
A. Koudamiloro
Africa Rice Center, 01 BP 2031, Cotonou, Benin
F.E. Nwilene
Africa Rice Nigeria Station, C/O IITA, PMB 5320, Ibadan, Nigeria
D. Silue
Africa Rice Center, 01 BP 2031, Cotonou, Benin
A. Togola
Africa Rice Nigeria Station, C/O IITA, PMB 5320, Ibadan, Nigeria
O. Oyetunji
Africa Rice Nigeria Station, C/O IITA, PMB 5320, Ibadan, Nigeria
Y. Sere
Africa Rice Tanzania Station, Dar-es-salam, Tanzania
M. Akogbeto
University of Abomey-Calavi, 01 B.P 4521 Cotonou, Benin
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 3 | Page No.: 153-162







ABSTRACT
Insects represent a major group of vectors of Rice yellow mottle virus (RYMV). This study aimed at identifying the main entomofauna transmitting RYMV to rice crop in Benin. Therefore, the transmission ability of 13 insect species was tested during the rice vegetative stages. These species belong to the Orthoptera order (Oxya hyla, Conocephalus longipennis, Paracinema tricolor, Acrida bicolor and Stenohippus aequus), the Coleoptera order (Chnootriba similis, Aulacophora foveicollis and Xanthadalia effusa), the Homoptera order (Cofana spectra, Nephotettix modulatus, Cofana unimaculata and Poophilis costalis) and the Diptera order (Diopsis thoracica). Among them, four species including P. tricolor, S. aequus, N. modulatus and P. costalis were identified for the first time as RYMV vectors. The species belonging to the Homoptera order appeared to be the most virulent, with higher values of viral titer. Virus distribution in the vector body parts was specific to each order. Basically, virus was more important in the Orthopteran, Coleopteran and Homopteran insects head part. It was also evident in the abdomen part of the Homopteran and the Orthopteran especially for P. tricolor, C. spectra and P. costalis. Disease severity was only observed in C. similis after inoculation and persisted until rice maturity with a score of 5 compared to the control which was 1 from a 1 to 9 scale. This study allowed identifying the presence of RYMV insect vectors in Benin among which, four are reported for the first time in Africa.
PDF Abstract XML References Citation
Received: November 05, 2013;
Accepted: February 27, 2014;
Published: March 28, 2014
How to cite this article
A. Koudamiloro, F.E. Nwilene, D. Silue, A. Togola, O. Oyetunji, Y. Sere and M. Akogbeto, 2014. Identification of Insect Vectors of Rice yellow mottle virus (RYMV) in Benin. Journal of Entomology, 11: 153-162.
DOI: 10.3923/je.2014.153.162
URL: https://scialert.net/abstract/?doi=je.2014.153.162
DOI: 10.3923/je.2014.153.162
URL: https://scialert.net/abstract/?doi=je.2014.153.162
INTRODUCTION
Rice represents 20% of consumed cereals behind sorghum (25%), corn (23%), millet (23%) and wheat which represents less than 10% in West Africa (Mendez del Villar et al., 2011).
In Africa, rice production was constrained by several diseases among which Rice yellow mottle virus (RYMV) is a main constraint (Sere et al., 2008; Ochola and Tusiime, 2011). RYMV is the principal viral disease of rice and was firstly reported in 1966 in Kenya (Bakker, 1970; Kouassi, 2008). It is endemic to Africa and is transmitted by a virus of the Sobemovirus genus (Rossel, 1986). In West Africa, it was found for the first time in 1975 (Raymundo and Buddenhagen, 1976), mostly in upland than in lowland ecologies (Awoderu, 1991). The disease is characterized by mottle and yellowing symptoms of variable intensities depending on the rice genotype and the virus pathotype. For some varieties, leaves become orange during the maturity of the infected plants. Other symptoms are partial emergence of the spikelet, partial or total sterility and significant reduction of the yield and even the death of the infected plant (Bakker, 1974; Raymundo and Buddenhagen, 1976; Awoderu, 1991). Similarly to all plant viruses, RYMV needs to move and periodically change hosts to ensure its life cycle and survival. The transmission of the disease is mechanical and can occur either during weeding, transplanting, friction between infected and healthy leaves or through insect damages (Abo et al., 2004; Uzest et al., 2011). Among all these transmission ways, insects are the most important (Sere et al., 2008; Nwilene et al., 2009; Onasanya et al., 2012). They were firstly considered as potential vectors because of their regular occurrence in the rice fields, but several studies have subsequently confirmed their vectorial capacity. Therefore, 30 insects were identified as vectors since the first identified by Bakker (1970). These included Chaetocnema pulla, Chnootriba similis, Trichispa sericea, Oxya hyla, Conocephalus longipennis, Paratettix sp., Zonocerus variegatus, Euscyrtus sp., Cofana spectra, Cofana unimaculata, Locris rubra and Locris maculata. They unintentionally transmitted the virus from rice to rice and from rice to alternative host plants, which are mostly gramineous (Nwilene, 1999; Nwilene et al., 2009). Currently, RYMV is persistently spreading in rice fields because of the high number of insect vectors specie, thus important yield losses are still recorded throughout the continent. Integrated Pest Management (IPM) an option taking into account the insect component requires the identification of the insect vectors in a given rice growing area that not yet inventoried. It is also important to determine the movement and distribution of the virus in the vectors’ body and to assess the influence of each vector on some phenotype characters of rice plant. These results will help to develop more efficient IPM strategies.
