
A.H. Fouly
Department of Agricultural Zoology, Faculty of Agriculture, Mansoura University, Mansoura 35516, Egypt
O.A. Nassar
Department of Agricultural Zoology, Faculty of Agriculture, Mansoura University, Mansoura 35516, Egypt
M.A. Osman
Department of Agricultural Zoology, Faculty of Agriculture, Mansoura University, Mansoura 35516, Egypt
Journal of Entomology
Year: 2013 | Volume: 10 | Issue: 4 | Page No.: 199-206







ABSTRACT
Eusieus scutalis (A.-H.), is a common phytoseiid mite in Egypt. However, the present study aims to throw more light on the biology and life table parameters of E. scutalis which was reared under laboratory conditions of 26°C and 70% R.H., on palm pollen, immature stages of Tetranychus urticae koch and Bemisia tabaci Genn. Life cycle averaged 5.20 and 6.19 days, 6.40 and 7.23 days, 7.30 and 7.85 days for male and female when fed on palm pollen, immature stages of T. urticae and B. tabaci, respectively. The highest R0 value of 17.88 expected female progeny per female was obtained with a diet of date palm and followed by 16.03 and 13.60 for T. urticae and B. tabaci, respectively. The intrinsic rate of natural increase (rm) and subsequently, the finite rate of increase (erm) averaged 0.232-1.26; 0.191-1.21 and 0.175-1.91 when E. scutalis was provided with date palm pollen, T. urticae and B. tabaci, respectively. Date palm pollen shortened the mean generation time and caused the highest rates of R0, rm and erm parameters followed by a diet of T. urticae. While B. tabaci proved to be the least favorite food source for E. scutalis.
PDF Abstract XML References Citation
Received: March 14, 2013;
Accepted: April 27, 2013;
Published: May 07, 2019
How to cite this article
A.H. Fouly, O.A. Nassar and M.A. Osman, 2013. Biology and Life Tables of Esieus scutalis (A.-H.) Reared on Different Kinds of Food. Journal of Entomology, 10: 199-206.
DOI: 10.3923/je.2013.199.206
URL: https://scialert.net/abstract/?doi=je.2013.199.206
DOI: 10.3923/je.2013.199.206
URL: https://scialert.net/abstract/?doi=je.2013.199.206
INTRODUCTION
Predacious mites of the family Phytoseiidae are cosmopolitan and important natural enemies of several phytophagous mite pests on various crops (Bond, 1989; Fouly and El-Laithy, 1992; Fouly, 1997; Gerson et al., 2003; Fouly et al., 2011; Mostafa, 2012). McMurtry and Croft (1997) characterized the phytoseiid mites according to their feeding habits into four major groups, I including spider mites specialists that do not feed on any other kinds of food. Group II including spider mites selective and group III of generalist that can feed prey mites or any other food. Group IV feed only on pollen.
Many species of phytoseiid are known to be obligatory predatory mites such as Phytoseiulus persimilis A.-H. and Glanderomus helveoulus (Chant) which are considered to be spider mites (Teranychid mites) predators (Caceres and Childers, 1991). Other species (especially in the genus Euseius) are facultative predators, which are not only mite predators but they also, have the ability to feed on other source of food such as white fly and pollen (Rasmy et al., 1987; Fouly and Hassan, 1991; Abou-Awad et al., 1992; Di Chiara and Tsolakis, 1995). This phenomenon increase species survival when natural animals preys are scared. These species are considered low density regulator, density independent (Abou-Setta and Childers, 1987, 1989; Fouly, 1997). Whereas, the major impact of "generalists", such as Euseius species, is at low population densities of spider mites, where they may prevent the widespread colonization and increase of the pest species (James, 1989; McMurtry, 1992) Also, field observation clearly showed that Euseius scutalis (A.-H.) is generally found in association with tetranychid mite species, scale insects, eggs and different immature stages of other insects as well as plant pollen (Yousef and El-Halawany, 1982; McMurtry et al., 1992; Al-Shammery, 2010, 2011).
In Egypt, E. scutalis is considered the most abundant and widely distributed phytoseiid mite (Zaher, 1986; El-Laithy and Fouly, 1992; Abou-Awad et al., 1998). It was noticed feeding on spider mites, Eriophyid and tenuipalpid mites, eggs and small insect species inhabiting the same habitat. Therefore, E. scutalis is considered one of the generalist that can feed and reproduce on a wide range of food source. The objective of this study improves our knowledge about biology and life table of E. scutalis when reared on different kind of food.
