
Nur Amin
Department of Plant Pests and Diseases, Hasanuddin University, South Sulawesi, Indonesia
La Daha
Department of Plant Pests and Diseases, Hasanuddin University, South Sulawesi, Indonesia
Nurariaty Agus
Department of Plant Pests and Diseases, Hasanuddin University, South Sulawesi, Indonesia
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 3 | Page No.: 142-152







ABSTRACT
Fungal endophytes are quite common in nature and some of them had been shown to have adverse effects against insects. The study aimed to isolate and identify of fungal endophytes from healthy pods of cocoa plant and to investigate their effects against the egg of cocoa pod borer (Conopomorpha cramerella (Snellen)) in the field as well as their endophytism on cocoa pods had been carried out. The results indicated that there were five fungal endophytes genera i.e., Trichoderma sp., Fusarium sp., Beauveria sp., Aspergillus sp. and one non-identified (mycelia sterilia) were found. All isolated endophytic fungus produced a negative effect on the survival of cocoa pod borer eggs but the greater effect were produced by Beauveria sp. and Aspergillus sp. treated in liquid form with the mean of 94 and 96% of egg mortality, respectively. Trichoderma sp., also produced a relatively high of 84 and 89% egg mortality for powder and liquid, respectively. Except Beauveria sp., Trichoderma sp., Fusarium sp. and Aspergillus sp. was found to penetrate in cocoa pod after two weeks of spraying. Beauveria sp., Trichoderma sp., Aspergillus sp. are endophytic fungus which provide potential biological control for cocoa pod borer.
PDF Abstract XML References Citation
Received: October 18, 2013;
Accepted: February 03, 2014;
Published: March 28, 2014
How to cite this article
Nur Amin, La Daha and Nurariaty Agus, 2014. The Study on the Role of Entomopathogenic Fungal Endophytes in Controling the Cocoa Pod Borer (Conopomorpha cramerella (Snellen)) (Lepidoptera: Gracillariidae) on Cocoa Plant. Journal of Entomology, 11: 142-152.
DOI: 10.3923/je.2014.142.152
URL: https://scialert.net/abstract/?doi=je.2014.142.152
DOI: 10.3923/je.2014.142.152
URL: https://scialert.net/abstract/?doi=je.2014.142.152
INTRODUCTION
The Cocoa Pod Borer (CPB) Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillaridae) is one of the most important limiting factors to cocoa production in Indonesia and Malaysia (Van Grinsven, 2003). Such various control tactics as insecticide application, sanitation, pruning and the use of ant predators had been practiced but not completely control obtained indicating that a new control tactic needed to keep the pest population under economic damage without an environmental negative impact produced.
In the course to seek an effective tactic control we come across that an interesting issue is the presence of fungal endophytes fungi on various such plant species as grasses and trees in various part of the world and the fungi seems to provide a protection of the plant hosts against such attacks by insect herbivores and pathogens. The term of endophyte was coined by the German scientist, Heinrich Anton De Bary in 1884 and used to define fungi and bacteria occurring inside plant tissues without causing any apparent symptoms in the host (Schulz et al., 2006). In the last few years fungal endophytes have been detected in hundreds of plants including such important agricultural commodities as wheat (Larran et al., 2002), bananas (Amin, 1994), maize (Amin, 2013a) tree palm oil. A number of authors have documented that the presence of endophytic fungi provide a protection of the plant hosts against insect herbivore (Clement et al., 2005), parasitic nematodes (Amin, 1994, 2013b; Elmi et al., 2000) and plant pathogens (Dingle and Mcgee, 2003; Wicklow et al., 2005). Other authors reported specifically the effects of endophytic fungi of Trichoderma atroviride MT-20 and S-2 as well as Fo162 added to the soil at sowing significantly reduced the number of white fly Trialeurodes vaporariorum, Aphis gossypii and Myzus persicae (Barahona, 2010). Variable effects however have sometimes been reported, e.g., Clement et al. (2005) reported different effects on two aphids, bird-cherry oat aphid, Rhopalosiphum padi (L.) and rose grass aphid, Metopopophium dirhodum (Walker) and the wheat stem sawfly Mayetiola destructor exposed to different wild barleys infected with Neotyphodium. Fewer studies have explored this relationship in nongrass systems. However, Jallow et al. (2004) reported drastic negative effects on larvae of Helicoverpa armigera (Hubner) reared on tomato plants infected with a nongrass endophyte, Acremonium strictum.
