
L.B. Silva
Programa de Pos-Graduacao em Agronomia Fitotecnia Universidade Federal do Piaui, Campus Profa. Cinobelina Elvas, Bom Jesus-PI, Brazil
G.S. Carvalho
Departamento de Ciencias da Natureza, Universidade Federal do Piaui, Campus Profa Cinobelina Elvas, Bom Jesus-PI, Brazil
A.C. Mancin
Departamento de Ciencias da Natureza, Universidade Federal do Piaui, Campus Profa Cinobelina Elvas, Bom Jesus-PI, Brazil
L.S. Silva
Departamento de Ciencias da Natureza, Universidade Federal do Piaui, Campus Profa Cinobelina Elvas, Bom Jesus-PI, Brazil
R.S.A. Nobrega
Centro de Ciencias Agrarias, Ambientais e Biologicas, Universidade Federal do Reconcavo da Bahia, Cruz das Almas-BA, Brazi
B.E. Pavan
Programa de Pos-Graduacao em Agronomia Fitotecnia Universidade Federal do Piaui, Campus Profa. Cinobelina Elvas, Bom Jesus-PI, Brazil
K. Maggioni
Programa de Pos-Graduacao em Agronomia Fitotecnia Universidade Federal do Piaui, Campus Profa. Cinobelina Elvas, Bom Jesus-PI, Brazil
E.M. Costa
Departamento de Ciencia do Solo, Universidade Federal de Lavras, Lavras-MG, Brazil
Journal of Entomology
Year: 2014 | Volume: 11 | Issue: 3 | Page No.: 111-126







ABSTRACT
Studies assessing the resistance of cowpea genotypes with respect to pest behavior are being developed and the selection of plants with insect resistance characteristics presents itself as an alternative to chemical control. The objective of the present study was to evaluate the behavior of Callosobruchus maculatus fed grains of Vigna unguiculata (L.) Walp (cv. "BR 17 Gurgueia") cultivated with five strains of symbiotic diazotrophic bacteria authorized by the Brazilian Ministry of Agriculture, Livestock and Supply (MAPA) for use as cowpea inoculants (UFLA 42C8; BR 3267; UFLA 0384; INPA 0311B; UFLA 48C5) and two control treatments (S/N, without nitrogen; C/N with nitrogen). Grain samples were infested with adult C. maculatus and maintained in laboratory conditions of 25°C and 12 h photophase. After 10 days of infestation the following insect behavioral and developmental parameters were evaluated: preference, number of grains perforated, number of grains with eggs, survival, emergence, insect weight, relative growth rate, consumption rate, conversion of ingested food, enzymatic activity of amylase, lipase, trypsin and chymotrypsin. The treatments most susceptible to attack by C. maculatus were: control C/N and UFLA 48C5, with greater attractiveness (28 and 23%, respectively) and survival (68 and 65%, respectively), while the most resistant were: INPA 0311B, UFLA 42C8 and control S/N, which reduced survival to 30, 24 and 19%, respectively.
PDF Abstract XML References Citation
Received: January 28, 2013;
Accepted: January 16, 2014;
Published: March 28, 2014
How to cite this article
L.B. Silva, G.S. Carvalho, A.C. Mancin, L.S. Silva, R.S.A. Nobrega, B.E. Pavan, K. Maggioni and E.M. Costa, 2014. Behavior of Callosobruchus maculatus Populations Fed with Vigna
unguiculata Grain Cultivated with Diazotrophic Bacteria Strains. Journal of Entomology, 11: 111-126.
DOI: 10.3923/je.2014.111.126
URL: https://scialert.net/abstract/?doi=je.2014.111.126
DOI: 10.3923/je.2014.111.126
URL: https://scialert.net/abstract/?doi=je.2014.111.126
INTRODUCTION
The cowpea (Vigna unguiculata L. Walp.), commonly called “feijão de corda” or “feijão macassar”, is a staple food of low-income populations in Northeastern Brazil. Among the different agricultural products found in tropical regions, the cowpea stands out for its high nutritional value, as well as its low cost of production. It is widely cultivated by smallholders, constituting a major component of the diet, especially in rural areas (Onwuliri and Obu, 2002; Almeida et al., 2010; Costa et al., 2011; Silva, 2012). In the state of Piauí, the cowpea occupies a prominent place in agriculture with an area of about 200 thousand hectares in first crop cultivation and 10 thousand hectares in second crop cultivation (Freire Filho et al., 2007).
The weevil Callosobruchus maculatus (Fabr.) (Coleoptera: Bruchidae) is a major pest of several species of Vigna, especially V. unguiculata, an important protein source in the tropics and subtropics. It can completely destroy grains after five months. The infestation by this bruchid starts in the field and continues in the storage, causing qualitative and quantitative losses. Currently the main method of control is by fumigation; however, resistant populations are quickly registered, resulting in control failures (Ahme et al., 2002; Manzoomi et al., 2010). Thus, several studies have attempted to select grains resistant to C. maculatus (IITA, 1990; Mbata, 1994; Seck, 1993; Chaves and Vendramim, 1995; Macedo et al., 2002; Farias et al., 2007; Melo et al., 2010), although few varieties with high resistance have been identified, hindering the development of resistant cultivars. The selection of plants with insect resistance characteristics presents itself as an alternative to chemical control, including for stored grain. Some studies have demonstrated the existence of cowpea genotypes resistant C. maculatus (Barreto and Quindere, 2000; Costa and Boica, 2004; Lima et al., 2001; Domingues et al., 2006; Marsaro and Vilarinho, 2011).
