
Rahim Eslamizadeh
Department of Forest Management, Universiti Putra Malaysia, 34300 UPM, Serdang, Selangor, Malaysia
Ahmad Said Sajap
Department of Forest Management, Universiti Putra Malaysia, 34300 UPM, Serdang, Selangor, Malaysia
Dzolkhifli Omar
Department of Plant Protection, Universiti Putra Malaysia, 34300 UPM, Serdang, Selangor, Malaysia
Nur Azura Adam
Department of Plant Protection, Universiti Putra Malaysia, 34300 UPM, Serdang, Selangor, Malaysia
Journal of Entomology
Year: 2013 | Volume: 10 | Issue: 4 | Page No.: 182-190







ABSTRACT
Problems of efficacy and resistance manifested by whiteflies as consequences of chemical sprayings warrant a serious attention. Such hazardous outcomes call for an alternative method of controlling whiteflies that is safe to the produce, people and environment. In this study, efficacy of a newly isolated Isaria fumosorosea Wize infecting whitefly, Bemisia tabaci (Gennadius), from Universiti Putra Malaysia (UPM) was compared with isolates from bagworms, Pteroma pendula (Joannis) under laboratory conditions. All stages of B. tabaci were susceptible to infection by I. fumosorosea isolates when bioassayed at 1x106 mL-1. Among all the isolates tested, UPM isolate was the most virulent, causing 91, 90, 86 and 89% mortality on eggs and second, third and fourth instars, respectively. A concentration-mortality response test of the isolate on second instar nymphs resulted 95% mortality within 6 days when treated with 1x108 conidia mL-1. The results conclude that UPM isolate was the most effective against all stages of B. tabaci than those solates from other insects. Thus UPM isolate has the potential to be further developed as a biopesticide for controlling B. tabaci.
PDF Abstract XML References Citation
Received: May 07, 2013;
Accepted: June 12, 2013;
Published: November 06, 2013
How to cite this article
Rahim Eslamizadeh, Ahmad Said Sajap, Dzolkhifli Omar and Nur Azura Adam, 2013. First Record of Isaria fumosorosea Wize (Deuteromycotina: Hyphomycetes) Infecting Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Malaysia. Journal of Entomology, 10: 182-190.
DOI: 10.3923/je.2013.182.190
URL: https://scialert.net/abstract/?doi=je.2013.182.190
DOI: 10.3923/je.2013.182.190
URL: https://scialert.net/abstract/?doi=je.2013.182.190
INTRODUCTION
Bemisia tabaci (Gennadius) (Hemiptera:Aleyrodidae), commonly known as sweet potato whitefly, is an economically important whitefly worldwide (De Barro et al., 2011) having more than 600 host plants (Oliveira et al., 2001). Whiteflies cause indirect damages by transmitting plant disease causing viruses. The vectored viruses have been recorded causing over 40 diseases of vegetable and fiber crops (Faria and Wraight, 2001). Apart from vectoring serious plant viruses, its high genetic variability existing among in its populations (Brown et al., 1995) has created populations not only different in its feeding and reproduction behavior but also on its virus transmission characteristics (Burban et al., 1992; Bedford et al., 1994). Over the years the pest has been kept under control with the use of insecticides. Insecticide sprayings unfortunately have resulted resistance in B. tabaci populations apart from directly contaminating the produce and polluting the environment. Thus a sustainable and effective method is highly desirable for controlling the pest. Among the biologically effective agents for controlling insect pests are entomopathogenic fungi. Entomopathogenic fungi are microbes that infect, develop mycosis and ultimately killed the insect hosts by mechanical injuries or toxins produced and released by the fungi. More than 20 different species of fungi have been recorded infecting whiteflies throughout the world. Among them were Beauveria bassiana (Balsamo) Vuillemin, Lecanicillium lecanii (Zimmerm.), Isaria fumosorosea Wize (Ifr) (as Paecilomyces fumosoroseus) (Faria and Wraight, 2001). Zhu and Kim (2011) reported whitefly, B. tabaci biotype Q was susceptible to entomopathogenic fungi infection and I. fumosorosea was one of the effective fungi. In this study, efficacy of a newly isolated I. fumosorosea infecting B. tabaci was compared with isolates from bagworms, Pteroma pendula (Joannis) under a laboratory condition.