A such entomopathogens as Beauveria bassiana (Balsamo) Vuillemin (Ascomycota: Hypocreales) has been reported as an endophyte on maize (Cherry et al., 2004; Wagner and Lewis, 2000; Arnold and Lewis, 2005), tomato (Leckie, 2002), in the cocoa, Theobroma (Evans et al., 2003), (Mejia et al., 2008), in oil palm (Amin et al., 2008) and in bananas (Akello et al., 2007). The mechanisms of insect control displayed by endophytic fungi was correlated with toxin production (Lattanzio et al., 2006) and the production of fungal metabolites (Bush et al., 1997; Clay and Schardl, 2002). The traditional mode of infection of such fungal entomopathogens as B. bassiana involves spore deposition on the insect cuticle followed by formation of a germ tube which through enzymatic and mechanical action penetrates the cuticle (Arnold and Lewis, 2005). Once in the hemocoel, hyphal growth causes tissue damage and nutrient depletion. Some entomopathogenic fungi are also known to produce metabolites, but their involvement in insect toxicosis is still unknowed (Arnold and Lewis, 2005).
Based on our experience in the last few years in South Sulawesi which one of the most important cocoa production area in Indonesia, it is very oven that there are some trees with pods found to be free from cocoa pod borer infestations while the pods of other trees around suffer serious damage. Now a question arises what make those pods free from cocoa pod borer or diseases damage? Might the pods be associated with the presence of such endophytic microorganisms as fungi and or bacteria. In order to answer the question the study had been carried out with the aim to isolate the associated microorganism and then the trials also made to investigate their effect on the cocoa pod borer.
MATERIALS AND METHODS
Isolation of fungal endophytes: Endophytic fungi were isolated according the protocols described by Petrini (1986) which were slightly modified based on preliminary tests. The health cocoa pods taken from the field were washed twice in distilled water then surface sterilized by immersion for 1 min in 70% (v/v) ethanol, 5 min in sodium hypochlorite (2.5 % (v/v) available chlorine) and 30 seconds in 70% (v/v) ethanol and then washed three times in sterilized distilled water for 1 min each time. After surface sterilization, the samples were cut into 5-7 mm pieces and aseptically transferred to plates containing Potato Dextrose Agar (PDA, pH 6.8, containing (g L-1): potato 200; dextrose 20; agar 15.) which had been autoclaved for 15 min at 121°C and then aseptically supplemented with 100 mg mL-1 of chloramphenicol (Pfizer) to suppress bacterial growth. Aliquots from the third wash were plated onto PDA to check that surface sterilization had been effective and they were then incubated at 28°C. Any fungi present was isolated, purified and then maintained at 4°C on PDA slopes for further identification. After 5 days of incubation the grown fungi were identified with reference to Barnett and Hunter (1998) and Dugan (2006).
Production of fungal endophyte in powder form: Such fungal endophyte isolated as described previously propagated in rice medium containing chitin (1.0 g). The rice medium that has been soaked for 3 h put into a flask 100 g and autoclaved at 121°C for 30 min and after which by using a corkborer (diameter 0.5 mm), five pieces of endophytic fungi were inoculated in once the fungi started growing, the flask were shaken to assure an even fungal growth. The grown fungi then incubated at 30°C for 48 h. The rice medium along with the fungi then blended to produce a powder for further study.
Production of fungal endophyte in liquid form: The fungal grown obtained propagated on Potato Dextrose Broth (PDB) containing 1 g chitin to 100 mL PDB and autoclaved at 121°C for 30 min. By using a corkborer (diam. 0.5 mm), five pieces of endophytic fungi were inoculated in the flask were shaken to assure an even fungal growth. After 2 weeks the fungus in the flask filtered with the use of Whatman paper No. 1 and the supernatant keep into the refrigerator for use in further study.
The impacts of isolated fungal endophytes on the eggs of Cocoa Pod Borer (CPB): The study was carried out to observe the impacts of fungal endophytes application on the survival of CPB eggs. There were 6 fungal endophytes treatments and with 3 replications were involved and 18 experimental units were then prepared consequently. The experimental units consisted of a pod with naturally infested eggs so 18 eggs infested pods were selected. The number of eggs on a pod samples was recorded and then sprayed with 1x106 mL-1 suspension of fungal conidia. The treatment was made every day for 3 days period. The eggs were allowed to develop and after 1 week from the first application the eggs were inspected and un hatch eggs then recorded. The percentage of un hatch eggs were subsequently determined.