Grain resistance to insects has been attributed to diverse factors. In maize, the presence of phenolic compounds and amylase inhibitors was responsible for the resistance of genotypes to attack by Sitophilus zeamais, as demonstrated by Serratos et al. (1993) and Marsaro et al. (2005), respectively. In grain of Phaseolus vulgaris, the presence of the protein arcelin confers resistance in bean genotypes to attack by Zabrotes subfasciatus (Wanderley et al., 1997), while in the species V. unguiculata trypsin inhibitors are responsible for the resistance of cowpea genotypes to C. maculatus (Gatehouse et al., 1989).
Genetic manipulation can result in grains with large pools of nitrogenous metabolites and reduce the damage caused by bruchids. The availability of nitrogen for the production of defense metabolites, such as flavonoids, alkaloids, terpenes and sterols, could result in the selection of V. unguiculata varieties resistant to C. maculatus; however, this theory has not been widely investigated.
The storage of grain resistant to pests, after a few generations of these organisms, will be susceptible to attack, due to the adaptation process of insects in confined conditions. Parallel to the study of resistant varieties, it is necessary to identify the genes responsible for the trait, to determine the mechanism of resistance and focus on the stability of the characteristic in future generations. Within this context, the objective was to evaluate the development and behavior of populations of Callosobruchus maculatus fed grains of Vigna unguiculata (Cultivar BR (Brazil) 17 Gurguéia) cultured with different strains of symbiotic diazotrophic bacteria.
MATERIALS AND METHODS
Varieties of cowpea: The grain of cowpea (Vigna unguiculata (L.) Walp.) Cv. BR 17 Gurguéia was produced by Costa et al. (2011) in the experimental area at the Federal University of Piauí (UFPI), Professor Cinobelina Elvas campus (CPCE), Bom Jesus PI (Piauí) (09°04'28'' south latitude, 44°C21'31'' west longitude and altitude of 277 m). Cultivation took place during the period May to June 2009 in a yellow latosol of medium texture in an area without prior use of any type of diazotrophic bacterial inoculant. The grain came from seven specific treatments (nitrogen source): BR 17 S/N (without nitrogen), BR 17 C/N (with nitrogen), BR 17 42C8 (inoculated with nitrogen fixing strain UFLA (Universidade Federal de Lavras) 03-154-42C8), BR 17 BR 3267 (inoculated with nitrogen fixing strain BR 3267), BR 17 UFLA 0384 (inoculated with nitrogen fixing strain UFLA 0384), BR 17 INPA (Instituto de Pesquisas da Amazônia) 0311B (inoculated with nitrogen fixing strain 0311B INPA) and BR 17 UFLA 48C5 (inoculated with nitrogen fixing strain UFLA 48C5).
The grain after harvest was stored in plastic bags at a temperature of -20°C (in freezer) for 15 days to remove the insects from the field. Prior to the experiments, the grain was removed from the freezer, placed in plastic containers covered with fine cloth and kept in the laboratory for six days to achieve moisture content equilibrium. The adults of C. maculatus used were obtained from a population maintained in the Laboratory of Entomology UFPI/CPCE.
Free-choice feeding test: In the choice test, one hundred (100) C. maculatus adult insects (age 1-5 days) were placed in an arena containing seven interconnected compartments: a central for the insects and the others containing 50 g of grain of each treatment and five replicates per treatment. After 24 h of infestation the interconnections between the compartments were closed with cotton, to record the number of individuals in each compartment.
No-choice feeding test: The no-choice test was conducted in plastic pots (50 g), each with 10 g of grains of each cowpea treatment and infested with 30 adults of C. maculatus. After 10 days, the insects were removed (alive or dead) and weighed. The beans were weighed and their content determined to estimate the mass lost to insect consumption. Five replicates were used for each treatment. Later nutritional indices of C. maculatus were calculated for each treatment as described by Isman et al. (1990) and Huang et al. (1997): Relative growth rate (daily weight gain per insect weight), relative consumption rate (daily consumption rate per insect weight) and conversion efficiency of ingested food ((relative growth rate/relative consumption rate) x100).
Enzymatic analysis: For the analysis of enzymatic activity, three random samples of 100 non-sexed adult insects reared on cowpea grains from each of the seven treatments were collected, immersed in 1.5% KCl solution and macerated in 6.0 mL of Tris-HCl 0.1 M pH 8.0. The homogenate was filtered with cotton gauze and centrifuged at 10,000 g max for 15 min. The precipitate was discarded and aliquots of the supernatant (enzyme extract) were retained for determination of total protein and enzymatic activity of amylase, lipase, trypsin and chymotrypsin.
For the amylase assay a kit from BIOCLIN (Quibasa-Basic Chemistry Ltda, Belo Horizonte, Minas Gerais, Brazil) containing substrate and color reagent was used. The reaction is based on fixed time kinetics (Caraway). The enzyme extract was incubated with the starch substrate and the reading was done in a UV-Vis spectrophotometer at a wavelength of 660 nm.
For the lipase assay the BIOCLIN kit (Quibasa-Basic Chemistry Ltda, Belo Horizonte, Minas Gerais, Brazil) containing buffer, enzyme inhibitor, color reagent, substrate and acetone was used. The reading was performed using a UV-Vis spectrophotometer at a wavelength of 410 nm.
