
M. Sharififard
Department of Medical Entomology, School of Health, Jundishapur University of Medical Science, Ahvaz, Iran
M.S. Mossadegh
Department of Plant Protection, College of Agriculture, Shahid Chamran University, Ahvaz, Iran
B. Vazirianzadeh
Department of Medical Entomology and Infectious and Tropical Diseases Research Center, Jundishapur University of Medical Sciences, Ahvaz, Iran
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 5 | Page No.: 282-288







ABSTRACT
The entomopathogenic fungi, Beauveria bassiana IRAN 187C and Metarhizium anisopliae IRAN 437C are the two most virulent isolates against the house fly, Musca domestica L. Effects of temperatures (15, 20, 25, 30, 35°C) and relative humidity levels of (45 and 75%) on virulence of the fungi were detected by immersion and inoculated larval bed methods for larvae and baiting method for adults. The results indicated no significant difference in larval mortality and larval cadaver sporulation at temperatures of 25-30°C and 45-75% RH for Ma 437 C. But Bb 187C caused higher mortality and cadaver sporulation at this thermal range and 75% RH. There was no significant difference in adult mortality between 20, 25 and 30°C for Ma 437 C and between 25 and 30°C for Bb 187 C. Mortality and fly cadaver sporulation were higher at the mentioned thermal range at 75% than 45% RH for both isolates. LT50 values decreased with temperature increasing between 15-30°C but increased again at 35°C. M. anisopliae caused more mortality and cadaver sporulation in wider ranges of temperature and humidity than B. bassiana. So, this isolate would be good candidate for microbial control of the house fly.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: February 16, 2012;
Published: March 27, 2012
How to cite this article
M. Sharififard, M.S. Mossadegh and B. Vazirianzadeh, 2012. Effects of Temperature and Humidity on the Pathogenicity of the Entomopathogenic Fungi in Control of the House Fly, Musca domestica L. (Diptera:Muscidae) under Laboratory Conditions. Journal of Entomology, 9: 282-288.
DOI: 10.3923/je.2012.282.288
URL: https://scialert.net/abstract/?doi=je.2012.282.288
DOI: 10.3923/je.2012.282.288
URL: https://scialert.net/abstract/?doi=je.2012.282.288
INTRODUCTION
The house fly, Musca domestica (L.) is one of the most active vectors of disease to affect humans. It quickly develops resistance to most chemical insecticides but entomopathogenic fungi are promising candidates for microbiological control of this pest (Geden et al., 1995). High rates of infection and rapid killing of the house fly by the hyphomycetes fungi Beauveria bassiana and Metarhizium anisopliae have been reported in numerous studies (Steinkraus et al., 1990; Barson et al., 1994; Bywater et al., 1994; Watson et al., 1995, 1996; Renn et al., 1999; Lecouna et al., 2005; Kaufman et al., 2005; Sharififard et al., 2011a).
There are parameters other than its pathogenicity that must be considered in the development of a fungal isolate suitable for use as a mycoinsecticide. In particular the environmental conditions under which the fungus will be active should take in to account during isolate selection (McCammon and Rath, 1994). Environmental factors that influence the virulence of entomopathogens must be considered for the successful development of the fungus as a bio-control agent. Of all the factors that influence epizootic qualities of a mycopathogen, none is more critical for sporulation, germination and invasion of the host than high humidity (RH>90%).
The rapidity of mycelia development and therefore, the rapidity of the evolution of infection depend on temperature. In general, optimum values fall between 20°C and 30°C (for example, 23°C for Beauveria brongniartii, 24°C for Entomophthora obscura, 25°C for Beauveria bassiana and Nomuraea rileyi and 27-28°C for Metarhizium anisopliae) with lower and upper limits of 5 and 35°C. Temperatures lower than the optima distinctly retard the development of mycosis without necessarily affecting total mortality (Ferron, 1978).
This study investigates the effects of humidity and temperature on the infection potential of fungal isolates, B. bassiana Iran 187C and M. anisopliae Iran 437C that their virulence has been demonstrated to M. domestica (Sharififard et al., 2011b).
