
N.N. Ntonifor
Department of Biochemistry and Microbiology, University of Buea, P.O. Box 63 Buea, Cameroon
M.C. Parr
Department of Plant and Animal Sciences, University of Buea, P.O. Box 63, Buea, Cameroon
J.A. Ewunkem
Department of Natural Resources and Environmental Design, College of Agriculture and Environmental Sciences, North Carolina A and T University, Greensboro, North Carolina, USA
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 2 | Page No.: 79-88







ABSTRACT
This study was conducted to elucidate the frequent occurrences of the tropical vagrant huntsman spider, Heteropoda venatoria in bananas, for which, the spider habitats, temporal abundance, distribution and dynamics were studied in banana agro-ecosystems in Cameroon. Seasonal observations of banana plantations and associated management practices revealed that banana agro-ecosystems and host plant architecture offered diverse suitable habitats for H. venatoria. The spider inhabited the soil litter spots/mulches, loose leaf sheaths (barks) of pseudostems and stumps, leaf petioles, spaces between banana flower bracts and clusters and those between fingers of bunches. Spider densities in these habitats varied with plant phenology and seasons. The spiders inhabited the loose leaf sheaths around pseudostems and their stumps, the stout tube-like leaf petioles and leaf sheaths of pre-flowering plants during dry spells or seasons. During the rains or when farms were under irrigation, the relatively more sheltered inner parts of flower bracts and spaces between the fruit clusters and those between the fingers of bunch were preferred. Higher densities of H. venatoria were recorded in pseudostems and stumps around refuse areas with distinct population gradients in the order of 0-10>10-20>20-30>30-40 m from the refuse. Spider densities in pseudostems and litter spots were lower during the rainy than the dry season. In contrast, highest and lowest spider densities in bunches were recorded during the rainy and dry seasons, respectively. This ecological information is vital in the sustainable management of the spider populations so as to advantageously exploit their beneficial predatory effects in banana agro-ecosystems.
PDF Abstract XML References Citation
Received: July 14, 2011;
Accepted: September 27, 2011;
Published: December 12, 2011
How to cite this article
N.N. Ntonifor, M.C. Parr and J.A. Ewunkem, 2012. Seasonal Abundance and Distribution of the Huntsman Spider, Heteropoda venatoria (Sparassidae: Araneae) in Banana Agro-ecosystems in Cameroon. Journal of Entomology, 9: 79-88.
DOI: 10.3923/je.2012.79.88
URL: https://scialert.net/abstract/?doi=je.2012.79.88
DOI: 10.3923/je.2012.79.88
URL: https://scialert.net/abstract/?doi=je.2012.79.88
INTRODUCTION
Bananas are an important food and cash crop in most developing tropical countries (Tripathi et al., 2009). Bananas and plantains (Musa spp.) provide more than 25% of the carbohydrate intake for over 70 million people and also serve as important sources of revenue for smallholder farmers in sub-Saharan Africa (Oselebe and Tenkouano, 2009). The fruits are highly nutritious and contain large amounts of carbohydrates and minerals like phosphorus, calcium and potassium as well as vitamins A and C (Adeniji and Empere, 2001; Adeniji et al., 2006). This food crop is rated as the fourth most valuable food after rice, wheat and milk (Ploetz, 2001). Bananas are also a major fruit in international trade and as such are often grown in commercial plantation systems where they are intensively managed (Karamura and Gold, 2000).
Spiders are amongst the most dominant and diverse arthropods on vegetation (Morais-Filho and Romero, 2008) and are thus important components of ecological webs in agroecosystems (Thorbek et al., 2004). Environmental characteristics such as host plant architecture as well as prey availability strongly influence the abundance and distribution of spiders on plants (Romero and Vasconcellos-Neto, 2005). Some spiders like the green lynx spider Peucetia viridans prefers plants that offer better sources of food, shelter and other favourable environmental conditions (Arango et al., 2000). Given that spiders are generalist predator species, they are useful in agriculture since they help in controlling pest insect and mite populations in fruit, vegetable and field crops (Symondson et al., 2002). Therefore, understanding the ecology of spiders in agroecosystems is important given the vital predatory role that these arachnids play in suppressing plant pests (Harwood and Obrycki, 2007).