Aliquots of the supernatant were dissolved in Tris-.05 M, pH 8.0 buffer and the final assay volume adjusted to 300 μL with the above mentioned buffer. After ten minutes of incubation at 37°C, 1.0 mL of BAPNA solution (N-benzoyl-D, L-arginine-ñ-nitroanilide) (0.4 mg mL-1) dissolved in Tris HCl 0.05 M, pH 8.0 buffer was added to the mixture and incubation continued for another 30 min at 37°C. The reaction was quenched by addition of acetic acid 30% (v/v) and substrate hydrolysis by the enzyme was monitored spectrophotometrically at 410 nm. The calculation of the activity of trypsin was made by determining the residual activity of trypsin in assay, in BAPNA in nmol mg-1 protein min-1.
Aliquots of the supernatant were dissolved in Tris-0.1 M, pH 8.0 buffer and the final assay volume adjusted to 500 μL with the above mentioned buffer. After pre-incubation for 10 min at 37°C, 30 μL of SAAP (N succinyl Ala-Ala-Pro-Phe p-nitroanilide) (0.3 mg mg mL-1) substrate dissolved in pure DMSO (dimethylsulfoxide) was added to the mixture and the incubation continued for another 30 min at 37°C. The reaction was stopped by adding acetic acid 30% (v/v) and substrate hydrolysis by the enzyme was monitored spectrophotometrically at 405 nm. The calculation of chymotrypsin activity was done by determining the residual activity of chymotrypsin in assay in SAAP nmol mg-1 protein min-1.
The protein concentration in the enzyme extracts was determined by the method of Warburg and Christian (1941) for determination of specific activity.
Statistical analysis: The data from the choice and no-choice tests and enzymatic activities were analyzed by analysis of variance (SAS Institute, 2002), followed by univariate analysis of variance and Tukey’s HSD test, when appropriate (SAS Institute, 2002).
Data for each trait were subjected to multivariate analysis of mean Euclidean distance to obtain the dissimilarity matrix among the bean treatments. Groupings were developed from the matrix generated by the UPGMA hierarchical and Tocher optimization methods. In parallel, an analysis was done for the same characteristics divided into two groups-nutritional content and enzyme activity. In the first group, 7 features were considered (Relative Growth Rate (RGR), Relative Consumption Rate (RCR), conversion efficiency of ingested food (ECI), % survival, number of insects and insect weight) and in the second group 4 features (activities of amylase, lipase, trypsin and chymotrypsin). These data, by group, were subjected to analysis of variance to obtain treatment means by characteristic and matrix of variance and residual covariance among traits to proceed with the multivariate analysis of Mahalanobis distance to obtain the dissimilarity matrix among treatments. Having the dissimilarity matrices, groupings were done with the UPGMA hierarchical and Tocher optimization methods; multivariate analyzes were performed with the aid of the Genes computational program (Cruz, 2006).
RESULTS
In the choice test insects were attracted to all treatments (grain grown with different nitrogen sources); however, there were significant differences in the percentage of insects attracted to each treatment within 24 h, therefore there were fewer eggs and emerged insects. The treatment INPA 0311B was less attractive (6.6%) and the treatment S/N (without added nitrogen) had fewer eggs and emerged insects (3.4) (Table 1). The most attractive treatment was C/N (28%), which had the greatest number of oviposits and emerging insects, signifying that the insects performed well with this treatment.
The percentage survival and nutritional indices were calculated for the C. maculatus population exposed for 10 days to cowpea grain from plants grown with strains of nitrogen-fixing bacteria and two controls, with and without addition of mineral nitrogen. The objective was to see if the insects differ as to the conversion of ingested food and strategies to minimize or compensate for grain quality as a food source.
The grain from plants grown with the strains INPA 0311B and UFLA 42C8 and from plants grown without the addition of mineral nitrogen (S/N) reduced the survival of C. maculatus to 30, 24 and 19%, respectively (Fig. 1).
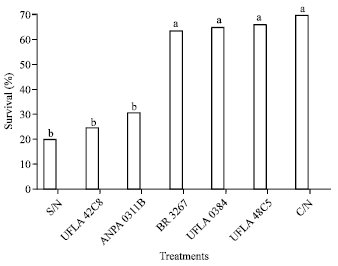 | |
| Fig. 1: | Survival (%) of adults of one strain of cowpea weevil (Callosobruchus maculatus) maintained on beans grown with different nitrogen sources. Means followed by the same letter do not differ in the least significant difference Tukey’s test (p<0.05). S/N-without nitrogen; C/N-with nitrogen; rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
| Table 1: | Mean percentage (±SE) of attractiveness to C. maculatus of grains from different cowpea treatments in free-choice test after 24 h of exposure |
 | |
| 1 S/N: Without nitrogen, C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia),* Means followed by the same letter do not differ by Tukey’s test (p<0.05) | |
Analyzing survival, number of individuals that emerged and body mass of insects in the no-choice bioassay, it can be said that in the treatment with grain from bean plants grown with the addition of mineral nitrogen (C/N), there is a greater percentage of weevil survival (68%) and the conditions are more favorable for development in comparison with the other treatments.
According to the analysis of variance of nutritional parameters for C. maculatus, there is a significant difference in relative growth rate (F = 74.83, p<0.0001), conversion efficiency of ingested food (F = 7.5, p<0.0001), relative consumption rate and activity of digestive enzymes (amylase and lipase) as a function of treatment (Table 2; Fig. 3, 4 and 5). The greatest amylase activity was seen in the treatment C/N (142 amylase dL mg-1 of protein). This explains the greater consumption (RCR: Fig. 3), survival and emergence (Fig. 2). In the treatment with added nitrogen (C/N), the insect’s performance was better, probably due to the availability of metabolic constituents needed for development.