MATERIALS AND METHODS
Collection of mites: Phytoseiid species Euseius scutalis (Athias-Henriot) and spider mites Tetranychus urticae Koch were collected from leaves and twigs of castor plants Ricinus commuunis and hepiscus plants Hibiscus mutabilis L. (Malvaceae) growing at the farm of the Faculty of Agriculture, Mansoura University, Mansoura district.
Samples of plant leaves and twigs were collected in cellophane bags, with small pieces of cotton wool soaked in ether and brought to the laboratory for direct examination using a stereoscopic binocular microscope. Some adult specimens were individually mounted in Hoyer’s medium on glass slides for microscopic identification.
Copulated adult females were left to lay eggs on leaf discs of hibiscus plant kept on moist cotton pad in Petri-dish (15 cm in diameter) where suitable of moisture was daily supplied to keep leaf discs fresh for a longer time. Culture were provided with small pieces of cover slide glass and few cotton fibers as resting sides and collect deposited eggs easily.
Biological studies
Culture of Euseius scutalis: Predator cultures were divided into three major groups according to the type of food. The first group was provided with T. urticae, which previously collected from leaves of castor bean plant Ricinus communis L. as food source.
The second group of the predatory mite was fed on date palm pollen while the third one was maintained on immobile stage of white fly Bemisia tabaci which was collected from cabbage Brassica oleracea var. capitata L.
All groups were incubated at 26±1°C and 75%±3 R. H. where all culture were noticed daily to obtain the newly deposited eggs.
Rearing of Eseius scutalis: Leaf discs of Hibiscus mutabilis L. (Malvaceae) one square inch each was used for rearing E. scutalis during its whole life span.
Newly deposited eggs were singly transferred from the culture to the aforementioned leaf discs which were kept on a moist cotton pad in Petri dishes (15 cm in diameter). The incubation period was recorded and the newly hatched larvae were confined singly to the leaf discs and fed on the immatures of the spotted spider mites, white fly and date palm pollen.
In all cases, the newly emerged females were exposed to newly, ales from the same food group listed before.
Data were noticed for the whole life span, where observations were noted twice daily. Each rearing experiment was started with not less than 30 newly hatched larvae. Data were statically analyzed using Costat program (CoStat Sowftware, 1990).
Individual development, survival and egg production were observed where life table parameters were calculated according to Birch (1948), Laing (1968) and using the Basic Computer Program of Abou-Setta et al. (1986).
RESULTS AND DISCUSSION
Duration of developmental and adult stages: As in other phytoseiid mite species, both sexes of E. scutalis pass through an egg, larval, protonymphal and deutonymphal stages before reaching the adulthood. Under experimental conditions of 26°C and 70% RH, the incubation period ranged from 1.0 to 2.0 days. As shown in Table 1, larval stages durated 2.04, 2.66 and 2.61 days for female and duarted 2.0, 2.20 and 2.50 days for male when fed on palm pollen, individuals of T. urticae and B. tabaci, respectively. All larvae have been also observed feeding before molting to protonymphs. Similar results were obtained in the duration of protonymph and deutonymph stages. Therefore the protonymph and deutonymph stages durated 1.14, 1.33 and 1.57 days and durated 1.19, 1.47 and 1.66 days for female, while it durated 1.00, 1.40 and 1.60 days and durated 1.20, 1.50 and 1.60 days for male when fed on palm pollen, T. urticae and B. tabaci, respectively. Generally, it could be easily noticed that male emerged first and lived for shorter time than female. Similar, a shorter developmental time for male phytoseiids was previously reported by Elbadry and Elbenhawy (1968), Fouly (1982), El-Laithy and Fouly (1992), Momen and El-Saway (1993) and Momen (1995). However, there was no significant difference between the life cycle of both sexes of E. scutalis when mite individuals preys on the same kind of food as shown in Table 1.
Life cycle averaged 5.20 and 6.19 days; 6.40 and 7.23 days 7.30 and 7.85 says for male and female fed on pollen grains, T. urticae and B. tabaci, respectively. Therefore, it was obviously noticed that B. tabaci gave the longest life cycle followed by the two spotted spider mites while palm pollen gave the shortest developmental time. While, El-Laithy and Fouly (1992) found that life cycle of E. scutalis averaged 6.8 and 7.81 days for males and females fed nymphs of T. urticae. Also, Osman et al. (2010) showed that female life cycle of E. scutalis was 6.26 days when fed nymphs of T. urticae.