MATERIALS AND METHODS
Study site: All tests were performed in Benin at the Africa Rice site located at 12 km in northern Cotonou (6°25 N in latitude and 2°20 E in longitude), between November 2010 and December 2012.
Insect species: Thirteen insect species present in rice at the vegetative stage and mostly found in Benin were collected for the experiment (Table 1).
Rice genotype: Bouake 189, a susceptible rice variety to RYMV (Abo et al., 2002; Sere et al., 2008) was used.
Inoculation conditions: Two mesh-covered cages (0.8x0.8x1 m) described by Nwilene et al. (2009) with minor modifications were used. The first cage contained three pots where 3 seedlings were cropped for 3 weeks old. The second cage contained healthy plants, on which the insects from the first cage were transferred.
RYMV isolate: The same highly virulent (BE/10/14) strain was used for all insects. This strain was collected from the low land ecology in southern Benin. The first propagation was made on Bouake 189. For the purpose, two grams of infected leaves were collected and mechanically grinded in 0.01 M phosphate buffer pH 7.0 at a ratio of 1:10. The crushed product was homogenized and filtered using cheese cloth.
| Table 1: | Insects tested for the ability of the Rice yellow mottle virus transmission |
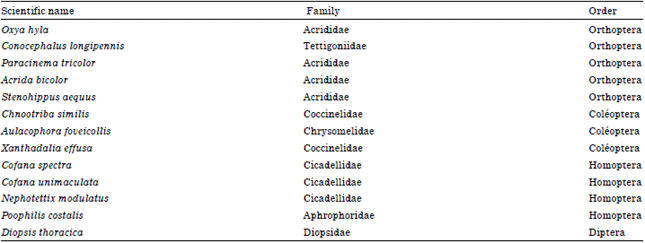 | |
The carborundum powder was added to this inoculum to induce leaves wounds and permit virus penetration into the leaves tissues. Inoculum was used just after its extraction.
Transmission tests for each insect specie: Two weeks after mechanically inoculating of seedlings of the first cage, 15 adult insects were introduced into it to allow them feed on infected plants for 48 h (to be sure that the insect were came into contact or fed on rice plant). Then, 10 insects coming from first cage were introduced in the second cage containing healthy seedlings, to test their ability to transmit RYMV. The remaining insects of first cage were sent to the laboratory to examine virus distribution in their body (head, thorax and abdomen). Three replications were performed for each insect specie.
Data collection: In the 2 cages, data on rice seedlings were collected at 28, 42 days After Inoculation (DAI) and at maturity. DAI were counted the day after mechanical inoculation took place in the first cage and the day after the ten insects were transferred in the second cage. Chlorophyll level (with SPAD 502 Chlorophyll meter) and disease severity (visual assessment) based on Standard Evaluation System (SES) on a scale of 1-9 (IRRI, 1988), were measured. For each insect, leaf samples were collected at 28 DAI in both cages and the corresponding insects were also collected two days after their stay in the first cage. Insects and leaf samples were stored in freezer (-20°C) for Antigen Coated Plate-Enzyme Linked Immunosorbent Assay (ACP-ELISA) tests.