The study on the endophytisms of the fungal isolates: The study was carried out to observe whether the fungal endophyte applied might be able to penetrate into the treated pod. For each form (powder and liquid) of fungus, 18 pods consisted of 6 small, 6 moderate, 6 full size pods, respectively for replication were selected from cocoa trees. The cocoa pods were sprayed with a fungal endophyte with a conidial suspension of 1x mL-1. After two weeks of application the treated pods were then harvested and fungal reisolation then made with the same procedures as described in previous section.
RESULTS AND DISCUSSION
Isolation and identification of fungal endophyte isolates: The results indicated that there were 5 genera of fungal endophytes isolated from healthy pods of cocoa plants, i.e., Trichoderma sp., Fusarium sp., Aspergillus sp., Beauveria sp. and 1 (one) unidentified (mycelia sterilia) (Fig. 1-5). In other study in South Sulawesi in addition to Fusarium sp. and Aspergillus sp., Curvularia sp., Geotrichum sp., Gliocladium sp. and Colletotrichum sp., had been isolated from the health leaves of cocoa plants which resistant to VSD disease (Amin et al., 2014). A number of researches also reported the presence of endophytic fungi in cocoa plants.
 | |
| Fig. 1: | Mycelia sterelia |
 | |
| Fig. 2: | Fusarium sp. |
 | |
| Fig. 3: | Beauveria sp. |
 | |
| Fig. 4: | Trichoderma sp. |
 | |
| Fig. 5: | Aspergillus sp. |
Hanada et al. (2010) isolated a number of genera of fungal endophytes including Trichoderma, Fusarium from braches and twigs tissues of cocoa trees in Brasil. They found that Trichoderma and Fusarium showed a high level activity against Phytophthora palmivora, the causal agent of the black pod rod disease of cocoa. The presence of Trichoderma and Fusarium on the tissues of cocoa branches was also reported by Rubini et al. (2005) and Posada and Vega (2005) reported the establishment of entomopathogen Beauveria bassiana as endophytes of cocoa plants if the fungus is inoculated in radicles of seedlings. Three fungal endophyte isolated from healthy Theobroma cocoa tissue and screened in vitro for antagonism against major pathogen of cocoa (Mejia et al., 2008).
The impact of fungal endophytes on egg survival: The results indicated that all fungal endophytes studied produced a negative effect on the egg survival for both applied in powder and liquid form but a greater effects was recorded in liquid one in which the means of egg mortality ranging from 63 to 96% if the eggs treated with a liquid form compared to from 51 to 84% if treated with a powder form, all the means of which are different significantly from that of 10% produced by control (p = 0.05) (Table 1).
| Table 1: | Mean of egg mortality (%) treated with fungal endophytes in a powder and liquid forms |
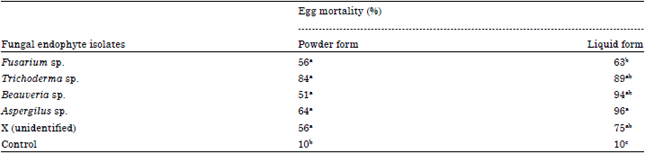 | |
| No. in the same column followed by same letters are not significantly different, (p = 0.05, Tukey’s test) | |
The means between fungal endophytes are not significantly different if they applied in the powder forms but in the liquid form, Aspergillus seems to produce the highest egg mortality (96%) is not significantly different from that of produced by Beauveria and Trichoderma but significantly different from that of produced by Fusarium and X isolate (p = 0.05) (Table 1). In general, endophytic Trichoderma sp., Beaveria sp. and Aspergillus sp., seem to be an important mortality factors for the cocoa pod borer egg which produced a relatively high mortality. Endophytic Trichoderma sp., showed a similar effect if it were applied in both powder and liquid forms with the mean of 84 and 89% egg mortality, respectively while a greater high mortality for Beauveria sp. and Aspergillus sp., application was only recorded in liquid forms with the mean of 94 and 96%, respectively compared with only 51 and 64% if they applied in the powder form (Table 1).