MATERIALS AND METHODS
Collecting and rearing B. tabaci: White fly, B. tabaci (biotype Asia 1) collected from eggplants, Solanum melongena, grown in greenhouses of Faculty of Agriculture, Universiti Putra Malaysia (UPM) were reared on eggplants in a greenhouse of Faculty of Forestry, UPM, at 65% ±5 RH and 25±1°C. Whitefly adults were introduced onto two-week old plants and were allowed to lay eggs for three days. Plants with eggs were transferred to new cages for subsequent development.
Selection of B. tabaci nymphal stages for bioassay: Different stages of whiteflies kept in different cages were used for the bioassay. Apart from the eggs, the instars were classified according to their sizes (L = length, W = width) as follows; second, (509±23.96 μm L, 303±18.95 μm W), third, (559±22.41 μm L, 352±17.75 μm W) and fourth, 613±14.67 μm L, 358±11.02 μm W). For the bioassays, the eggplant leaves with respective stages of whitefly were cut off and petioles wrapped with wet cotton were placed in 10 cm diameter Petri dishes.
Isolation I. fumosorosea from infected B. tabaci: Naturally infected adults and nymphs of B. tabaci were obtained from eggplants grown in greenhouses of the Faculty of Agriculture, Universiti Putra Malaysia (UPM). The samples were kept in 6-cm Petri dishes with a piece of moistened filter paper and maintained at the 25°C for 24-72 h to allow development of the fungi. Conidia from mycosed B. tabaci were isolated, plated onto Petri dishes containing Potato Dextrose Agar (PDA) and incubated at 25°C for 3 days. A single conidium isolation to obtain a pure fungal culture was conducted following a method described by Choi et al. (1999). The inoculated PDA in Petri dishes were incubated at 25°C for 3 days. Samples were examined and identified under a stereomicroscope (LEICA MZ6). Mycelia were mounted in lacto phenol/cotton blue (0.01% w/v) and observed under a phase contrast microscope (Nikon ECLIPSE E200). Photographs were taken using a Nikon camera fitted on the microscope. Length and width of fungal structures (conidia, conidiophores, phialides and hyphae) from fresh infected cadavers were measured to enable specific identification. They were identified according to taxonomic keys and monographs of Samson (1975). Portions of the pure culture were maintained on agar slants at 4°C.
Preparation of specimen for scanning electron microscopy (SEM): Infected second nymphs were placed into separate vials and fixed in 4% glutaraldehyde for 12 h, washed in three changes of 0.1 M sodium cacodylate buffer for 10 min each, fixed in osmium tetroxide for 2 h and rinsed in three changes of 0.1 M sodium cacodylate buffer. The specimens were dehydrated in a series of acetone (35, 50, 75 95 and 100%). The dehydrated specimens were critical point dried for about 30 min, mounted onto specimen stubs, sputter coated and viewed under a Philips 400 Scanning Electron Microscope (SEM) at Bioscience (IBS), UPM.
Propagation of isolates: One newly isolated I. fumosorosea and nine I. fumosorosea previously isolated from bagworms P. pendula were screened against B. tabaci (Table 1). The isolates were repropagated from a single conidium to obtain pure cultures. To achieve this, 100 μL of conidial suspensions (1x106 conidia mL-1) was plated on PDA medium with 1% yeast extract and incubated at 25°C for 3 days. Following incubation, a single colony was transferred to another Petri dish with PDAY medium and incubated at 25°C for one week. Conidia were harvested by washing the dishes with an aqueous solution of 10 mL 0.1% Tween 80. Subsequently, conidia suspension was filtered through a filter paper to remove mycelia and then shaken for 5 min in a vortex mixer to homogenize the suspension. Conidia were counted in an improved Neuberger hemocytometer and aliquots were adjusted to the desired concentration (Sevimab et al., 2009).