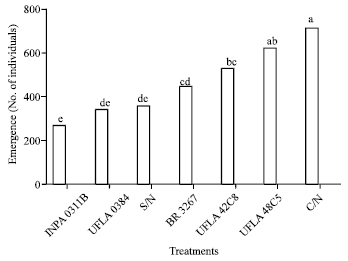 | |
| Fig. 2: | Emergence (No. of individuals) of adults of one strain of cowpea weevil (Callosobruchus maculatus) maintained on beans grown with different nitrogen sources. Means followed by the same letter do not differ in the least significant difference Tukey’s test (p<0.05). S/N: Without nitrogen, C/N: With nitrogen; rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
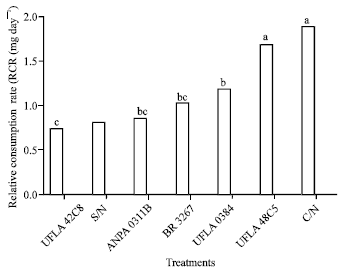 | |
| Fig. 3: | Relative consumption rate (RCR (mg day-1) of one strain of cowpea weevil (Callosobruchus maculatus) maintained on grains grown with different nitrogen sources. Means followed by the same letter do not differ by the least significant difference Tukey’s test. S/N: Without nitrogen, C/N: With nitrogen; rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
The trypsin activity of homogenate of insects maintained on grain from treatment BR 3267 was lower (63 nmol mg-1 min-1) compared to the other treatments and for the activity of chymotrypsin was no significant difference (Table 2).
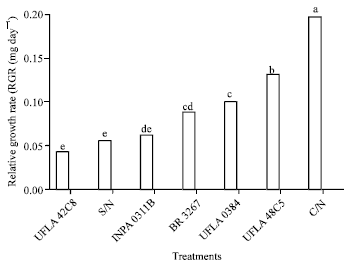 | |
| Fig. 4: | Relative growth rate (RGR (mg day-1) of one strain of cowpea weevil (Callosobruchus maculatus) maintained on grains grown with different nitrogen sources. Means followed by the same letter do not differ by the least significant difference Tukey’s test. S/N: Without nitrogen; C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
| Table 2: | Activity of energy and digestive metabolism enzymes (Mean±SEM) in a population of C. maculatus maintained on grain grown with different nitrogen sources |
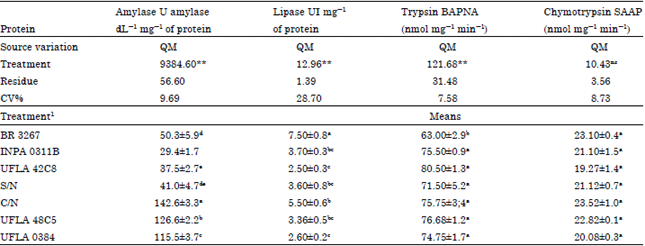 | |
| 1 S/N: Without nitrogen, C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia, UI: Lipase units, BAPNA: Ná-Benzoyl-L-arginine 4-nitroanilide hydrochloride, SAAP: Succinyl ALA-ALA-PRO-PHE p-Nitranilide), QM: Mean of squares, CV%: Coefficient of experimental variation, ns not significant ** e * significant at 1% and at 5% probability by F test, means followed by the same letter in the columns do not significantly differ by Tukey’s test (p<0.05) | |
The relative growth rate (Fig. 4) of insects exposed to the treatments C/N and UFLA 48C5 (0.195±0.006 and 0129±0.004 mg day-1, respectively) was higher than for the other treatments (S/N, UFLA 0384, BR 3267 and INPA 0311B). A similar result was observed for the relative consumption rate (Fig. 3) 1.881±0.006 and 1.678±0.009 mg day-1 for C/N and UFLA 48C5, respectively vs. 0.853±0.010, 0.787±0004 and 0.726±0002 mg day-1, for S/N, UFLA 0384 and BR 3267, respectively.
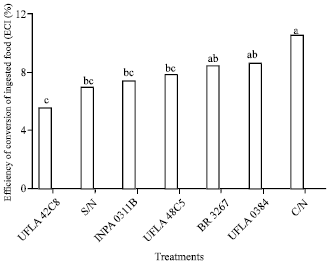 | |
| Fig. 5: | Efficiency of conversion of ingested food (ECI(%)) of one strain of cowpea weevil (Callosobruchus maculatus) maintained on grains grown with different nitrogen sources. Means followed by the same letter do not differ by the least significant difference Tukey’s test. S/N: Without nitrogen; C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
This outcome was expected as a function of non-adaptation to the food source to which the insects were exposed (no-choice test). The variation in results as to efficiency in the conversion of ingested food (%) (Fig. 5) was basically determined by the growth rate of C. maculatus. In the treatment C/N, greater consumption and ECI (efficiency of conversion of ingested food) were observed and consequently a greater growth rate. In contrast, the treatment UFLA 42C8 was associated with a lower consumption rate (RCR), growth rate (RGR) and efficiency of conversion of ingested food than the other treatments.
The percentage of attractiveness and mortality achieved in each treatment reflects its differential susceptibility to grain and non-adaptation to the food source. The consumption rate of the group of insects confined on grains obtained from plants without the addition of mineral nitrogen decreased, in contrast to the treatment with addition of mineral nitrogen, noting that similar results were observed in the treatment inoculated with the symbiotic diazotrophic bacteria strain UFLA 48C5. In the treatment with the strain 0311B INPA, there is less consumption, survival and conversion of ingested food, which means less damage to stored cowpea grains. The efficiency of conversion of ingested food, which is derived from the relative growth rate and the rate of consumption, may be associated with the accumulation of nitrogen in the grain, the presence of nutritional factors and the absence of substances toxic to the insect.