Duration adulthood, male lived for 21.50, 19.8 and 18.3 days while females lived for 24.14, 21.76 and 20.42 days after feeding palm pollen, T. urticae and B. tabaci, respectively. Therefore, it can be concluded that pllen grains prolonged E. scutalis adult longevity followed by immatures of T. urticae while B. tabaci, occupied the last rank. Therefore, kind of food significantly affected mite longevity Table 1.
As in most phytoseiid species, newly emerged females accepted copulation with one waiting males just after emergence. Several copulation are necessary to lay the maximum number of eggs. These results agree with findings of McMurtry and Scriven (1964), De Moreas and McMurtry (1981) and Marisa and Sauro (1990).
Before oviposition, an average female lives for 1.00, 2.04 and 1.57 days when E. scutalis female was provided with pollen grains, T. urticae and B. tabaci, respectively.
Postoviposition period lasted 8.19, 5.76 and 2.17 days as the previous food source offered, respectively Table 1.
| Table 1: | Duration (in days) of developmental and adult stages of E. scutalis (A.-H.) fed on different preys under laboratory conditions |
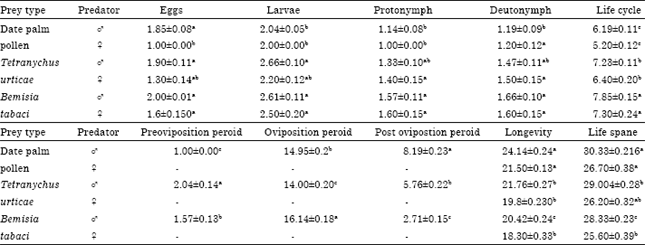 | |
| Means in each column followed by a different letter are significantly different, p<0.05 by Costat test. Values are Mean±SE, ♂: Male, ♀: Female | |
Female fecundity: It was clearly noticed that each female laid most of its eggs during the period locating in the first 10 days of oviposition period, hence it fed on greater amount of food. Data in Table 1 showed that adult female of E. scutalis started laying eggs after 1.00, 2.04 and 1.57 days when reared palm pollen, T. urticae and B. tabaci, respectively. Eggs production reached its peak after 10, 9 and 7 days when the same prey kinds were provided, respectively. During these periods (from start to peak), female mite laid an average of 59.6, 62.3 and 52.9% from the whole number of eggs. After that, it was obviously noticed that egg deposition gradually decreased and reached its lowest level after 17, 18 and 19 days after feeding on the previous diets, respectively.
Generally, females mite laid a total average number of 31.93, 28.63 and 24.30 eggs per female with a daily rate of 1.60, 1.45 and 1.30 eggs per female per day per female per day when it was provided with palm pollen, T. urticae and B. tabaci, respectively. Table 2. Similar results were obtained by Allawi (1991) who found that E. scutalis fed on castor bean, corn and pollen collected by honey bees laid a total number of 52.20, 33.50 and 40.53 eggs per female and a daily rate of 2.10, 1.43 and 1.98 eggs, respectively. Moreover, El-Laithy and Fouly, 1992 found that E. scutalis and A. swirskii fed on T. urticae laid 13.5 and 27.8 eggs per female, respectively. feeding on B. tabaci only, female mite of E. scutalis laid an average of 1.0 eggs per day (Fouly and Hassan, 1991).
Therefore, it can be concluded that the most important period for egg production not only for E. scutalis but also for most of phytroseiidae mites is within the first ten days. These observation agree with those obtained by Bounfour and McMurtry (1987), Abou-Setta and Childers (1989) and Fouly (1997) with regard to E. scutalis, E. mesembrinus and Propriosipsis asetus, respectively.
Life table studies: The percentage of daughter females in the total population of E. scutalis averaged 56, 54 and 54% when the food was palm pollen. T. urticae and B. tabaci, respectively. These values were considered later in the calculation of life table parameters.
Data in Table 2 shown that the mean generation time (T) of E. scutalis at 26°C and 70% R.H. was affected by diet. A diet of palm pollen provided the shortest T time (12.39 days), while the longest T time 14.91 days occurred when feeding on B. tabaci.
| Table 2: | Effect of diets on the life table parameters of Euseius scutalis 26°C and 70% R.H |
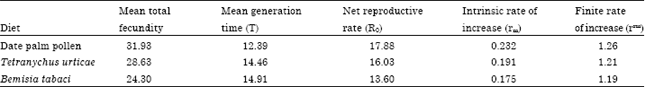 | |
Similar results were obtained by El-Laithy and Fouly (1992) who found that T time averaged 14.68 days for E. scutalis fed on nymphs of T. urticae under similar laboratory conditions. Mean total fecundity was significantly affected by diet. The highest female egg production of E. scutalis (31.93 eggs per female) was obtained with a diet of pollen while the lowest egg production was obtained when individuals were fed on B. tabaci (24.30 egg per female) as shown in Table 2. Similarly, Allawi (1991) found that the mean total fecundity of E. scutalis fed on corn pollen.