Production of isolates: Each isolate was produced on cooked rice. Autoclavable polypropylene plastic bags (18x25 cm), filled with 250 g of fungal- inoculated rice, was incubated for 14 days at 25±1°C for fungal development and sporulation. The conidiated rice grains were spread out in clean plastic trays and dried in a drying cabinet at 25±3°C. After five days, when the moisture content was about 20%, conidia were extracted from the rice by sieving through a brass laboratory sieve (300 mm mesh). After sieving, conidia were dried further in an auto-desiccator with a built-in hygrometer for four days to reduce the moisture content to around 5%.
Bioassay: The viability of conidia of each isolate was determined by placing a drop of each diluted conidial suspension on PDAY. The PDAY was covered with a sterile microscope cover slip and percent germination was assessed after 24 h of incubation at 24±2°C. Conidia were considered to have germinated if the germ tube was longer than the diameters of the conidia (Yeo et al., 2003). The number of germinated and ungerminated conidia was counted and percent viability was calculated. Isolates with viability more than 95% were used in the bioassay. The 1x106 mL-1 conidia suspension was prepared for all isolates. The control was treated with 0.1% Tween 80 in sterile water (Burges and Thompson, 1971). Eggplant plant leaves with at least 30 the each stage of the eggs, second, third and fourth instars nymph of B. tabaci, were excised and the petiole wrapped with moistened absorbent cotton wool. Eggs and nymphs were counted and they were dipped in the conidia suspension (1x106 mL-1) for 2-3 seconds or in the control solution. The leaves were air dried for 5 min and returned separately into Petri dishes lined with a piece of moistened filter paper. Mortality was recorded daily and only those whiteflies that were showing symptoms of fungal infection as manifested by sporulation of the fungus on their cuticle were counted as a kill by the pathogen (Cuthbertson et al., 2005). The experiment was replicated four times.
| Table 1: | List of I. fumosorosea isolates showing their insect hosts and geographic location |
 | |
Concentration-mortality response of UPM isolate: The most effective isolate was chosen and a stock concentration of 5x108 conidia mL-1 as determined using a Neubauer hemocytometer, was prepared. The suspension was serially diluted in distilled water to 1x108, 1x107, 1x106, 1x105 and 1x104 mL-1 conidia. Second instar nymphs were used in the bioassay. Leaves with at least 30-second instar nymphs were excised from the plant. Each leaf was dipped in a different freshly prepared conidia suspension 1x106 mL-1 for 2-3 seconds or in 0.1 Tween 80 aqueous solutions for controls. The experiment was replicated four times and mortality was recorded daily.
Data analysis: All percentage of mortality data were corrected using Abbott’s formula. The LT50 and LC50 values were calculated by Probit Analysis (statistical package Polo Plus version1), based on Finney (1952). An Analysis of Variance (ANOVA) was performed on the data and the treatment means were separated by Tukey’s multiple range tests at 0.05% level of significance.
RESULTS
Symptom of infection on B. tabaci: This paper reports a new record of natural infection of I. fumosorosea on B. tabaci in Malaysia. Figure 1a and b, respectively, show infected nymphs and adults with the mycelia covering their body. A microscopic observation shows that the conidia germinated on the insect cuticle geminated after one day of infection when incubated at 25±1°C. and 65%±5 RH. The mycelia grew and colonized the surface of cuticle between second and third day and then they penetrated through and reached the hemocoel between third and fourth day. The hyphae then ramified within the host body cavity (Fig. 2). Infected insect usually died after six or seven days after infection and new propagules formed on or in the insect.
Growth on potato dextrose agar (PDA): Cultures grew moderately fast on PDA, producing of a basal felt with raised floccose overgrowth, homogeneous diameter and regular borders, reverse initially colourless but later becoming pink (Fig. 1-3). Hyphae are flat walled, hyaline, 2.5±0.2 μm wide (n = 20). Conidiophores, 2.4±0.2 μm wide (n = 20), are smooth walled, colourless, consisting of verticallate branches bearing whorls of 4-6 phialides. Phialides with a globose basal portion tapering into a long distal neck of 6.7±0.5x1.9±0.2 μm (n = 20). Conidia are hyaline to slightly pink, cylindrical to fusiform measuring 4.8±0.5x1.1±0.3 μm (n = 50). Figure 4 shows whorl of phialides bearing conidia (a) and conidial mass (b).