The mode of action of how endophytic fungi might protect plants from such insect herbivore attacks had been argued by many authors. Fungal endophytes grow systemically in intercellular spaces of the above-ground plant tissues including seeds and produce variety of alkaloids (Clay, 1998; Bultman et al., 2003) or fungal metabolites (Bush et al., 1997; Clay and Schardl, 2002). The production of alkaloid substances by endophytes-containing plants might protect the associated plants from herbivores due to antibiosis or feeding deterrence mechanisms (Crawford et al., 2010). An antibiosis substance is a toxic which might kill such victim as insects or microbial pathogens if they contact with or consume the substance. Examples of toxin production by endophytic fungi, notably those colonizing grasses are considerably abundant in the literature. Azevedo et al. (2000) reviewed that the protection of Canadian fir that against the spruce budworms resulted from the production of toxic secondary metabolites by endophytic fungi. In Prestidge and Gallagher (1988) and Pocasangre et al. (2000) reported a relationship between the presence of the fungus A. lolii in Lolium perenne and the growth, survival and feeding behaviour of Listronotus bonariensis larvae. In this case, the reduction in insect attacks towards infected plants was due to a strong toxin, lolitrem B, also toxic to mammals. This toxin, once added to insect diets, reduced insect growth and survival. Its assimilation occurs by ingestion but not by absorption through the insect integument.
Arnold and Lewis (2005) specifically discussed mode of action of Beauveria bassiana that the fungus deposits spore on the insect cuticle followed by formation of a germ tube and through enzymatic and mechanical action it penetrates the cuticle. Once in the hemocoel, hyphal growth causes tissue damage and nutrient depletion. Mode of action of isolates fungal endophytes on the eggs as found in the study might work in similar way.
The involvement of the enzymes in fungal endophyte mode of action as argued by Arnold and Lewis (2005) is supported by other authors. The production of chitinases by such endophytic fungi as Streptomyces sp., were reported by Haggag and Abdallh (2012) and Tang-Um and Niamsup (2012) and a number of fungal endophytes including Trichoderma sp., Beauveria sp., and Aspergillus sp. was reported by Matsumoto (2006).
Protease is an enzyme which capable of breaking down proteins while chitinase is an enzyme which capable of breaking down or digest chitins. The insect exoskeleton (insect cuticles) and egg shells (chorions) is mainly made of chitin and protein components and if the insects or eggs come contact with the such endophytic fungal product substances as digestive enzymes, the exoskeletons or chorions will degraded consequently and by a combination with an antibiosis substances will accelerate the dead of the insect or egg victim. In the case of the eggs, the chemical substances produced by fungus might also enter directly through the mycrophile, an egg hole for entrance of sperms.
It is assumed that the more rapidly action might take place depend on the such amount of the fungal substances produced in a time as performed by liquid form of fungal isolates studied which greatly reduced the egg survival compared to powder form except for Trichoderma sp., isolate. The enzyme production depend on such conditions as pH of culture, age of inoculums, type of fermentation, submerged or solid (Matsumoto, 2006). The enzymatic activities of Lecanicillium fungicola remarkably increased in liquid culture when pH varied from acid to alkali. The chitinolytic activity in filamentous as B. bassiana, Trichoderma harzianum, Aspergilus fumigates, Lecanicillium lecanii, Metarhizium anisopliae, Fusarium generally increases with culture time of incubation (Matsumoto, 2006). On the other side that synthesis of secondary metabolites by the plant have been shown to play an important roles in the interaction between plants and arthropods (Dicke et al., 2003; Turlings and Wackers, 2004; Van Poecke and Dicke, 2004; Arimura et al., 2005). Adults white ply evaluate the tactile and chemical cues of the plant surface after landing to determine the suitability of a plant as shelter or as a feeding and/or oviposition host (Walling, 2000). Changes in the synthesis of these cues stimulated by Fo162 also influence the behaviour of the insect on the Cucurbitacea plants hosts. The resulting effect increased plant fitness in a hostile environment as reported by Howe and Schaller (2008).
Reisolation of fungal isolates treated: The results indicated that after 2 weeks, Trchoderma sp. and Fusarium sp., were able to penetrate in the treated cocoa pods if fungus treated in both powder and liquid form. Aspergilus sp. and X isolates on the other hand the penetration only occurred if the fungus treated in the powder form and Beauveria sp. was not detected in the pod for period of two weeks (Table 2). It seems that Beauveria sp. might need more time in order to penetrate in the pod since Posada et al. (2010) reported that Beauveria bassiana is an endophyte in cocoa pods after two or three months inoculation on the flowers. The ability of the associated fungus to penetrate in the pod proofed that they could grow in the inter celuler of the pod and showed the ability of endophytism and the cocoa pods are conducive for the growth of naturally and inoculated fungal isolates consequently.