In recent years, spiders especially the giant crab spiders or huntsman spiders, Heteropoda venatoria (Linnaeus) (Sparassidae: Araneae) have occasionally appeared in marketed bananas (Edward, 2009). The flattened body and long flexible legs of this relatively large cosmotropical spider species enables it to fit surprisingly into small cracks and crevices (Edward, 2009). H. venatoria is highly valued in tropical countries because they capture and feed on cockroaches, other domestic insects (Shukla and Lele, 2008) and field insect pests of crops. Despite the innocuous nature of H. venatoria to humans, its frequent occurrences in high numbers in human habitations and especially in marketed bananas calls for concern. The presence of this spider in marketed bananas scares consumers of the commodity and this can affect market prices of this highly valuable fruit and hence can lead to financial losses to the banana industry. This underscores why banana producers, consumers and associated stakeholders need explanations for the recurrent occurrences of H. venatoria in marketed bananas and how the situation can be mitigated in banana agro-ecosystems. Therefore, the objectives of this study were to identify; (1) the various habitats of H. venatoria in banana plantations, (2) study the spider abundance, temporal distribution and dynamics on banana plants and also (3) identify cultural practices that can be used to minimize the spider densities especially in banana bunches.
MATERIALS AND METHODS
Study site: The study was carried in Tiko situated at 04°04’ 22″ N and 09°19’ 40″ E in the South Western region of Cameroon in the Gulf of Guinea in Africa. The site has sandy loam soils, an annual rainfall of about 2000 mm and two main seasons; the dry which runs from mid-November to mid-March and the rainy season from mid-March to mid-November. Two types of banana and/or plantain cropping systems were practiced in this area; the subsistence system and the commercial plantation system (Karamura and Gold, 2000; Okolle et al., 2006). The subsistence system often consists of a few mats of rain-fed plantains and/or bananas often mulched with kitchen refuse and garbage; products from this are used as food and/or as a source of revenue. The plantation system consists of one cultivar of desert banana planted in mats of about 2 m between and within rows and intensively managed often with irrigation and fertilized with mineral fertilizers. Products from these commercial plantations are predominantly for exportation. The commercial plantation used in the study had a surface area of 479.9 hectares with a total of 988,815 plants.
Identification of spiders and their habitats in banana farms: Spiders collected from the banana farms were taken to the Department of Plant and Animal Sciences, University of Buea, Cameroon for identification. All identifications were done using taxonomic keys (Platnick and Levi, 1973) and pictorial guides (Edwards, 1979) and by comparing with voucher specimens.
To identify the various habitats of huntsman spiders in banana farms, two commercial banana farms and their surrounding subsistence banana and/or plantain farms were selected and sampled weekly for two months. Each week, 100 randomly selected banana mats from each commercial farm and ca 10% of those in any surrounding subsistence farm were sampled. A mat in the commercial farms comprised of a parent banana plant with two to three follower younger plants all attached to a common corm while in a subsistence farm there could be up to six followers per mat. On each sampling date, all the mulch within a two meter circumference of each mat was carefully lifted with a wooden rod and checked for the presence of H. venatoria spiders. Thereafter, inflorescences and bunches when present, all leaf surfaces, petioles, loose barks of every pseudostem or stump of each mat were systematically checked to record all the sites inhabited by H. venatoria. All observations were done between 08.00-10.00 h when the spiders were resting after the night hunt.