 | |
| Fig. 1(a-b): | (a) Nymphs and (b) adults of B. tabaci infected with I. fumosorosea. Note mycelia covering their body |
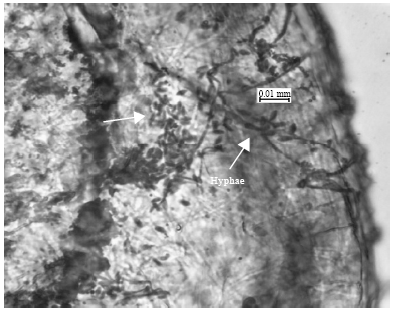 | |
| Fig. 2: | Fungal hyphae ramifying inside the body of B. tabacci nymph |
 | |
| Fig. 3: | Purified I. fumosorosea culture in a Petri dish |
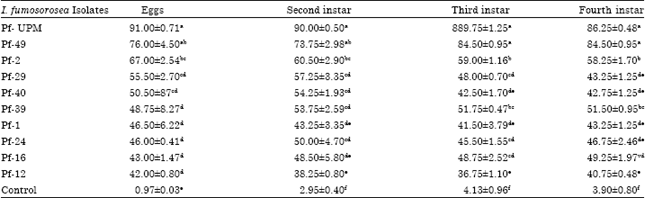
Bioassay on B. tabaci: Conidia of each isolate used in the bioassay had more than 95% germination. All isolates were infective towards eggs, second, third and fourth instars of B. tabaci but with varying degrees of infectivity. Highest mortality was recorded from B. tabaci that had been treated with the newly isolated I. fumosorosea. The isolate caused 91% infection on the eggs and mortalities ranging from 84 on the fourth to 91.2% on the second instars. The next comparable potent isolate was Pf49 which caused mortality of more than 70% of all treated stages of whiteflies. It caused 76.00% mortality on the eggs and 73.75 to 84.50% on the nymphs. The isolates Pf 39, Pf40, Pf 29 and Pf 2 caused mortalities ranging from 48.75 to 67.00% on the eggs, 53.75 to 60.50% on the second instar, 51.75 to 59.00% on the third and fourth instars.
 | |
| Fig. 4: | Phialides I. fumosorosea with (a) conidia and (b) conidial mass |
| Table 2: | Means comparison of mortality of eggs and second, third, fourth in stars of B. tabaci treated with 1x106 conidia mL-1 of I. fumosorosea isolates |
 | |
| Value are presented and mean±SD. Means followed by the same letter within the same column are not significantly different using Turkey’s multiple range test at 0.05 level of significance | |
The remaining isolates, Pf12, Pf16, Pf 24 and Pf1, caused 42.00 to 46.50% mortalities on the eggs and 36.75 to 51.50% on the second, third and fourth instars (Table 2). The least infective isolate was Pf-12 where only 42.80% of the eggs and less than 40% of the nymphs were infected.
Concentration-mortality response of B. tabaci treated with UPM isolate: The concentration-mortality response tests on the second instar nymphs shows that there was a linear relation between dose and mortality with a goodness of fit test (χ2 = 12.27, df = 12, p<0.05) indicates low data deviation and t value of the slope (t = 8.69). The percentage mortality significantly increased with the increase in the conidial concentration treated on the nymphs. After seven days of inoculation 100% cumulative mortality was achieved when the nymphs were treated with 1x108 conidia mL-1 . Concentrations 1x105 to 1x107 conidia mL-1 caused mortalities ranging from 55 to 95% and there were significant differences between concentrations (p<0.05) (Fig. 5). The lowest cumulative mortality, 50%, was recorded from nymphs that had been treated with 1x104 mL-1 conidia. Mortality in the control whiteflies was 1%. The LC50 value of this fungus against the second instars six days after treatment was 5.577x105 conidia mL-1 (Table 3). The LT50 values calculated following immersion of second instars of B. tabaci in aqueous suspensions of I. fumosorosea 1x104, 1x105, 1x106 , 1x107, and 1x108 conidia mL-1 are shown in Table 4.