Data analysis: Analysis of variance and comparison of means for the viral content, chlorophyll content, severity score and yield were performed using GenStat Discovery (GenStat, 2003). Means were separated by Student-Newman-Keuls (SNK) method.
RESULTS
Identification of new insect vectors: The viral titers obtained from leaves infested by insects were two fold higher than the negative control (0.08). Thirteen insects were statistically very different to the negative control (p<0.001). According to insect orders, Homopterans were found as the most virulent with P. costalis (0.41), C. spectra (0.51), C. unimaculata (0.49) and N. modulatus (0.60) while Orthopterans (O. hyla and C. longipennis), the least virulent ones.
| Table 2: | Viral titer by Optical Density (OD) at 405 nm (±SE) at 28 DAI in rice leaves for each insect |
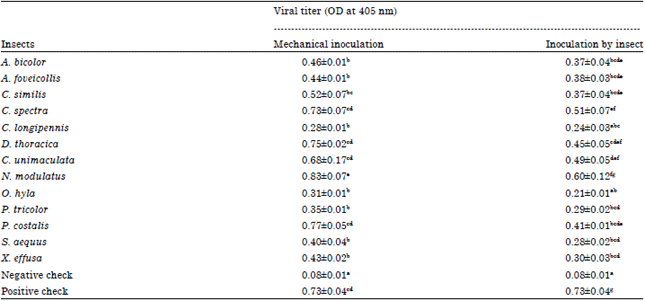 | |
| In the same column, means with different letter are significantly different at 5% level (Student Newman Keuls, p<0.001), Mechanical inoculation is the infected plants on which insects were firstly fed for 48 h, Thereafter, these insects were transferred on healthy plants (Inoculation by insect), Leaf samples were collected at 28 DAI (mechanical and by insect inoculation), DAI: Days After Inoculation, ±SE: Standard error | |
| Table 3: | Viral titer by Optical Density (OD) at 405 nm (±SE) in rice leaves for the four new insect vectors at 28 DAI |
 | |
| In the same column, means with different letter are significantly different at 5% level (Student Newman Keuls, p<0.001) | |
Out of these 13 insects, 4 were tested for the first time. These included P. tricolor (0.29), S. aequus (0.28), P. costalis (0.41) and N. modulatus (0.60) (Table 2). Their vectorial capacity has been confirmed by new tests (p<0.001) according the same protocol like the first tests. P. tricolor (0.29), S. aequus (0.26), P. costalis (0.28) and N. modulatus (0.29) belonged to the same statistical group with viral titer twofold higher than the control (0.11) (Table 3).
Determination of the movement and distribution of virus in the body part of the insect vectors: Two days after staying on mechanically inoculated seedlings, the virus was supposed to be preferentially localized in the body of the insect vector. Insects brought to the laboratory, were dissected into three parts (head, thorax and abdomen), crushed and subjected to ACP-ELISA (Fig. 1). Analyses of the body parts of insect vectors showed that there was no significant difference (p = 0.1) between head, thorax and abdomen for the location of virus for insect species. The location of the virus in the vector was specific to each insect order (p<0.001).