Indeed the research findings might over a great hope that endophytic fungus studied i.e., Aspergillus sp., Beauveria sp. and Trichoderma sp., seem to be potential for biological control agents of cocoa pod borer, a destructive cocoa pest in the worldwide. If the fungus suspensions sprayed on the cocoa pods, more than 90% the pod borer eggs might not be able to hatch and the young larvae emerging from the remaining unhatch eggs will bore and penetrate inside the pods and they will be then attacked or completely eliminated by the endophytic fungus applied earlier. Thus to obtain the best result from endophytic fungus application, the fungus should be sprayed as early as possible or when the cocoa trees produce a small pods (less than one month old) to enable the fungus colonized inside the pods.
| Table 2: | Reisolation of fungal endophyte isolate (%) in a powder and liquid forms |
 | |
When the pods gain a size of 9-10 cm in length (generally 3-4 months old), a suitable size for egg oviposition by cocoa pod borer, an intensive spray will be made but this is needed to be investigated.
CONCLUSION
It can be concluded that, Beauveria sp., Trichoderma sp. and Aspergillus sp. are endophytic fungus which provide potential as biological control agent for cocoa pod borer.
ACKNOWLEDGMENT
We would like to thank the Minister of the National Education and Culture, Republic of Indonesia for the financial support provided for the study, under the Contract of National Research Priority in Masterplan of acseleration and Extention of Indonesian Economic Development 2011-2025. We also expand our thank to the head of Hasanuddin University Research Institute for his valuable advice during the study.
REFERENCES
- Akello, J.T., T. Dubois, C.S. Gold, D. Coyne, J. Nakavuma and P. Paparu, 2007. Beauveria bassiana (Balsamo) Vuillemin as an endophyte in tissue culture banana (Musa spp.). J. Invertebr. Pathol., 96: 34-42.
CrossRefDirect Link - Arimura, G., C. Kost and W. Boland, 2005. Herbivore-induced, indirect plant defences. Biochim. Biophys. Acta, 1734: 91-111.
Direct Link - Azevedo, J.L., W. Maccheroni Jr., J.O. Pereira and W.L. de Araujo, 2000. Endophytic microorganisms: A review on insect control and recent advances on tropical plants. Electron. J. Biotechnol., 3: 40-65.
CrossRefDirect Link - Bultman, T.L., M.R. McNeil and S.L. Goldson, 2003. Isolate-dependent impacts of fungal endophytes in a multitrophic interaction. Oikos, 102: 491-496.
CrossRef - Bush, L.P., H.H. Wilkinson and C.L. Schardl, 1997. Bioprotective alkaloids of grass-fungal endophyte symbioses. Plant Physiol., 114: 1-7.
Direct Link - Cherry, A.J., A. Banito, D. Djegui and C. Lomer, 2004. Suppression of the stem-borer Sesamia calamistis (Lepidoptera; Noctuidae) in maize following seed dressing, topical application and stem injection with African isolates of Beauveria bassiana. Int. J. Pest Manage., 50: 67-73.
CrossRef - Clay, K., 1988. Fungal endophytes of grasses: A defensive mutualism between plants and fungi. Ecology, 69: 10-16.
Direct Link - Clay, K. and C. Schardl, 2002. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat., 160: S99-S127.
Direct Link - Clement, S.L., L.R. Elberson, N.A. Bosque-Perez and D.J. Schotzko, 2005. Detrimental and neutral effects of wild barley-Neotyphodium fungal endophyte associations on insect survival. Entomologia Experimentalis et Applicata, 114: 119-125.
CrossRef - Crawford, K.M., M.J. Land and J.A. Rudgers, 2010. Fungal endophytes of native grasses decrease insect herbivore preference and performance. Oecologia, 164: 431-444.
CrossRefDirect Link - Dicke, M., R.M.P. van Poecke and J.G. de Boer, 2003. Inducible indirect defence of plants: From mechanisms to ecological functions. Basic Applied Ecol., 4: 27-42.
CrossRef - Dingle, J. and P.A. Mcgee, 2003. Some endophytic fungi reduce the density of pustules of Puccinia recondita f. sp. tritici in wheat. Mycol. Res., 107: 310-316.