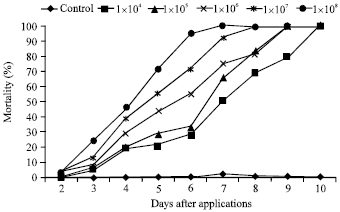 | |
| Fig. 5: | Cumulative mortality of B. tabaci second instar nymph after application of fungal Isaria fumosorosea UPM isolate at different doses of conidia |
| Table 3: | Lethal Concentrations (LC) values of I. fumosorosea UPM isolate against B. tabaci second instar nymph |
 | |
| Table 4: | Median lethal time (LT50) values of B. tabaci second instar nymph treated with different conidia concentrations of I. fumosorosea UPM isolate |
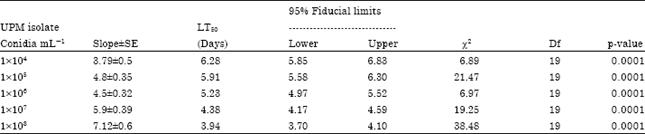 | |
The data indicated that the LT50 values decreased with the increase in the conidial concentration. The LT50 for 1x104 was 6.38 days and the value shortened by one day to an average of 5.91 days when the concentration was increased ten folds to 1x105 mL-1. The LT50 values subsequently shortened to 5.23, 4.38 and 3.94 days when the whiteflies were treated with concentrations of 1x106, 1x107 and 1x108 mL-1, respectively.
DISCUSSION
Entomopathogenic fungi are an important option in Integrated Pest Management (IPM) programs for controlling sap-sucking insects such as whiteflies (eggs and nymphs) and aphids (Lacey et al., 1996). One the entomopathogenic fungi that can be considered for sap sucking insects was I. fumosorosea. The I. fumosorosea UPM isolate obtained from naturally infected B. tabaci in a greenhouse of Universiti Putra Malaysia, a new record, was very effective against its host. The fungus grew relatively fast, produced an abundant conidia and it was more effective than those I. fumosorosea isolated from P. pendula. As expected variations in their virulence to the target pest occurred among the isolates. In similar studies elsewhere, various isolates of L. lecanii and I. fumosorosea from aphids, whiteflies and other insects showed distinct differences in their ability to infect whiteflies. Isolates collected from whitefly, Trialeurodes vaporariorum, were usually more effective against eggs and nymphs of whiteflies than those isolated from other insects including aphids (Hall, 1982; Kitazawa et al., 1984; Masuda and Kikuchi, 1992) and strains that originate from whiteflies caused higher mortalities on their eggs and nymphs (Hall, 1982; Chandlaer et al., 1993). Mesquita et al. (1996), however, reported that I. fumosorosea isolated from B. tabaci was also effective against Russian wheat aphid, Diuraphis noxia. Genetic variability in I. fumosoroseus could be one of the reasons for the differences in their virulence against the whitefly (Tigano-Milani et al., 1995) and these variations also occurred in other entomopathogenic fungi infecting their hosts. For example, varied virulence occurred in isolates of Beauveria bassiana and Metarhizium anisopliae and M. anisopliae var. acridum on house fly, Musca domestica Anderson et al. (2011). Concluded that the variations in virulence was attributed to the quantitative speed of the fungal growth. Thus in an effort to select a suitable isolate to be used in a pest management program, a wide range of isolates need to be screened against its target pest because an infective fungal isolate is a prerequisite for a successful pest management.
CONCLUSION
Results from this study proved that I. fumosorosea of UPM isolate was effective against B. tabaci. Thus isolate has the potential to be used as one of the microbial control agents for managing B. tabaci. Subsequent studies on its production, delivery system and persistence in the pest environment need to be carried out.
ACKNOWLEDGMENT
This research was partly supported by Research University Grant Scheme (RUGS) of Universiti Putra Malaysia, Serdang, Malaysia.
REFERENCES
- Anderson, R.D., A.S. Bell, S. Blanford, K.P. Paaijmans and M.B. Thomas, 2011. Comparative growth kinetics and virulence of four different isolates of entomopathogenic fungi in the house fly (Musca domestica L.). J. Invertebr. Pathol., 107: 179-184.