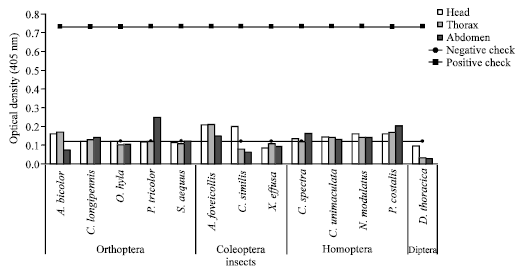 | |
| Fig. 1: | Virus localization and distribution into the body parts of the insect vectors. No significant difference (p = 0.1) between insect sections (head, thorax and abdomen) when considering insect species. The virus has lowly localized in the insect organism relative to the checks |
| Table 4: | Severity symptoms (±SE) and infection impact on the chlorophyll level (±SE) |
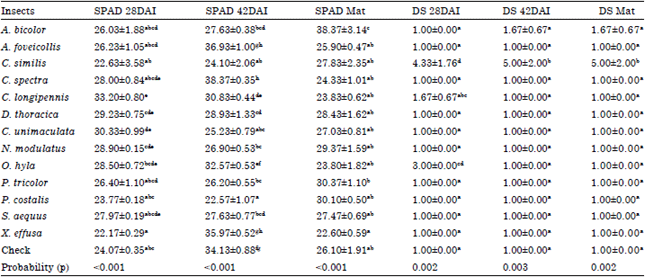 | |
| In the same column, means with different letter are significantly different at 5% level by SNK. Data was collected on inoculated plants by insects. Severity of symptoms was visual assessment of RYMV on the plant leaves based on Standard Evaluation System (SES) on a scale of 1-9, chlorophyll level concerned the evaluation of RYMV impact on the chlorophyll in the leaf samples, DS: Disease Severity, Mat: Maturity, DAI: Days After Inoculation, ±SE: Standard Error | |
Severity of RYMV symptoms and infection impact on the level of chlorophyll: The variation of the chlorophyll level was highly significant (p<0.001) at the three sampling dates (28 DAI, 42 DAI and maturity). At 28 DAI and maturity, the non-inoculated control, with SPAD value of 24.07 and 26.10, respectively, showed intermediate values of chlorophyll content. Concerning the disease severity, analyzes showed also a significant difference between the three dates: 28 DAI (p = 0.002), 42 DAI (p = 0.003) and maturity (p = 0.003). With only C. similis, RYMV symptoms were noticeable from 28 DAI to maturity (Table 4).
DISCUSSION
Identification of new insect vectors: The 13 insects were therefore confirmed as RYMV vectors by the ACP-ELISA test including four (P. tricolor, S. aequus, P. costalis and N. modulatus) for the first time in Africa. Previous studies had stated the vectorial capacity of 9 remaining insects (Bakker, 1971; Banwo et al., 2001; Nwilene et al., 2009). Homopterans which were the most virulent, are biting and sucking insects and are different from Orthoptera and Coleoptera which are chewing insects. They could collect the virus by introducing their stylets composed of two canals, directly into the host plant. At the same time, the first canal is used to suck the sieve while the second is used to inject virus particles into the host plant during feeding. They can transmit many pathogen agents to plant. This group transmits 80% of the diseases caused by arthropods (Giustina, 2002).
Indeed, Orthoptera and Coleoptera insects are able to collect the virus within their mouthparts and the virus does not need to be inside the insect to multiply (absence of salivary glands). These insects could transmit the virus collected by the mouthparts to a safe plant during the next feeding (Hebrard et al., 1999; Uzest et al., 2011).
The description of new insect vectors was done.
Nephotettix modulatus Melichar, 1912: Nephotettix (the green leafhopper) belongs to the Cicadellidae family and to the Deltocephalinae sub-family. The genus comprises several species included N. modulatus Melichar, N. virescens Distant, N. cincticeps Uhler and N. nigropictus Stal. The 3 last species are considered as important vectors of rice viral diseases including tungro, yellow dwarf, yellow-orange leaf and transitory yellowing, in Asia (Brugidou and Sire, 2002). In West Africa, two species (N. afer Ghauri and N. modulatus) were found on rice (Ghauri, 1968; Heinrichs and Barrion, 2004). The species found in Benin, N. modulatus (3-5 mm long) also called N. africanus Emeljanov, has green color with black marks on the wings and has a head with yellow scutellum. It is often confused with N. nigropictus. It has a separated black band on the head vertex that can be reduced at the middle (Wilson and Claridge, 1991). The life cycle is to 2-3 weeks with 4-10 days for the incubation period of eggs deposited on the leaf sheaths and 5 days for the nymphal stages. Nephotettix leafhoppers were distributed in all rice ecologies with a preference for the low land agrosystems. Apart from rice N. modulatus can be found on grasses such as Rottboellia cochinchinensis Clayton, Ischaemum rugosum Salisb and Paspalum vaginatum Swartz (Heinrichs and Barrion, 2004). Like all leafhoppers, it feeds on the aerial parts of the plant where it sucks plant sap from xylem and phloem by introducing its stylet into the sheaths. This causes direct damage to rice plants and transmission of pathogen such as viruses (Dale, 1994). However, it has been only recently shown to be a possible rice virus vector in West Africa (Heinrichs and Barrion, 2004). It is identified for the first time as RYMV vector in Benin.