Abundance and seasonal distribution of H. venatoria in banana farms: Based on the results of the identification of the habitats of H. venatoria above, 100 pseudostems (50 bunched and 50 pre-flowering plants) were randomly selected from each of two commercial farms and sampled weekly as described above. On each visit, all H. venatoria present in each habitat were counted and especially after rainfall in order to also determine the effect of rainfall on the vertical distribution of the spiders on banana plants. For more detailed sampling, each banana pseudostem was stratified into 3 strata on the basis of heights from the ground: stratum 1: ≤1 m from the base of the plant; stratum 2: >1≤2.5 m; stratum 3: >2.5 to the leaves and bunches. Once weekly from April 2006 to March 2007, all loose leaf sheaths on the pseudostems were gently and carefully opened, observed then the number and positions of huntsman spiders present in each stratum recorded on a scouting sheet. An adjustable ladder was used to sample the inflorescences and bunches. In addition, on each sampling date, 50 upright pseudostem stumps attached to the mat in situ and also 50 litter spots (mulches) were also randomly sampled from each farm. Rainfall data for the study area and period were obtained from the Meteorological Station of the Cameroon Development Cooperation in Tiko, Cameroon.
Given that preliminary observations showed a high association of H. venatoria with refuse dumps in the banana farms, these refuse sites were also selected for detailed sampling. All banana pseudostem residues after harvest lying horizontally on the ground within 50 m around refuse areas were stratified as follows: 0-10, >10-20, >20-30 and >30-50 m away from the refuse dump. Within each of these strata, 50 pseudostem residues and other trash were sampled once weekly from April 2006 to March 2007, by using a wooden rod to carefully lift the refuse to record the number of spiders present in each stratum.
Effect of farm sanitization on H. venatoria densities: To determine the effects of farm sanitization on H. venatoria densities, a one hectare area of the farm with high densities was selected for the study. Each month from June to December 2007, baseline data of spider population densities on 50 pseudostems were taken. Then 25 out of these pseudostems were debarked and all dry leaves on the plants removed and used as mulches. Subsequently, once each week, the number of spiders on each of 25 sanitized and non sanitized pseudostems were counted and recorded.
Thereafter, monthly densities of the spiders per 25 sanitized and non sanitized pseudostems each were calculated and compared using a student t-test.
Statistical analysis: Data for the distribution of spiders around refuse dumps were analyzed by Analysis of Variance (ANOVA) using the Statistical Package for Social Scientists (SPSS). Differences between means were assessed using the LSD test at p = 0.05. Monthly spider densities in the sanitized and non-sanitized pseudostems were compared using a student t-test (Gomez and Gomez, 1984).
RESULTS
Habitats of H. venatoria in banana farms: Regardless of whether it was a commercial or subsistence farm, both the adults and juveniles of H. venatoria were highly prevalent in the banana farms. Given the large and flattened body nature of H. venatoria that enables this large spider to fit into small cracks and crevices, the banana plant architecture therefore offered a diversity of habitats for the spider to exploit. At harvest, the banana pseudostems were cut off to leave stumps of various heights which were also inhabited by the spiders. The habitats and hence the distribution of the spiders on the plants varied with the banana plant phenology (Table 1). On the pre-flowering banana plants, there were three habitats namely; soil litter/mulches, the loose (partially opened) leaf sheaths around the pseudostems and the stout leaf petioles or stalks which extend to the tube-like leaf sheaths often inhabited by the relatively smaller and more active juveniles. These habitats mentioned above were often exploited by the spiders during the dry seasons or dry spells of the rainy season. During the shooting (flowering) stage of the plant, the inside parts of the purplish-to-greenish fleshy bracts and beneath the placental leaf of the inflorescence were also inhabited by the spiders. The bunched banana plant offered two additional habitats; when the flower emerged, the spaces between the clusters of the fruits (hands) and those between the banana fingers on the bunches were also inhabited by H. venatoria. If the bunches were eventually protected with polythene tree bags, the spiders also hided inside the bags as an additional (man-made) habitat. These relatively more shady habitats were often preferred during rainy periods or when the farms were under irrigation. The mulches of pruned leaves, chopped pseudostem portions and assorted litter around the bases of the banana mats also served as supplementary habitats for the spiders. However, <5% of the spiders were found in the litter spots in the farms which in contrast harboured many toads, frogs, agama lizards and geckoes which preyed on the spiders.