It was 33.50 eggs per female but it significantly increased to 52.20 eggs per female on castor bean pollen.
The net reproduction rate (R0 = ΣLxM x) which is a product of mean total fecundity, survival rate and sex ratio, followed the same pattern as mean total fecundity. The highest R0 value of 17.88 expected female progeny per female was obtained with a diet of date palm and followed by 16.03 and 13.60 for T. urticae and B. tabaci, respectively. This agree with the findings of El-Laithy and Fouly (1992) who found that R0 of E. scutalis and A. swirskii were 17.22 and 22.97 when provided with T. urticae.
On the other hand, Sabelis (1981) reviewed that the evaluation of the predator as biological control agent obviously depends on its reproductive capacity which positively correlated with prey consumption during its oviposition period. He found that prey consumption of most species formed a ratio of about 70% of the biomass o deposited eggs, in other words, the total eggs deposited during oviposition period represented about 70% from the total protein of consumed prey.
However, the intrinsic rate of natural increase (rm) is a key demographic parameter useful for predicting the population growth potential of an animal under given environmental conditions (Birch, 1948; Fouly, 1997; Ricklefs and Miller, 2000), because rm reflects an overall effect on development, reproduction and survival (Southwood and Henderson, 2000).
The intrinsic rate of natural increase (rm) and subsequently, the finite rate of increase (erm) averaged 0.232-1.26; 0.191-1.21 and 0.175-1.19 when E. scutalis was provided with date palm pollen, T. urticae and B. tabaci, respectively. Lower parameters were obtained by El-Laithy and Fouly (1992) who stated that rm values of E. scutalis fed T. urticae was 0.146 while erm was 1.157. the same authors found that these values were 0.167 and 1.180 for rm and erm when A. swirskii fed on the same prey, respectively. On the other hand, Osman et al. (2010) showed that the net rate of natural increase (rm) of E. scutalis was 0.229, whereas the finite rate of increase (erm) 1.25 when reared on nymphs of T. urticae. However, estimates of rm are difficult to compare between studies, because of genetic variation, differences in rearing methods and other environmental conditions and variable assumptions entering these estimations (Roy et al., 2003).
CONCLUSION
From the previous results, it can be concluded that pollen grains shortened the mean generation time and gave the best R0, rm and erm parameters followed by a diet of T. urticae. While fly B. tabaci proved to be the least suitable food source for the predatory mite E. scutalis. Generally, the mean total fecundity has a positive effect on rm values, whereas a longer mean generation time have a negative effect on rm values.
REFERENCES
- Abou-Awad, B.A., A.S. Reda and S.A. Elsawi, 1992. Effects of artificial and natural diets on the development and reproduction of two phytoseiid mites Amblyseius gossipi and Amblyseius swirskii (Acari: Phytoseiidae). Insect Sci. Appl., 13: 441-445.
CrossRef - Abou-Awad, B.A., A.A. El-Sherif, M.F. Hassan and G.M. Abou-Elela, 1998. Studies on development, longevity, fecundity and predation of Amblyseius olivi Nasr and Abou-Awad (Acari: Phytoseiidae) on various kinds of prey and diets. J. Plant Dis. Prot., 105: 538-544.
Direct Link - Abou-Setta, M.M. and C.C. Childers, 1989. Biology of Euseius mesembrinus (Acari: Phytoseiddae): Life tables and feeding behavior on tetranychid mites on citrus. Environ. Entomol., 18: 665-669.
Direct Link - Abou-Setta, M.M., R.W. Sorrell and C.C. Childers, 1986. Life 48: A basic computer program to calculate life table parameters for an insect or mite species. Florida Entomol., 69: 690-697.
CrossRefDirect Link - Allawi, T.F., 1991. Biological and ecological studies on indigenous and introduced phytoseiid mites: 1. Mass rearing and study of the effect of different pollen and other plant derivatives on the fecundity and longevity of Euseius scutalis (A.-H.). Dirasat, 18: 57-67.