Poophilus costalis Walker, 1851: Poophilus costalis belongs to the Aphrophoridae family. It was recorded as rice pest in Japan, India (Saini et al., 2011) and also in West Africa (Heinrichs and Barrion, 2004). Also called spittle bug, this froghopper feeds also on maize, sorghum, millet, sugarcane and other grass species (Ajayi, 2000; Shih et al., 2005). The adult is a small jumping insect of 9-11 mm long and is generally brownish. The nymphs always remain inside a foamy spittle mass. The anterior margin of the flat head is widely round with ovoid eyes. It has 3 black spots at mid length with subquadrate plate below these spots. The scutellum is brownish, flat with a pointed yellow apex and a median longitudinal yellow band. Legs are also brown (Teetes et al., 1995; Heinrichs and Barrion, 2004). Because of its damage mature plants become yellowing and wilting. Sometimes the damage leads to the death of seedlings (Bonzi, 1981). It was also reported as vectors of Pierce’s disease virus (Delong and Severin, 1950) and yellow leaf blotch of sorghum (Murty et al., 1994). For the first time it is identified as RYMV vector in Benin.
Paracinema tricolor Thunberg, 1815: Paracinema tricolor is an Orthopteran belonging to Acridae family and Oedipodinae sub-family. It includes three species where two are typically from Sub-saharan Africa which are P. tricolor and P. luculenta (Mestre and Chiffaud, 2006).
With a slight conical head, P. tricolor is a graminicole and granivorous insect. It has been found on rice in Benin (Mestre, 1988; Mestre and Chiffaud, 2006). It is greenish and can become brownish with green lateral faces on the body and also a brown dorsal face. Its pronotum has two lateral brown bands. The wings are hyaline or bluish. Its posterior tibia expanded in the apical part, are red bright. The antennae are filiform. Female and male are 32-40 mm and 24-29 mm long respectively. It flies accurately and has a strong preference to resettle on grass stem with the head in upright position and prefers wet habitat with tall grass. It is a chewing insect identified as a new vector of RYMV in Benin.
Stenohippus aequus Uvarov, 1926: This species belongs to the Acrididae family and to the Gomphocerinae sub-family. It includes 9 species among which 6 are present in Africa. S. aequus and S. xanthus were recorded in Benin (Fishpool and Popov, 1984; Mestre and Chiffaud, 2006). This Orthopteran insect is variably brownish with lateral or dorsal bands which are bright or dark. Its hyaline wings are occasionally slightly smoked at the apex. The hind tibiae are yellow to greyish that makes a big difference with Dnopherula spp where the tibiae are partially red. Its head is conical with laterals carinae of pronotum which are more or less marked. The antennas are filiform. The female and male are 16-27 and 13-20 mm long, respectively. It is the second chewing insect identified as a new vector of RYMV in Benin.
Determination of the movement and distribution of virus in the body part of the insect vectors: Concerning the distribution of virus inside the insect vectors (Head, thorax or abdomen), this study showed that the virus localization is not specific to insect species but specific to insect order. Generally virus is more important into the head in the three insect orders and into the abdomen to the Homoptera and Orthoptera orders (P. tricolor, C. spectra and P. costalis) as previously specified by Onasanya et al. (2012).
Severity of RYMV symptoms and infection impact on the level of chlorophyll: The physiological damages caused by insect vectors such as chlorophyll level and symptom severity on the rice plant are specific to and depend on infection impact of each insect. When the virus infection is not important, rice plants can overcome using its own mechanisms of defense and develop normally as healthy plants. But when infection becomes important, symptoms are observed up to maturity like infected plants by C. similis. However, it is reported that some varieties can still generate green leaves with very small and hardly perceptible streaks despite the attack by virus (Raymundo and Buddenhagen, 1976). Severity of symptoms in field conditions may also be affected by light intensity, day length, temperature and humidity (Bakker, 1974; Albar et al., 1995).
As mentioned by Nwilene et al. (2009), O. hyla feeds for a short period and is very mobile, thus its is able to induce very high phyllophagous activity. It can sometimes feed on all the primary tiller of the rice plant. The phenological stage of rice crop is affected, causing a delay in maturation. This was also observed with A. bicolor and C. similis where about all the leaves of rice plants were infested. It should also be noted that O. hyla incidence did not result in a high viral titer (0.21) compared to the control (0.73). Severity of leaf symptoms was not always considered as key factor in varieties assessment for resistance to RYMV as mentioned by Issaka (2000).