Regarding the number of grains perforated, the data are consistent with the growth rates. For the treatment C/N (with added nitrogen) there was a larger number of grains perforated, an average of 12.6±2.2 and 5.6±0.5 for the treatment UFLA 42C8. Callosobruchus maculatus has a significantly altered feeding behavior, in relation to the content and accumulation of nitrogen in cowpea grains, as well as insect growth, consumption and efficiency in the conversion of food into body mass.
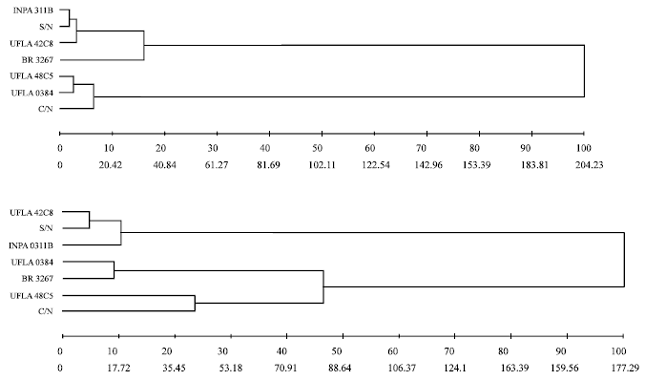 | |
| Fig. 6: | Dendrogram obtained by the UPGMA clustering method constructed from the dissimilarity matrix of seven cowpea treatments by the Mahalanobis method for enzymatic activity characteristics, above and nutritional indices, below (line above distance % of total distance, line below absolute distance). S/N: Without nitrogen; C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
Lipase and amylase showed significant differences in their activities among the seven treatments evaluated (F = 57.39, glerro = 6, p<0.0001 for amylase) (Table 2). The enzyme extract from the insect groups fed on with grains from plants grown with the addition of mineral nitrogen (C/N) and the strains UFLA 0384 and UFLA 48C5 had higher amylase activity. In the analyses of lipase activity (F = 9.80, glerro = 6, p<0.0001), insects fed with grains from the treatment inoculated with the strain BR 3267 showed higher lipase activity and lower trypsin activity; chymotrypsin activity did not differ between treatments.
Figure 6 shows the dendrograms obtained from the dissimilarity matrix among the cowpea treatments, based on enzymatic activity and nutritional indices. The dendrograms grouped the bean treatments similarly, indicating that the nutritional indices of the insects are linked to the enzymatic activity, since the grouping of the treatments coincided. In the dendrogram for enzyme activity the formation of two groups can be seen, one constituted by the treatments INPA 311B, S/N (without added nitrogen), UFLA 42C8 and, more distant from other representatives of this group, BR 3267. The two groups were cohesive in that within each group, with the exception of the treatment BR 3267, the treatments did not separate by more than 10% of the total distance available. Thus, treatments within each group are similar to the enzymatic activity.
| Table 3: | Grouping of cowpea treatments by the Tocher optimization method, from the dissimilarity matrix obtained by the Mahalanobis method for enzymatic activity and nutritional indices and mean Euclidean distance for all features together |
 | |
| 1 S/N: Without nitrogen, C/N: With nitrogen, rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras, BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) | |
Comparing the groups by the hierarchical and optimization methods in Table 3, based on the same characteristics, it is confirmed that treatment BR 3267, despite being less divergent from group 1, forms a separate group from the others. Thus, it can be stated that the treatment BR 3267 is different from all other treatments, although it presents a behavior closer to the first group.
Comparing the two groups in terms of enzymatic activity and nutritional indices, the treatments INPA 311B, S/N (without added nitrogen) and UFLA 42C8 were grouped together by the optimization method and were very close in the dendrograms, with percentage distance not exceeding 10% of the absolute distance. The treatments C/N (with added nitrogen) and UFLA 42C5 also clustered together by all methods and traits.
When the grouping was done based on all the traits of these treatments, once again they grouped together, demonstrating the similarity of the traits analyzed. The treatment BR 3267 stands out for its lack of standardization in the groupings; sometimes behaving similarly to treatment group 1 and other times group 2. However, by the grouping method of Tocher for nutritional indices it formed a group with treatment UFLA 0384, showing that this treatment also did not show a pattern of behavior similar for all the traits analyzed.
In Fig. 7, it is seen that the treatment UFLA 0384 was only linked to the treatments C/N (with added nitrogen) and UFLA 48C5 from 70% of the absolute distance, while by the Tocher method it categorized in group 1, underlining the absence of a pattern of dissimilarity according to the characteristics analyzed.
DISCUSSION
As for the attractiveness of the treatments, INPA 0311B was less attractive, but presented favorable conditions for oviposition and insect emergence compared to the treatment without added nitrogen (S/N). This can be explained by the low nutritional quality as a function of the nitrogen source. Smiderle and Schwengber (2008) reported that the yield and quality of cowpea grain increases as a function of N levels.
In the treatment with added mineral nitrogen, the growth rate of C. maculatus was superior to others, which is explained by the research of Costa et al. (2011), who found that the cultivation of V. unguiculata with added mineral nitrogen results in a greater accumulation of nitrogen in the cowpea grain. The efficiency of conversion of ingested food, which is obtained from the relative growth rate and the rate of consumption, can be associated with the accumulation of nitrogen in the grain, the presence of nutritional factors and the absence of substances toxic to the insect.