Abundance and temporal distribution of H. venatoria in the habitats: During the day, the vagrant H. venatoria spiders were less active and often stayed concealed in the various habitats in the banana agro-ecosystem. At all times and regardless of the type of farm, only one adult spider was found under each pseudostem bark.
| Table 1: | Various habitats occupied by Heteropoda venatoria spiders in a banana farm |
 | |
Generally, most of the adults often perched facing downwards inside the loose leaf sheaths (barks) of the pseudostems and pseudostem stumps at heights of ≤2 m from the bases. The vertical distribution of the spiders in the different strata of the pseudostem is shown in Table 2. At the peaks of the rainy season (June to September), when excessive rain water run off collected and stagnated at the bottoms of the leaf sheaths, more of the spiders were found perched in the second stratum (>1<2 m) from the bases of the pseudostems as well as inside the relatively more sheltered banana bunches. However, from the onset of the dry season (October) through to the early months of the rainy season (April and May), more of the spiders descended further down into the first stratum at ≤1 m from the bases of the pseudostems. There was hardly more than one adult spider found under each loose leaf sheaths of the pseudostems. Occasionally, during the dry season, a few juvenile spiders resided inside the same stout leaf stalk often located at heights ≥2.5 m.
Seasonal densities of H. venatoria in various habitats: The densities of spiders in the pseudostems were lower during the rainy season (May-October, 2006) compared to the dry season, with the lowest value of 8 spiders per 50 pseudostems obtained in August 2006 (Fig. 1). At the onset of the dry season around ending of October 2006, the spider densities started increasing, to attain a peak of 21 spiders per 50 pseudostems in February 2007. Therefore, the trends of the spider population variations followed the monomodal rainfall pattern of the study area. The trends of the spider densities in the bunches were a direct opposite of those for the pseudostems and stump. In the bunches, the population started building up at the onset of the rains (April) to a peak in September, then started dropping from the onset of the dry season (October) to a trough in February. By and large, throughout the dry season, the number of spiders/50 bunches was relatively lower compared to the rainy season (Fig. 2). The population trends of the litter spots or mulches were similar to those of the pseudostems with higher densities during the dry seasons compared to the rainy season when the population crashed to zero from July to November (Fig. 3).
| Table 2: | Monthly vertical distribution (Mean No.±SE) of Heteropoda venatoria spiders per 100 combined pre-flowered and bunched banana plant pseudostems (2006-2007) |
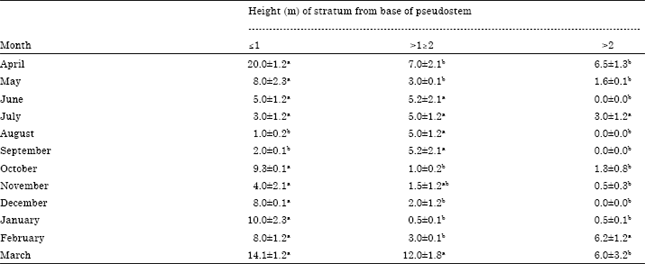 | |
| Stratum 1: ≤1 m from base of the plant; stratum 2: >1≤2 m from the base while, stratum 3: >2 m from base to all the leaves. Means followed by the same letter in a row are not significantly different (p>0.05) | |
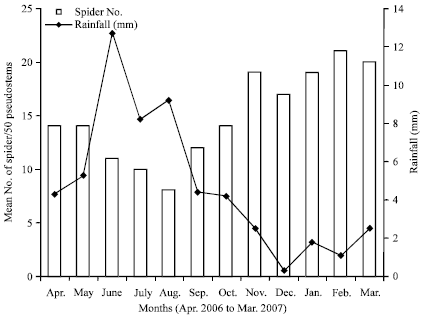 | |
| Fig. 1: | Seasonal variations in mean number of Heteropoda venatoria spiders per 50 banana pseudostems and rainfull in Tiko, Cameroon (2006-2007) |
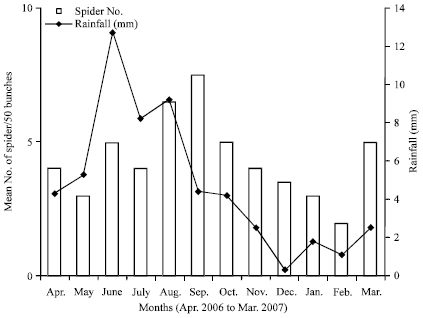 | |
| Fig. 2: | Seasonal variations in mean number of Heteropoda venatoria spiders per 50 banana bunches and rainfall in Tiko, Cameroon (2006-2007) |
Distribution of H. venatoria around refuse dumps: Overall, there were higher densities of the huntsman spiders in the barks of banana pseudostems and pseudostem stumps around refuse dumps in the banana farms. Generally, spiders in these areas were relatively larger. Within these vicinities, a spider population concentration gradient was observed with the highest concentration recorded within 0-10 m away from the refuse dump. By and large, the H. venatoria population gradient was in the order 0-10>10-20>20-30>30-40 m away from the refuse areas (Table 3). There were significant (p<0.01) differences in the populations within these intervals.