CONCLUSION
This study paved way for the identification of four insect vectors of RYMV, for the first time, in Africa (P. tricolor, S. aequus, N. modulatus and P. costalis). RYMV location in these vectors was specific to each insect order. The Homopterans were the most virulent. This result could lead future research to develop new management strategies to control the RYMV vectors.
ACKNOWLEDGMENTS
The authors would like to acknowledge Africa Rice Center and Benin Government for their financial and academic support. Our thanks also go to Dr George Gorgen for the certification of the identity of some insects, Dr Mounirou Sow, Mrs. Adronke OLUDARE whose comment helped to improve this manuscript and Mrs. Louisiane BACHABI for his technical help in the laboratory.
REFERENCES
- Abo, M.E., M.N. Ukwungwu and A. Onasanya, 2002. The distribution, incidence, natural reservoir hosts and insect vectors of Rice Yellow Mottle Virus (RYMV), genus Sobemovirus in Northern Nigeria. Tropicultura, 20: 198-202.
Direct Link - Ajayi, O., 2000. How Important are Spittle Bugs on Sorghum? In: Entomology in Nation Building: the Nigerian Experience, Proceedings of the ESN 30th Annual Conference, Dike, M.C., O. Ajayi, S.O. Okunade, N.O. Okoronkwo and A.A. Abba (Eds.). Entomological Society of Nigeria, Kano, Nigeria, pp: 91-97. Entomological Society of Nigeria, Kano, Nigeria, pp: 91-97.
- Awoderu, V.A., 1991. The rice yellow mottle virus situation in West Africa. J. Basic Microbiol., 31: 91-99.
CrossRefDirect Link - Bakker, W., 1970. Rice yellow mottle, a mechanically transmissible virus disease of rice in Kenya. Neth. J. Plant Pathol., 76: 53-63.
CrossRefDirect Link - Bakker, W., 1971. Three new beetle vectors of rice yellow mottle virus in Kenya. Neth. J. Plant Pathol., 77: 201-206.
CrossRefDirect Link - Banwo,O.O., R.H. Makundi, R.S. Abdallah and J.C. Mbapila, 2001. Identification of vectors of rice Yellow mottle virus in Tanzania. Arch. Phytopathol. Plant Protect., 33: 395-403.
CrossRef - Nwilene, F.E., 1999. Current status and management of insect vectors of Rice Yellow Mottle Virus (RYMV) in Africa. Insect. Sci. Applied, 19: 179-185.
CrossRef - Nwilene, F.E., A.K. Traore, A.N. Asidi, Y. Sere, A. Onasanya and M.E. Abo, 2009. New records of insect vectors of Rice Yellow Mottle Virus (RYMV) in Cote d'Ivoire, West Africa. J. Entomol., 6: 189-197.
CrossRefDirect Link - Ochola, D. and G. Tusiime, 2011. Survey on incidences and severity of rice yellow mottle virus disease in Eastern Uganda. Int. J. Plant Pathol., 2: 15-25.
Direct Link - Saini, M.S., N. Razak and I. Ahmad, 2011. Poophilus costalis walker (Hemiptera: Cercopoidea: Aphrophoridae): A possible constraint to commercial exploitation of Lavendula angustifolia mill in kashmir himalaya with affinity for c3 photosynthetic plants. J. Med. Plants Res., 5: 2278-2282.
Direct Link - Sere, Y., A. Onasanya, F.E. Nwilene, M.E. Abo and K. Akator, 2008. Potential of insect vector screening method for development of durable resistant cultivars to rice yellow mottle virus disease. Int. J. Virol., 4: 41-47.
CrossRefDirect Link - Delong, D.M. and H.H.P. Severin, 1950. Spittle-insect vectors of Pierce's disease virus. Hilgardia, 19: 339-381.
Direct Link - Shih, H.T., H.C. Tsay and J.T. Yang, 2005. Notes on the geographic distribution and host plants for Poophilus costalis (walker, 1851) (Hemiptera: Cercopoidea: Aphrophoridae). Plant Protect. Bull. (Taipei), 47: 171-178.
Direct Link - Brugidou, C. and C. Sire, 2002. Rice viruses: Current knowledge, agronomic impact and control methods. Virology, 6: 431-443.
Direct Link