvae while growing after egg hatched, causes discoloration of leaves, which’s turned from green to yellowish (Fig. 10). Feeding activities causes premature leaf fall, death of shoot apices and in severe cases, die back of the stems. D. harrisoni larvae feed on the young leaves and causes irreversible distortion of leaves (Fig. 11).
DISCUSSION
The study of the numerical fluctuations of psyllid population in Yaounde region permits us to identified in 2009 in the Yaounde region six peaks which could correspond to the six generations of D. eastopi psyllids populations and five peaks corresponding to the five generations of D. harrisoni psyllids populations. In 2010. the same number of generation was recorded with D. eastopi psyllids populations against seven generations of D. harrisoni. The number of generations of D. eastopi remained stable from one year to another. Otherwise, the number of generation of D. harrisoni varied from one year to another. In Nigeria, Osisanya (1974b) studied the population dynamic of the same psyllids species and recorded only three peaks corresponding to three generation all over the year. The number of generation varied from one region to another. Yana et al. (2009) after monthly survey in the Yaounde region, recorded three peaks which could explain not the number of generations but the main period of outbreaks of psyllid; the number of generations could be more express with weekly survey as we did during this study.
Females of D. eastopi seem lay their eggs on the lower face of young leaves. In the absence of young leaves, eggs were also laid on the lower face of old leaves of the host plant. After hatching, feeding activities of larvae cause fading and falling of young leaves. Females of D. harrisoni lay their eggs exclusively on young leaves. After hatching, the larvae cause leaf curl. The leaf curl is irreversible, because attacked leaves no longer return to their original shape; they turn yellow and drop prematurely. Our observations also indicate that periods of heavy outbreaks in 2009 were mid-November to late December, February for D. eastopi and mid-February to early March for D. harrisoni. In 2010. adults of D. eastopi showed main periods of outbreaks between late January and February, early November to late December. Regarding D. harrisoni, the outbreak periods was between early February and March, early November to late December.
However, despite these outbreaks, severe damage never results in the death of our experimental plants. These observations are consistent with those of Osisanya (1974b) on the same psyllid. Indeed, the work of Osisanya (1969) showed that the psyllid damage on plants and insolation is more severe when they are young. Temperature, relative humidity, rainfall, wind speed and insolation had an influence on the numerical variation of individuals of various stages of D. eastopi and D. harrisoni. Indeed, these climatic factors promote good growth of the host plants T. scleroxylon. The abundance of carbohydrates in the leaves would promote adult activity that proliferates at temperatures between 28 and 32°C. These observations confirm the findings of the work of Osisanya (1970). The host plant phenology could be the main factor influencing the population dynamic of the two psyllids of T. scleroxylon. The presence of youngest or oldest leaves is determinant to the development of psyllids. However, attacks decrease with age, as old trees tend to be more resistant. The integrated pest management will take into consideration the main flushing periods of the plant and the outbreak periods of two psyllids in the Yaounde region. A good knowledge of the biology of D. eastopi and D. harrisoni is important to ensure the effectiveness of chemical control and protection of seedlings and saplings. Similarly, the search for natural enemies of the psyllid is a major challenge for the limitation of natural populations of these pests.
CONCLUSION
Diclidophlebia eastopi and D. harrisoni are major pests of T. scleroxylon in Cameroon. Females of D. eastopi lay their eggs on the lower face of youngest and oldest leaves. After hatching, feeding activities of larvae cause fading and falling of young leaves. Females of D. harrisoni lay their eggs exclusively on youngest leaves of the host plant. After hatching, larvae cause irreversible leaf curl. Numerical fluctuations in 2009 showed six distinct generations of D. eastopi and five generations of D. harrisoni. In 2010. numerical fluctuations of psyllids population showed six generations for D. eastopi and seven for D. harrisoni. Abiotic factors such as temperature, relative humidity and rain fall and insolation had an impact on numerical fluctuations of two psyllids species in the Yaounde region. Since the presence of youngest or oldest leaves is very determinant to the development of psyllids, the host plant phenology could be the main factor influencing the population dynamic of the two psyllids on T. scleroxylon. The integrated pest management will take into consideration the main flushing periods of the plant and the outbreak periods of two psyllids in the Yaounde region. Host plant phenology also has an influence on the numerical variation of individuals of the two species. Indeed, periods of extensive leaf usually occur after the first rain and the end of the dry season should be particularly monitored.
ACKNOWLEDGMENT
We thank the Director of the National Support Agency Forestry Development (ANAFOR) in Yaounde and staff Woodyard Mbalmayo for technical assistance.
REFERENCES
- Aubert, B., M. Garnier, J.C. Cassin and Y. Bertin, 1988. Citrus greening disease survey in East and West African countries South of Sahara. Proceedings of the 10th Conference of International Organization of Citrus Virologists, November 17-21, 1986, Valencia, Spain, pp: 231-237.