CrossRefDirect Link - Dugan, F.M. and F.M. Dugan, 2006. The Identification of Fungi: An Illustrated Introduction with Keys, Glossary and Guide to Literature. American Phytopathological Society, Saint Paul, MN, USA., ISBN: 9780890543368, Pages: 176.
Direct Link - Elmi, A.A., C.P. West, R.T. Robbins and T.L. Kirkpatrick, 2000. Endophyte effects on reproduction of a root‐knot nematode (Meloidogyne marylandi) and osmotic adjustment in tall fescue. Grass For. Sci., 55: 166-172.
CrossRefDirect Link - Evans, H.C., K.A. Holmes and S.E. Thomas, 2003. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog., 2: 149-160.
CrossRef - Hanada, E.H., A.W.V. Pomella, H.S. Costa, J.L. Bezerra, L.L. Loguercio, J.O. Pereira, 2010. Endophytic fungal diversity in Theobroma cacao (cacao) and T. grandiflorum (cupuacu) trees and their potential for growth promotion and biocontrol of black-pod disease. Fungal Biol., 114: 901-910.
CrossRef - Jallow, M.F.A., D. Dugassa-Gobena and S. Vidal, 2004. Indirect interaction between an unspecialized endophytic fungus and a polyphagous moth. Basic Applied Ecol., 5: 183-191.
CrossRef - Tang-Um, J. and H. Niamsup, 2012. Chitinase production and antifungal potential of endophytic Streptomyces strain P4. Maejo Int. J. Sci. Technol., 6: 95-104.
Direct Link - Larran, S., A. Perello, M.R. Simon and V. Moreno, 2002. Isolation and analysis of endophytic microorganisms in wheat (Triticum aestivum L.) leaves. World J. Microbiol. Biotechnol., 18: 683-686.
CrossRefDirect Link - Lattanzio, V., V.M.T. Lattanzio and A. Cardinali, 2006. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Res., 2006: 23-67.
Direct Link - Mejia, L.C., E.I. Rojas, Z. Maynard, S.V. Bael and A.E. Arnold et al., 2008. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control, 46: 4-14.
CrossRefDirect Link - Amin, N., 2013. Investigation of culture filtrate of endophytic fungi Nigrospora Sp. isolate Rs 10 in different concentrations towards root-knot nematode Meloidogyne Spp. Indian J. Sci. Technol., 6: 5177-5181.
Direct Link - Pocasangre, L., R.A. Sikora, V. Vilich and R.P. Schuster, 2000. Survey of banana endophytic fungi from central america and screening for biological control of the burrowing nematode (Radopholus similis). InfoMusa, 9: 3-5.
Direct Link - Posada, F. and F.E. Vega, 2005. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedlings (Theobroma cacao). Mycologia, 97: 1195-1200.
CrossRefDirect Link - Rubini, M.R., R.T. Silva-Ribeiro, A.W. Pomella, C.S. Maki, W.L. Araujo, D.R. Dos Santos and J.L. Azevedo, 2005. Diversity of endophytic fungal community of cacao (Theobroma cacao L.) and biological control of Crinipellis perniciosa, causal agent of witche's broom disease. Int. J. Biol. Sci., 1: 24-33.
CrossRefPubMedDirect Link - Van Poecke, R.M. and M. Dicke, 2004. Indirect defence of plants against herbivores: Using Arabidopsis thaliana as a model plant. Plant Biol., 6: 387-401.
Direct Link - Haggag, W.M. and E.G. Abdallh, 2012. Purification and characterization of chitinase produced by endophytic Streptomyceshygroscopicus against some phytopathogens. J. Microbiol. Res., 2: 145-151.
CrossRefDirect Link - Wagner, B.L. and L.C. Lewis, 2000. Colonization of corn, Zea mays, by the entomopathogenic fungus Beauveria bassiana. Applied Environ. Microbiol., 66: 3468-3473.
Direct Link - Walling, L.L., 2000. The myriad plant responses to herbivores. J. Plant Growth Regul., 19: 195-216.
CrossRefPubMedDirect Link - Wicklow, D.T., S. Roth, S.T. Deyrup and J.B. Gloer, 2005. A protective endophyte of maize: Acremonium zeae antibiotics inhibitory to Aspergillus flavus and Fusarium verticillioides. Mycol. Res., 109: 610-618.
Direct Link