MATERIALS AND METHODS
M. domestica: Adult house flies were collected from a dairy farm with a sweeping net and transferred to the laboratory where they were reared at 26°C, 50±5% Rh and photoperiod of 14:10 (L:D). Adults were maintained in cages (40x40x40 cm3) covered by gauze. Water and food in the form of sugar and powdered milk were provided and replenished every 24-48 h. The larval medium comprised of 55 g wheat bran, 3 g date extract and 2 g dried alfalfa suspended in 140 mL water. One cup (250 mL volume) of this medium was left in each cage for adult oviposition and subsequent development of larvae. The food was replaced every 24-48 h.
Fungi: A previous study has indicated that M. anisopliae strain IRAN 437 C and B. bassiana IRAN 187 C were the most virulent isolates against the house fly, M. domestica which caused higher mortality in shorter time than others (Sharififard et al., 2011a) so these isolates were selected for this study. They were cultured on Sabouraud dextrose agar with yeast extract (SDAY) for 2 weeks at 27°C, 75±5% RH and photoperiod of 12:12 (L:D). Sporulation cultures were harvested by scraping the dry conidia from the surface of the culture plate with a scalpel and depositing them in to sterile distilled water containing 0.01% Tween-80. Concentration of the suspension was determined using a hemocytometer.
Effect of temperature and humidity on infection of larval house fly by fungi
Larval Immersion method: To test the larva, an aqueous suspension of 108 conidia mL-1 was used. Groups of 25 medium size larvae were immersed simultaneously for 10 sec in the conidial suspension and each group was transferred to the larval medium in a white disposable container. Control groups were treated with 0.01% Tween-80 solution. There were four replicates for each treatment. All larval containers were incubated at 15, 20, 25, 30, 35°C and for each temperature at 45 and 75% relative humidity.
Inoculated larval bed method: Plastic cups containing 50 g larval medium were inoculated with 1 mL of stock solution to give 109 conidia/g of each tested isolates. Groups of 25 larvae were released in to each cup. Control groups were treated with a 0.01% Tween-80 solution. There were four replicates for each treatment. All larval containers were incubated at 25, 30°C and for each temperature at 45 and 75% relative humidity. In both larval methods dead larva were held in larval bedding to monitor mortality induced by the fungi. Externally sporulated cadavers were recorded as dead due to fungal infection.
Effect of temperature and humidity on infection of adult house fly by fungi: 108 conidia g-1 of fungi were inoculated with a semi-solid bait containing sugar, powdered milk and water. A 9 cm diameter Petri dish containing 10 g of the inoculated bait was left in each cage. Twenty adults were left in each cage and each treatment consisted of four replicates. The cages were then incubated at 15, 20, 25, 30, 35°C and for each temperature at 45 and 75% relative humidity. Cages were checked daily over a period of 9 days following the presentation of fungus. The control group was fed with safe bait. Cadavers were removed and the surface was sterilized and transferred to a sterile Petri dish with damp filter paper. Sporulating cadavers were recorded as mortality due to fungal infection.
Data analysis: Data from this study were analyzed by factorial analysis of variance (ANOVA) using a two factor complete randomized design in MSTATC software. Percentages of mean of mortality were compared using Duncan’s multiple range test at α = 0.05. LT50 values and 95% confidence limits of each value for different treatments were calculated by using probit method with SAS software. When there was no overlap at a 95% CL of lethal time values, the difference between treatments was considered significant.
RESULTS
Effect of temperature and humidity on infection of larval house fly by fungi: The results of larval immersion in 108 conidia mL-1 suspension of M. anisopliae showed that larval mortality was completely temperature dependent and increased from 15-30°C but decreased again at 35°C. M. anisopliae produced the greatest mortality at the temperatures of 25 and 30°C in both 45 and 75% RHs. This fungal isolate had similar efficacy in controlling house fly larva at the thermal range of 25-30°C and relative humidity of 45-75% RH (Table 1).