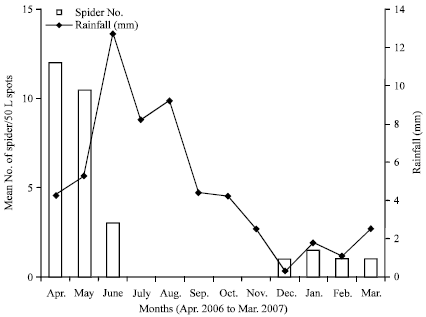 | |
| Fig. 3: | Seasonal variations in mean number of Heteropoda venatoria spiders per 50 litter spots and rainfall in Tiko, Cameroon (2006-2007) |
| Table 3: | Monthly mean number (±SD) of Heteropoda venatoria spiders per 50 banana pseudostem stumps located at varied distances (m) from a refuse dump in a banana farm, Tiko, Cameroon (2006-2007) |
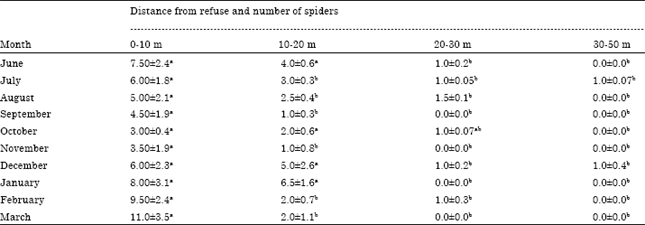 | |
| Means followed by the same letter in a row are not significantly different (p>0.05) | |
Similarly, other arthropods, predominantly insects were also clearly more abundant at the immediate vicinities of these refuse areas and their populations similarly decreased with increased distance away from the refuse. In the same perspective, lizards, geckoes, toads, frogs and birds were often also more abundant on the ground around the vicinities of these refuses areas.
Effect of farm sanitization on H. venatoria densities: When the plants in the farm were sanitized through the removal of dry leaves and loose barks from the pseudotems, the number of spiders in the sanitized area dropped drastically. There were significantly lower numbers (p<0.05) of spiders on the sanitized plants compared to the non-sanitized plants irrespective of the season (Fig. 4). In most cases, the spider densities in the sanitized pseudostems were at most half of those in the non-sanitized.