Direct Link - Bonani, J.P., A. Fereres, E. Garzo, M.P. Miranda, B. Appezzato-Da-Gloria and J.R.S. Lopes, 2010. Characterization of electrical penetration graphs of the Asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomol. Exp. Applicata, 134: 35-49.
CrossRefDirect Link - Brennan, E.B., G.F. Hrusa, S.A. Weinbaum and W. Levison, 2001. Resistance of Eucalyptus species to Glycaspis brimblecombei (Homoptera: Psyllidae) in the San Francisco bay area. Pan-Pac. Entomol., 77: 249-253.
Direct Link - Burckhardt, D., 1987. Jumping plant lice (Homoptera: Psylloidea) of the temperate neotropical region. Part 2: Psyllidae (subfamilies diaphorininae, acizziinae, ciriacreminae and psyllinae). Zool. J. Linnean Soc., 90: 145-205.
CrossRefDirect Link - Burckhardt, D., 2005. Ehrendorferiana, a new genus of Neotropical jumping plant lice (Insecta: Hemiptera: Psylloidea) associated with conifers (Cupressaceae). Organ. Divers. Evolut., 5: 317-319.
CrossRef - Burckhardt, D., D.C. Alene, D. Ouvrard, J.L. Tamesse and J. Messi, 2006. Afrotropical members of the jumping plant-louse genus Diclidophlebia (Hemiptera: Psylloidea). Invert. Syst., 20: 367-393.
CrossRefDirect Link - Burckhardt, D. and D. Mifsud, 2003. Jumping plant-lice of the Paurocephalinae (Insecta, hemiptera, psylloidea): Systematics and phylogeny. Contrib. Nat. History, 2: 3-34.
Direct Link - Burckhardt, D. and D. Ouvrard, 2012. A revised classification of the jumping plant-lice (Hemiptera: Psylloidea). Zootaxa, 3509: 1-34.
Direct Link - Leon, J.H.D., M. Setamou, G.A. Gastaminza, J. Buenahora and S. Caceres et al., 2011. Two separate introductions of Asian citrus psyllid populations found in the American continents. Ann. Entomol. Soc. Am., 104: 1392-1398.
CrossRef - Dzokou, V.J., J.L. Tamesse and D. Burckhardt, 2009. Jumping plant-lice of the family psyllidae (Hemiptera: Psylloidea) from West-Cameroon: Biodiversity and host plants. J. Entomol., 6: 1-17.
CrossRefDirect Link - Hodkinson, I.D., 1974. The biology of the Psylloidea (Homoptera): A review. Bull. Entomol. Res., 64: 325-338.
CrossRefDirect Link - Marutani-Hert, M., W.B. Hunter and D.G. Hall, 2009. Establishment of Asian citrus psyllid (Diaphorina citri) primary cultures. In vitro Cell. Dev. Biol.-Anim., 45: 317-320.
CrossRefDirect Link - Marutani-Hert, M., W.B. Hunter and J.K. Morgan, 2011. Associated bacteria of asian citrus psyllid (Hemiptera: Psyllidae: Diaphorina citri). Southwestern Entomol., 36: 323-330.
CrossRefDirect Link - Milaire, H.G., 1987. La protection phytosanitaire des vergers de pommiers et de poiriers par la lutte integree [Phytosanitary protection of apple and pear orchards by integrated control]. Phytoma Defense des Cultures, 392: 38-48.
Direct Link - Nyeko, P., G. Edwards-Jones and R.K. Day, 2002. Population dynamics of herbivorous insects and potential arthropod natural enemies on Alnus species in Kabale district, Uganda. Agroforestry Syst., 56: 213-224.
CrossRef - Osisanya, E.O., 1970. Effect of shade on the rate of infestation of Triplochiton scleroxylon by Diclidophlebia species. Entomologia Experimentalis Applicata, 13: 125-132.
CrossRef - Osisanya, E.O., 1974. Aspects of the biology of Diclidophlebia eastopi Vondracek and D. harrisoni Osisanya (Homoptera, Psyllidae). Bull. Entomol. Res., 64: 9-17.
CrossRefDirect Link - Tamesse, J.L., J. Messi, T.X. Nguyen and S. Quilici, 1999. PrEsence de Trioza erytreae Del Guercio, le psylle des agrumes dans les principales zones ecoclimatiques du Cameroun [Presence of Trioza erytreae Del Guercio, the citrus psyllid in major eco-climatic zones of Cameroon]. Fruits, 54: 311-321.
Direct Link - Tiwari, S., A.D. Gondhalekar, R.S. Mann, M.E. Scharf and L.L. Stelinski, 2011. Characterization of five CYP4 genes from Asian citrus psyllid and their expression levels in Candidatus Liberibacter asiaticus-infected and uninfected psyllids. Insect Mol. Biol., 20: 733-744.
CrossRefDirect Link