CrossRefPubMedDirect Link - Bedford, I.D., R.W. Briddon, J.K. Brown, R.C. Rosell and P.G. Markham, 1994. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Applied Biol., 125: 311-325.
CrossRefDirect Link - Brown, J.K., D.R. Frohlich and R.C. Rosell, 1995. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol., 40: 511-534.
CrossRefDirect Link - Burban, C., L.D.C. Fishpool, C. Fauquet, D. Fargette and J.C. Thouvenel, 1992. Host associated biotypes within West African populations of the whitefly Bemisia tabaci (Genn.) (Homoptera, Aleyrodidae). J. Applied Entomol., 113: 416-423.
Direct Link - Chandlaer, D., J.B. Heala and T. Gillespie, 1993. Germination of the Entomopathogenic Fungus Verticillium lecanii on Scales of the Glasshouse Whitefly Trialeurodes vaporariorum. Biocontrol. Sci. Technol., 3: 161-164.
CrossRef - Choi, Y.W., K.D. Hyde and W.W.H. Ho, 1999. Single spore isolation of fungi. Fungal Divers., 3: 29-38.
Direct Link - Cuthbertson, A.G.S., K.F.A. Walters and C. Deppe, 2005. Compatibility of the entomopathogenic fungus Lecanicillium muscarium and insecticides for eradication of sweet potato whitefly, Bemisia tabaci. Mycopathologia, 160: 35-41.
Direct Link - De Barro, P.J., S.S. Liu, L.M. Boykin and A.B. Dinsdale, 2011. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol., 56: 1-19.
CrossRefPubMedDirect Link - Faria, M. and S.P. Wraight, 2001. Biological control of Bemisia tabaci with fungi. Crop Prot., 20: 767-778.
CrossRefDirect Link - Hall, R.A., 1982. Control of whitefly, Trialeurodes vaporariorum and cotton aphid, Aphis gossypii in glasshouses by two isolates of the fungus, Verticillium lecanii. Ann. Applied Biol., 101: 1-11.
CrossRefDirect Link - Lacey, L.A., J.J. Fransen and R. Carruthers, 1996. Global Distribution of Naturally Occurring Fungi of Bemisia, their Biologies and Use as Biological Control Agents. In: Bemisia 1995: Taxonomy, Biology, Damage and Management, Gerling, D. and R. Mayer (Eds.). Intercept Andover Publishers, UK., pp: 401-433.
- Mesquita, A., L.A. Lacey, G. Mercadier and F. Leclant, 1996. Entomopathogenic activity of a whitefly derived isolate of Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes) against the Russian wheat aphid, Diuraphis noxia (Homoptera: Aphididae) with the description of an effective bioassay method. Eur. J. Entomol., 93: 69-75.
- Oliveira, M.R.V., T.J. Henneberry and P. Anderson, 2001. History, current status and collaborative research projects for Bemisia tabaci. Crop Prot., 20: 709-723.
CrossRefDirect Link - Sevimab, A., I. Demira, E. Tanyelia and Z. Demirbaga, 2009. Screening of entomopathogenic fungi against the European spruce bark beetle, Dendroctonus micans (Coleoptera: Scolytidae). Biocontrol. Sci. Technol., 20: 3-11.
CrossRefDirect Link - Tigano-Milani, M.S., R.J. Honeycutt, L.A. Lacey, R. Assis, M. McClelland and B.W.S. Sobral, 1995. Genetic Variability of Paecilomyces fumosoroseus isolates revealed by molecular markers. J. Invertebr. Pathol., 65: 274-282.
CrossRefPubMedDirect Link - Yeo, H., J.K. Pell, P.G. Alderson, S.J. Clark and B.J. Pye, 2003. Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and their pathogenicity to two aphis species. Pest Manage. Sci., 59: 156-165.
CrossRefPubMedDirect Link - Zhu, H. and J.J. Kim, 2011. Susceptibility of the tobacco whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) biotype Q to entomopathogenic fungi. Biocontrol. Sci. Technol., 21: 1471-1483.
CrossRef