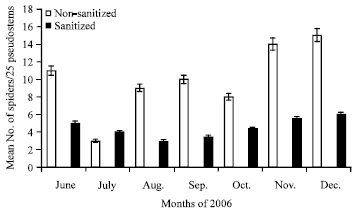 | |
| Fig. 4: | Mean number of Heteropoda venatoria spiders per 25 sanitized and non-sanitized banana pseudostems each during various months of 2006 in Tiko, Cameroon |
DISCUSSION
The various habitats exploited by H. venatoria in banana farms shows that the banana plant architecture and agroecosystem offer a diversity of suitable habitats for the spiders. All these habitats are within close proximity to food sources for the spiders which included preys such as cockroaches, moths, flies, ants as well as colonies of mealybugs, aphids and scale insects on the banana pseudostems. Generally, spiders frequently inhabit plant parts that offer sources of food, shelter and favorable environmental conditions (Arango et al., 2000; Morais-Filho and Romero, 2008). A critical observation of all these habitats revealed that they had many crevices or close fitting sites into which the spiders could hide since they fit closely into the crevices as their preferred sites (Edward, 2009). The flattened bodies and highly flexible long limbs of these spiders enable them easily fit into cracks and crevices. Since, these spiders always hide in close fitting sites, it is apparent that they are positively thigmotactic i.e., contact loving. Generally, the huntsman spider also usually relies on native vegetation and litter for shelter, breeding, feeding and protection against desiccation like in other spiders (Romero and Vasconcello-Neto, 2004).
The vertical and temporal distribution of the spiders in the various habitats appeared to be a function of food availability, absence or presence of predators and distance away from stagnant rain water run-off. Most adults often perched singly facing downward inside the loose leaf sheaths of the pseudostems and stumps at heights of ≤2 m from the bases which often contained stagnant rain water run-off. This is consistent with the observations of Airame and Sierwald, 2000. who observed that on average Heteropoda species perched facing downwards at a distance of 0.93±1.27 m from the water surface in primary forest when hunting. This might have been a strategy to avoid the numerous predators in the litter but at the same time being at close proximity to their abundant preys in the mulches at the bases of the banana mat. Micky and Gary (1992) postulated that such downward orientation is the most practical for detecting prey and/or mate that originates from below the habitat. The solitary nature of adult spiders in the pseudostems may be because of territorial defense in order to ensure access to resources. The higher densities of spiders in the pseudostems and stumps close to refuse areas understandably should be due to the abundance of preys in the assorted litter in the refuse areas. The soil litter had high densities of flies, ants and cockroaches as preys but also harboured many frogs, toads and especially agama lizards and geckoes which preyed heavily on the spiders. This partly explains why the litter spots had the lowest density of spiders and often only during the dry season when fewer frogs and toads were present. The spider population in the litter crashed to zero during the rainy season because at this period, the soils become saturated and pools of stagnant water often avoided by the spiders are very prevalent. Moreover, these stagnant water pools also attracted more of the predatory toads and frogs.
During the banana flowering stage, more insects visited the inflorescences of the plants and thus more spiders were attracted to feed on these insect preys. Subsequently, these spiders also moved into the more sheltered bunches as they developed. Spiders densities in banana bunches increased as the rains began till a peak in October and then dropped as the dry season approached because most of the spiders migrated to the bunches during the rains to seek for refuge (Bradley, 1993) or shelter from the rains. When it rains, the rain water runs off from the banana leaf lamina into the tube-like leaf stalks and flows down the pseudostems. The reverse was true during the dry season when more spiders migrated to the pseudostems/stumps and soil litter in search of preys. At harvest, some of these spiders remain in their hideouts in banana bunches and may eventually be transported to human habitations or to market places. Therefore, the probability of transporting more H. venatoria spiders to human habitations and shops is higher for the rainy than the dry season.
The knowledge of the various habitats of H. venatoria on banana plants can be used to manage the spider population on the crop. For example, when banana loose pseudostyem barks were removed (sanitized) in this study, the spider population dropped to at least half of the densities in non-sanitized pseudostems. This was as a result of the elimination of spider hideouts in the form of the loose pseudostem barks. Given that these spiders are natural enemies of banana insect pests, their populations can be sensibly managed in order to advantageously exploit their beneficial effects. For instance, when improvised banana pseudostem bark collar traps were attached on banana pseudostems, most of the spiders sought refuge in these collar traps rather than the bunches. Such alternative refuges can be used to conserve the spiders on the pseudostems to enable them prey on the noxious aphids, mealybugs and scale insects on banana plants. Since, the huntsman spiders are brownish in color just like the pseudostem barks, the spiders can camouflage better in these traps since they blend with the background. Such cryptic coloration can enhance the efficacy of H. venatoria as natural enemies of banana insect pests. Other predatory spiders use colors for crypsis from their prey insects and their own enemies (Heiling et al., 2003).