| Table 1: | Effects of temperature and humidity on virulence of M. anisopliae IRAN 437C against M. domestica |
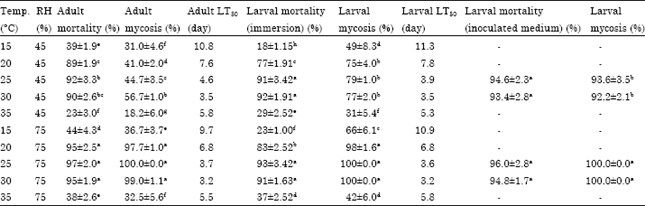 | |
| ANOVA results: For larvae: Differences between mortality: Temperatures (F = 393.46, df = 4, p<0.0001), Relative humidities (F = 8.85, df = 1, p = 0.0057) and Interaction of temperature and relative humidity (F = 0.759, df = 4). For fungal sporulation on larval cadavers: Temperatures (F = 50.74, df = 4, p<0.0001), Relative humidities (F = 60.09, df = 1, p<0.0001) and Difference in interaction of temperature and relative humidity was not significant (F = 0.1735, df = 4), For adults: Differences between mortality: Temperatures (F = 272.67, df = 4, p<0.0001), Relative humidities (F = 17.84, df = 1, p = 0.0002) and The interaction treatments of temperature and relative humidity (F = 1.32, df = 4, p = 0.285). For fly cadaver sporulation: Temperatures (F = 115.59, df = 4, p<0.0001), Relative humidities (F = 300.09, df = 1, p<0.0001) and Interaction of temperature and relative humidity (F = 27.64, df = 4, p<0.0001) | |
Mortality was statistically higher at 25-30°C levels and exceeded 70% at 20°C in both humidity variables (45 and 75% RH). In spite of equality of mortality rates both 45 and 75% RH, the cadaver sporulation (mycosis) was greatest at the thermal range of 20-30°C and 75% RH and mycosis means were not different but percentage of sporulation at 45% RH and these temperatures should also be taken into consideration (Table 1). In addition, the results from the inoculated bed with conidium of M. anisopliae confirmed that the difference between larval mortality was not significant at the temperature range of 25-30°C and relative humidity of 45-75% RH. Larval mortality was similar in the thermal range of 25-30°C and 45-75% RH but cadaver sporulation (mycosis) was higher at this thermal range and 75% RH (Table 1). The lethal time for larval killing was completely dependent on temperature and relative humidity. At constant humidity, LT50 values decreased with temperature increase from 15-30°C and increased again at 35°C. Larval mortality time was not significantly different in both studied RHs at constant temperature.
Larval mortality and cadaver sporulation with B. bassiana were equal in the temperature range of 25-30°C with 75% RH compared to 45%. In the thermal range of 25-30°C with 75% RH larval mortality and cadaver sporulation were 81-82 and 99-100%, respectively. In this thermal range with 45% RH mortality and cadaver sporulation were 50 and 60%, respectively. The test results of the inoculated larval bed confirmed the results of larval immersion. So, Bb 187C caused the highest mortality and cadaver sporulation at the mentioned thermal range and 75% RH (Table 2).
Effect of temperature and humidity on infection of adult house flies by fungi: Virulence of Ma 437 C against adults was alike at temperatures of 20, 25 and 30°C in each relative humidity level. The highest mortality occurred in the thermal range of 20-30°C and 75% RH but it was remarkable and exceeded 80% in this thermal range and 45% RH. Fly cadaver sporulation (mycosis) with Ma 437 C was equal in 20, 25, 30°C and 75% RH and the greatest mycosis was observed at the thermal range of 20-30°C (Table 1).