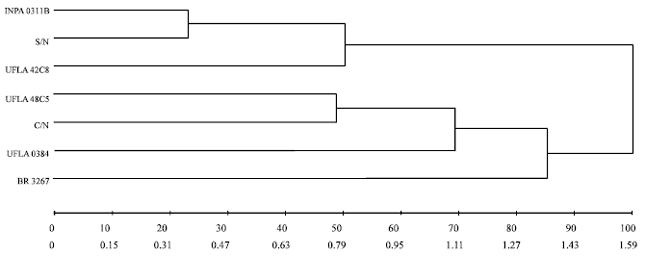 | |
| Fig. 7: | Dendrogram obtained by the UPGMA clustering method constructed from the dissimilarity matrix of seven cowpea cultivars of by the Euclidean distance method, average for the characteristics enzymatic activity and nutritional indices (line above distance % of total distance, line below absolute distance). S/N: Without nitrogen, C/N: With nitrogen; rhizobium strains recommended by the Brazilian Ministry of Agriculture: UFLA 42C8, BR 3267, UFLA 0384, INPA 0311B, UFLA 48C5 (UFLA: Universidade Federal de Lavras; BR: Brazil, INPA: Instituto Nacional de Pesquisas da Amazônia) |
These results are important from an economic standpoint because Costa et al. (2011) found that the productivity of V. unguiculata in the treatment inoculated with INPA 0311B resulted in grain yields equivalent to control fertilized with mineral nitrogen and higher than average for the state of Piaui. In the treatment INPA 0311B and control treatment C/N (added mineral nitrogen) there is a greater accumulation of nitrogen in the grain, higher grain production and high resistance to attack and development of C. maculatus. These finding are probably due to the presence of substances toxic to the insect, a consequence of the accumulation of nitrogen in the grain.
The resistance of cowpea cultivars can be supplanted by biotypes of C. maculatus, capable of altering the plant-insect relationship. While the use of the plant by the pest is maintained at low levels for several generations, the intensity of the selection pressure would produce a population of insects adapted, as evidenced by Shade et al. (1996). Such a change in the response of C. maculatus to the resistant cultivar TVu 2027 was demonstrated by Dick and Credland (1986). In C. maculatus populations from Yemen and Nigeria, adaptation of the insects occurred rapidly, leading within three generations to the evolution populations able to survive on this cultivar. Thus, maintaining resistance for prolonged periods will be constrained by the appearance of other adapted strains (Lima et al., 2001).
To increase the resistance of grain cultivars, studies seek to understand the mechanisms of resistance found in native plants or in the bruchids themselves that attack a particular crop plant but not another (Sales et al., 2000). In this study the focus was to determine whether there was a difference in the response of C. maculatus to grain grown with different nitrogen sources and it was found that grain with higher nitrogen content, mineral and/or derived from biological fixation, is more susceptible to the development of the bean weevil and that interference occurs in the activity of energy and digestive metabolic enzymes.
It was found that the activity of amylase and lipase were inhibited by the treatments INPA 0311B, UFLA 42C8 and S/N (without added nitrogen) and according to Sales et al. (2000), both the common bean and the cowpea have defensive compounds that protect seeds against different herbivores. Among these are tannins, cyanogenic glycosides, non-protein amino acids and proteins such as protease and inhibitors of amylase, lectins, chitinases and B-1,3-glucanases. These defensive compounds are ineffective against the host-specific bruchids Callosobruchus maculatus and Zabrotes subfasciatus that attack cowpea and common bean, respectively.
The use of plants resistant to insect attack represents a promising alternative to reduce damage, considering that plants produce chemical defenses, such as protease inhibitors, that can inhibit insect enzymes (Kuroda et al., 1996). Insects of the family Bruchidae have as their main digestive enzymes serine and cysteine protease. According to the results presented in this study, it was seen that there was a reduction in enzyme activity, which entails the mobilization of energy sources for the normal development of C. maculatus. The grains of cowpea plants grown with different nitrogen sources are assimilated differently by the insects.
Various types of plant proteins have been identified as being toxic to different groups of insects, for example plant lectins. The insects discriminate grains containing these proteins. Factors such as hardness of grain, seed size, lack of nutritional factors and presence of toxic substances may decrease the damage of C. maculatus to cowpea seeds. According to the results presented in this study the variety BR 17 Gurguéia, when inoculated with the strain INPA 0311B, produces grain more resistant to attack by C. maculatus under storage conditions. The grain contain sugars, organic acids, amino acids, proteins and peptides, flavonoids, anthocyanins, alkaloids, terpenes and steroids, with a greater amount of nitrogen available, it will be possible to produce more of these molecules that can participate in the defense mechanisms of the plant. Varieties of V. unguiculata resistant to bruchids are rich in phenolics, alkaloids and terpenes (Ndakidemi and Dakora, 2003).
Guzman-Maldonado et al. (1996) in relating the physical and chemical characteristics of varieties of the common bean (Phaseolus vulgaris L.) showed deterrent activity as a function of the amount of lectin present in the grain, reducing the number of insects per unit during grain storage. The insects, through their sensory system, are capable of detecting substances harmful to their metabolism/development. It is well documented that the sensory system of insects is very important for regulating their behavior, reproduction and survival, including location of food, partner and determination of oviposition sites (Hekmat-Scafe et al., 2002; Ban et al., 2003). Messina et al. (1987a, b) report that the labial and maxillary palps of C. maculatus are the main organs used for discrimination of host and oviposition behavior. The chemoreceptors are mediated by soluble proteins that are present in the lymph chemo-sensillae. A particular type of pest exposed to a particular variety of resistant grain, in the case of stored grain pests, initially refuses the food source due to non-adaptation, but in confinement, after a few generations, begins a process of adaptation to the food source. These characteristics of some insects can be used to develop new concepts for insect control.