Direct Link - Al-Shammery, K.A., 2010. Different biological aspects of the predaceous mite Euseius scutalis (Athis-Henroit) and the effects due to feeding on three tetranychid mites in Saudi Arabia. Asian J. Biol. Sci., 3: 77-84.
Direct Link - Al-Shammery, K.A., 2011. Plant pollen as an alternative food source for rearing Euseius scutalis (Acari: Phytoseiidae) in Hail, Saudi Arabia. J. Entomol., 8: 365-374.
CrossRefDirect Link - Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26.
CrossRefDirect Link - Bond, J., 1989. Biological studies including population growth parameters of the predatory mite Amblyseius barkeri [Acarina.: Phytoseiidae] at 25°C in the laboratory. Entomophaga, 34: 275-287.
CrossRef - Caceres, S. and C.G. Childers, 1991. Biology and life tables of Galendromus helveolus (Acari: Phytoseiidae) on Florioda citrus. Environ. Entomol., 20: 224-229.
Direct Link - Marisa, C. and S. Sauro, 1990. Biological observations and life table parameters of Amblyseius cucumeris (Oud.) (Acarina: Phytoseiidae) reared on different diets. Redia, 73: 569-583.
Direct Link - Di Chiara, S.R. and H. Tsolakis, 1995. The effect of Phenacoccus madeirensis green (Coccoidea: Pseudococcidae) on some biological parameters of four species of phytoseiid mites (Parasitiformes: Phytoseiidae). Isr. J. Entomol., 29: 301-307.
Direct Link - Elbadry, E.A. and E.M. Elbenhawy, 1968. The effects of pollen feeding on the predatory efficiency of Amblyseius gossipi (Acarina: Phytoseiidae). Entomologia Experimentalis et Applicata, 11: 273-277.
CrossRef - Fouly, A.H., 1997. Effects of prey mites and pollen on the biology and life tables of Proprioseiopsis asetus (Chant) (Acari, Phytoseiidae). J. Applied Entomol., 121: 435-439.
CrossRefDirect Link - Fouly, A.H. and A.Y.M. El-Laithy, 1992. Immature stages and life history of the predatory mite species Amblyseius barkeri (Hughes, 1948) (Acarina, Gamasida, Phytoseiidae). Deutsche Entomol. Zeitschrift, 39: 427-435.
CrossRefDirect Link - James, D.G., 1989. Influence of diet on development, survival and oviposition in an Australian phytoseiid, Amblyseius victoriensis (Acari:Phytoseiidae). Exp. Applied Acarol., 6: 1-10.
CrossRef - McMurtry, J.A., 1992. Dynamics and potential impact of generalist phytoseiids in agroecosysterns and possibilities for establishment of exotic species. Exp. Applied. Acarol., 14: 371-382.
CrossRef - McMurtry, J.A. and B.A. Croft, 1997. Life styles of phytoseiid mites and their roles in biological control. Ann. Rev. Entomol., 42: 291-321.
CrossRef - McMurtry, J.A., J.G. Morse and H.G. Johnson, 1992. Studies of the impact of Euseius species (Acari: Phytoseiidae) on citrus mites using predator exclusion and predator release experiments. Exp. Applied Acarol., 15: 233-248.
CrossRef - Mostafa, E.S.M., 2012. Laboratory studies on Euseius metwallyi a predator of the spider mite Tetranychus urticae on fruit trees in Egypt (Acarina: Phytoseiidae: Tetranychidae). J. Entomol., 9: 107-114.
CrossRefDirect Link - Osman, M.A., G.M. Abou-Elella and A.A. Tawfik, 2010. Role of four phytoseiid mite species and acarophagous ladybird, Stethorus gilvifrons (Mulsant) as bioagents of the two spotted spider mite Tetranychus urticae Koch. Acarines, 4: 47-55.
Direct Link - Rasmy, A.H., M.E. El-Bagoury and A.S. Reda, 1987. A new diet for reproduction of two predaceous mites Amblyseius gossipi and Agistemus exsertus (Acari: Phytoseiidae, Stigmaeidae). BioControl, 32: 277-280.
CrossRef - Roy, M., J. Brodeur and C. Cloutier, 2003. Effect of temperature on intrinsic rates of natural increase (rm) of a coccinellid and its spider mite prey. BioControl, 48: 57-72.
CrossRef - Fouly, A.H., M.A. Al-Deghairi and N.F. Abdel Baky, 2011. Biological aspects and life tables of Typhlodromips swirskii (Acari: Phytoseiidae) fed Bemisia tabaci (Hemiptera: Aleyroididae). J. Entomol., 8: 52-62.
CrossRefDirect Link