CONCLUSION
The tropical huntsman spider prefers close fitting sites and crevices as habitats due to its flattened body and long flexible legs. The banana-agroecosystem and plant architecture provides many of such suitable hideouts including litter spots/mulches, loose pseudostem leaf sheaths, leaf petioles, spaces between flower bracts and clusters and between fingers of bunches. During the rains, most of the spiders migrate into more sheltered habitats notably bunches and hence are more likely to be inadvertently transported with the fruits after harvest. However, banana pseudostem and plantation sanitization practices can advantageously conserve these beneficial natural enemies in the agroecosystem to prey on the inimical arthropod pests of banana.
REFERENCES
- Adeniji, T.A., L.O. Sanni, I.S. Barimalaa and A.D. Hart, 2006. Determination of micronutrients and colour variability among new plantains and banana hybrids flour. World J. Chem., 1: 23-27.
Direct Link - Airame, S. and P. Sierwald, 2000. Hunting and feeding behavior of one Heteropoda species in lowland rainforest on Borneo (Aranae, Sparassidae). J. Arachnol., 28: 251-253.
Direct Link - Arango, A.M., V. Rico-Gray and V. Parra-Tabla, 2000. Population structure, seasonality and habitat use by the green lynx spider Peucetia viridans (Oxyopidae) inhabiting Cnidoscolus aconitifolius (Euphorbiaceae). J. Arachnol., 28: 185-194.
Direct Link - Harwood, J.D. and J.J. Obrycki, 2007. Web-site selection strategies of linyphiid spiders in alfalfa: Implications for biological control. BioControl, 52: 451-467.
CrossRefDirect Link - Heiling, A.M., M.E. Herberstein and L. Chittka, 2003. Pollinator attraction-crab spiders manipulate flower signals. Nature, 421: 334-334.
CrossRefDirect Link - Micky, D.E. and L.M. Gary, 1992. Life cycle and habitat preference of the facultively arboreal wolf spiders. J. Arachnol., 20: 157-164.
Direct Link - Morais-Filho, J.C. and G.Q. Romero, 2008. Microhabitat use by Peucetia flava (Oxyopidae) on the glandular plant Rhyncanthera dichotoma (Melastomataceae). J. Arachnol., 36: 373-378.
Direct Link - Okolle, J.N., M. Mashhor and A. Abu-Hassan, 2006. Seasonal abundance of the banana skipper, Erionota thrax (Lepidoptera: Hesperiidae) and its parasitoids in a commercial plantation and a subsistence farm in Penang, Malaysia. Int. J. Trop. Insect Sci., 26: 197-206.
Direct Link - Oselebe, H.O. and A. Tenkouano, 2009. Ploidy versus gender effects on inheritance of quantitative traits in Musa species. Aust. J. Crop Sci., 3: 367-373.
Direct Link - Romero, G.Q. and J. Vasconcellos-Neto, 2004. Spatial distribution patterns of jumping spiders associated with terrestrial bromeliads. Biotropica, 36: 596-601.
CrossRefDirect Link - Romero, G.Q. and J. Vasconcellos-Neto, 2005. The effects of plant structure on the spatial and microspatial distribution of a bromeliad-living jumping spider (Salticidae). J. Animal. Ecol., 74: 12-21.
CrossRefDirect Link - Tripathi, L., M. Mwangi, S. Abele, V. Aritua, W.K. Tushemereirwe and R. Bandyopadhyay, 2009. Xanthomonas wilt: A threat to banana production in East and Central Africa. Plant Dis., 93: 440-451.
CrossRef
Mbua Christopher Parr Reply
I am the author of this wonderful work. We took a lot of time and sacrifice to do it. We hope to do better and more studies on this huntsman spider.
Good luck