A study developed with cowpea achieved significant grain yields using inoculants of nodule-forming bacteria in a process known as nitrogen fixation (Almeida et al., 2010; Costa et al., 2011). In the study it was found that treatments with nodule-forming bacterial inoculants for biological nitrogen fixation produced grain yields similar to treatment with mineral nitrogen, with higher resistance to attack by C. maculatus.
One of the strategies to develop plant varieties resistant to insect pests is to select plants that possess characteristics that interfere with or reduce the growth of insects. As noted in this study, the reduction in amylase activity is an important factor, because it interferes with the absorption of carbohydrate reserves present in the grain. Many studies have utilized amylase inhibitors, proteins, as a strategy in the study of new biopesticides (Macedo et al., 2004; Murad et al., 2006; Bonavides et al., 2007).
The treatments with greater emergence in the no-choice test were those most preferred in the choice test. This fact may be associated with chemical factors such as the release of substances attractive for oviposition. Potrich et al. (2010) reports that the attractiveness of S. zeamais can be associated with the content present, or released by the grain such as odor and flavor.
CONCLUSION
It is important to conduct a more detailed physical and chemical study of cowpea grain resulting from V. unguiculata grown with different nitrogen sources. Depending on the amount of nitrogen available in the soil during cultivation and the availability of this element stored in the grain, substances that inhibit weevil feeding may be present in greater or lesser amounts in the grain. Examples are arcelin, which confers resistance to Z. subfasciatus and trypsin inhibitors responsible for the antibiosis. A less preferred cultivar in a condition of free choice becomes susceptible and shows increased damage when the insect has no other food source. This highlights the importance of nutritional indices and technological quality, in that it will be possible to observe which cultivar is more resistant to insect attack.
ACKNOWLEDGMENT
We gratefully acknowledge the National Counsel of Scientific and Technological Development (CNPq) for granting the scientific initiation scholarship to Banco do Nordeste and financial support this study.
REFERENCES
- Ahme, S., M.A. Khan and N. Ahmad, 2002. Determination of susceptibility level of phosphine in various strains of Dhora (Callosobruchus maculatus F.). Int. J. Agr. Biol., 4: 329-331.
Direct Link - Almeida, A.L.G., R.M.C.M. Alcantara, R.S.A. Nobrega, J.C.A. Nobrega, L.F.C. Leite and J.A.L. Silva, 2010. Produtividade do feijao-caupi cv BR 17 Gurgueia inoculado com bacterias diazotroficas simbioticas no Piaui. Rev. Bras. de Cienc. Agrarias, 5: 364-369.
CrossRefDirect Link - Ban, L., A. Scaloni, A. Brandazza S. Angeli, L. Zhang, Y. Yan and P. Pelosi, 2003. Chemosensory proteins of Locusta migratoria. Insect Mol. Biol., 12: 125-134.
CrossRef - Barreto, P.D. and M.A.W. Quindere, 2000. Resistance of cowpea genotypes to the weevil. Pesqui. Agropecu. Bras., 35: 779-785.
Direct Link - Bonavides, K.B., P.B. Pelegrini, R.A. Laumann, M.F. Grossi-de-Sa and C. Bloch Jr. et al., 2007. Molecular identification of four different α-amylase inhibitors from baru (Dipteryx alata) seeds with activity toward insect enzymes. J. Biochem. Mol. Biol., 40: 494-500.
PubMedDirect Link - Costa, E.M., R.S.A. Nobrega, L.V. Martins, F.H.C. Amaral and F.M.S. Moreira, 2011. Nodulation and yield of Vigna unguiculata (L.) Walp. by strains of rhizobia in Bom Jesus, PI. Rev. Cienc. Agron., 42: 1-7.
CrossRef - Costa, N.P.C. and A.L. Boica Jr, 2004. Effect of cowpea genotypes, Vigna unguiculata (L.) Walp., on the development of Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae). Neotrop Entomol., 33: 77-83.
CrossRefDirect Link - Dick, K.M. and P.F. Credland, 1986. Changes in the response of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) to a resistant variety of cowpea. J. Stored Prod. Res., 22: 227-233.
CrossRef - Domingues, S.J.S., F.R. Melo, J.M. Aguiar, A.G. Affonso and J.S.A. Giuli et al., 2006. Resistance of Vigna unguiculata (cowpea) seeds to Callosobruchus maculatus is restricted to cotyledonary tissues. J. Sci. Food Agric., 86: 1977-1985.
CrossRefDirect Link - Farias, L.R., F.T. Costa, L.A. Souza, P.B. Pelegrini and M.F. Grossi-de-Sa et al., 2007. Isolation of a novel Carica papaya α-amylase inhibitor with deleterious activity toward Callosobruchus maculatus. Pestic. Biochem. Physiol., 87: 255-260.
Direct Link - Gatehouse, A.M.R., S.J. Shackley, K.A. Fenton, J. Bryden and A. Pusztai, 1989. Mechanism of seed lectin tolerance by a major insect storage pest of Phaseolus vulgaris, Acanthoscelides obtectus. J. Sci. Food Agric., 47: 269-280.
CrossRefDirect Link - Guzman-Maldonado, S.H., A. Marin-Jarillo, J.Z. Castellanos, E.G. De Mejia and J.A. Acosta-Gallesgosc, 1996. Relationship between physical and chemical characteristics and susceptibility to Zabrotes subfasciatus (Boh.) (Coleoptera: Bruchidae) and Acanthoscelides obtectus (Say) in common bean (Phaseolus vulgaris L.) varieties. J. Stored Prod. Res., 32: 53-58.
CrossRef - Hekmat-Scafe, D.S., C.R. Scafe, A.J. McKinney and M.A. Tanousy, 2002. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res., 2: 1357-1369.
PubMedDirect Link - Huang, Y., J.M.W.L. Tan, R.M. Kini and S.H. Ho, 1997. Toxic and antifeedant action of nutmeg oil against Tribolium castaneum (Herbst) and Sitophilus zeamais Motsch. J. Stored Prod. Res., 33: 289-298.
CrossRefDirect Link - Kuroda, S., C. Tokunaga, Y. Kiyohara, O. Higuchi and H. Konishi et al., 1996. Protein-protein interaction of zinc finger LIM domains with protein kinase C. J. Biol. Chem., 271: 31029-31032.
PubMedDirect Link - Lima, M.P.L., J.V. Oliveira, R. Barros and J.B. Torres, 2001. Identification of Genotypes of Cowpea Vigna unguiculata (L.) Walp. Resistant to Callosobruchus maculatus (Fabr.) (Coleoptera: Bruchidae). Neotrop. Entomol., 30: 289-295.
CrossRefDirect Link - Macedo, M.L.R., M.M. Castro and M.G.M. Freire, 2004. Mechanisms of the insecticidal action of TEL (Talisia esculenta lectin) against Callosobruchus maculates (Coleoptera: Bruchidae). Arch. Insect Biochem. Physiol., 56: 84-96.
CrossRefDirect Link - Macedo, M.L.R., M.G.M. Freire, J.C. Novello and S. Marangoni, 2002. Talisia esculenta lectin and larval development of Callosobruchus maculatus and Zabrotes subfasciatus (Coleoptera: Bruchidae). Biochim. Biophys. Acta, 1571: 83-88.
CrossRefPubMedDirect Link - Manzoomi, N., G.N. Ganbalani, H.R. Dastjerdi and S.A.S. Fathi, 2010. Fumigant toxicity of essential oils of Lavandula officinalis, Artemisia dracunculus and Heracleum persicum on the adults of Callosobruchus maculates (Coleoptera: Bruchidae). Munis Entomol. Zool., 5: 118-122.
Direct Link - Marsaro Jr., A.L., S.M.N. Lazzari, E.L.Z. Figueira and E.Y. Hirooka, 2005. Amylase inhibitor in corn hybrids as a resistance factor for Sitophilus zeamais Motschulsky (Coleopt era: Curculionidae). Neotrop Entomol., 34: 443-450.
CrossRef - Melo, R.A., V.A. Forti, S.M. Cicero, A.D.L.C. Novembre and P.C.T. Melo, 2010. Use of X-ray to evaluate damage caused by weevils in cowpea seeds. Hortic. Bras., 28: 472-476.
CrossRef - Messina, F.J., J.L. Barmore and J.A.A. Renwick, 1987. Oviposition deterrent from eggs of Callosobruchus maculates: Spacing mechanism or artifact? J. Chem. Ecol., 13: 219-226.
CrossRef - Messina, F.J., J.L. Barmore and J.A.A. Renwick, 1987. Host selection by ovipositing cowpea weevils: Patterning of input from separate sense organs. Entomologia Experimentalis et Applicata, 43: 169-173.
CrossRefDirect Link - Murad, A.M., R.A. Laumann, T.D.A. Lima, R.B.C. Sarmento and E.F. Noronha et al., 2006. Screening of entomopathogenic Metarrhizium anisopliae isolates and proteomic analysis of secretion synthesized in response to cowpea weevil (Callosobruchus maculatus) exoskeleton. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 142: 365-370.
CrossRefPubMed - Ndakidemi, P.A. and F.D. Dakora, 2003. Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Funct. Plant Biol., 30: 729-745.
Direct Link - Onwuliri, V.A. and J.A. Obu, 2002. Lipids and other constituents of Vigna unguiculata and Phaseolus vulgaris grown in Northern Nigeria. Food Chem., 78: 1-7.
CrossRefDirect Link - Sales, M.P., I.R. Gerhardt, M.F. Grossi-de-Sa and J. Xavier-Filho, 2000. Do legume storage proteins play a role in defending seeds against bruchids? Plant Physiol., 124: 515-522.
Direct Link - Serratos, J.A., A. Blanco-Labra, J.A. Mihm, L. Pietrzak and J.T. Arnason, 1993. Generation mean analysis of compounds in maize grain and susceptibility to maize weevil Sitophilus zeamais infestation. Can. J. Bot., 71: 1176-1181.
CrossRefDirect Link - Shade, R.E., L.W. Kitch, P. Mentzer, L.L. Murdock, 1996. Selection of a cowpea weevil (Coleopera: Bruchidae) biotype virulent to cowper weevil resistant landrace TVu 2027. J. Econ. Entomol., 89: 1325-1331.
Direct Link - Silva, R., 2012. Project with cowpea leads to new perspectives bordering the lower Madeira. Brazilian Agricultural Research Corporation-Embrapa, July 24, 2012. http://www.embrapa.br/imprensa/noticias/2012/julho/4a-semana/projeto-com-feijao-caupi-leva-novas-perspectivas-aos-ribeirinhos-do-baixo-madeira/.
- Isman, M.B., O. Koul, A. Luczynski and J. Kaminski, 1990. Insecticidal and antifeedant bioactivities of neem oils and their relationship to azadirachtin content. J. Agric. Food Chem., 38: 1406-1411.
Direct Link