| Table 2: | Effects of temperature and humidity on virulence of B. bassiana IRAN 187C against M. domestica |
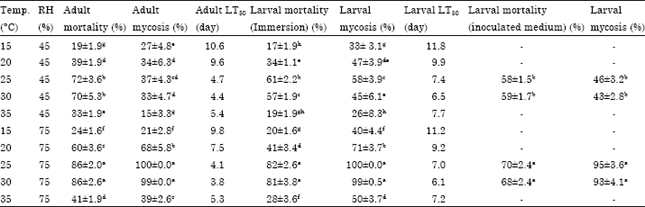 | |
| ANOVA results: For larvae: Differences between mortality: Temperatures (F = 131.97, df = 4, p<0.0001), Relative humidities (F = 43.8, df = 1, p = 0.0057) and The interaction of temperature and relative humidity (F = 4.52, df = 4, p = 0.006). For larval cadaver sporulation (mycosis): Temperatures (F = 31.67, df = 4, p<0.0001), Relative humidities (F = 23.2, df = 1, p<0.0001). For adults: Differences between mortality: Temperatures (F = 160.09, df = 4, p<0.0001), Relative humidities (F = 51.2, df = 1, p<0.0001) and The interaction treatments of temperature and relative humidity (F = 2.54, df = 4, p = 0.06). For fly cadaver sporulation: Temperatures (F = 54.15, df = 4, p<0.0001), Relative humidities (F = 197.9, df = 1, p<0.0001) and Interaction of temperature and relative humidity (F = 27.2, df = 4, p<0.0001) | |
Pathogenicity of Bb 187 C in the adult control was not significantly different in 25 and 30°C in a constant humidity. But adult mortality and fly cadaver sporulation levels were higher in the temperature range of 25-30°C and 75% RH than at 45% RH. However no sporulation occurred in the mentioned temperature range and 45% relative humidity but adult mortality rate in these conditions was (70-72%) (Table 2).
So, similar to the larval results the optimum temperature and relative humidity for activity of Bb 187C in the control of adult house fly was 25-30°C and 75%.
Recorded mycosis with both isolates was the result of leaving the sterilized fly cadavers in the Petri dish with damp filter paper and then incubating at 75% RH. At 45% RH, sporulation occurred only in this condition and no sporulation was observed when fly cadavers were left the humid environment. So, the greatest virulence was observed in the thermal range of 20-30°C with 75% RH for Ma 437C and in 25-30°C with 75% RH for Bb187C. Also the greatest mycosis for both strains was observed in these conditions.
Lethal time for adult killing decreased with temperature increase from 15-30-35°C and decreased with increased RH. Consequently, the shortest lethal time was recorded at thermal range of 20-30°C and 75% RH (Table 1, 2).
DISCUSSION
It has been demonstrated that relative humidity is not a key factor in the infection of M. domestica larva by M. anisopliae IRAN 437C compared to B. bassiana. Ma 437C caused higher mortality and cadaver sporulation in a wider temperature and humidity ranges (20-30°C and 45-75%) than Bb 187C (25-30°C and 75%) but Bb 187C pathogenicity in larval populations also occurred at 45% RH.
It can be asserted that moisture and temperature of the bedding probably supported the development of mortality and sporulation of entomopathogenic fungi in the larval population at 45% RH. It is thought that such larvae persisting in the bedding could serve as inoculum for subsequent infection of larva. Larval mortality significantly decreased at 35°C in both humidity levels possibly due to the effect of high temperature on shortening the larval growing period, acceleration of larval ecdysis or due to an inappropriate impact on spore growth and germination. This result was confirms the research by Tang and Hou (2001) that reports the percentage mortality of Helicoverpa armigera larva decreased from 96.2% at 30°C to 3.3% at 35°C.
Although, higher relative humidity resulted in higher cadaver sporulation and thus the production of more inoculum to induce fungal epizootiology in the larval population but in an environment with low humidity the necessary threshold of inoculum for fungal survival and epizootiology is provided in the larval bed.
Humidity influences the pathological effect of fungi mainly through sporulation and initiation of spore germination on insect integument. However, it is not always necessary to maintain the inoculated larvae in high humidity for better infection (Tang and Hou, 2001). Lethal time for house fly larvae decreased with humidity (45-75% RH) but the difference was not statistically significant. It seems that relative humidity of an environment does not affect the lethal time scale of house fly larva because of existing the favorable humidity for fungus pathogenicity and spore germination in the larval bed. Larval LT50 decreased with temperature increase from 15-30°C. Tang and Hou (2001) reported that lethal time of H. armigera larva decreased from 11.9 day at 15°C to 7.3 day at 30°C but was not measurable at 35°C.
Cadaver sporulation is a necessary factor for epizootiology of entomopathogenic fungi in insect population and fungal dispersion. In this study it was found that the isolate Ma 437 C was a suitable option to achieve this goal. Because sporulation of this isolate on larval cadavers occurred easily in a wide range of temperatures and humidity levels.
Adult control by both isolates showed higher efficacy of Ma 437 C against adult populations in wider temperature range of (20-30°C) than Bb 187C (25-30°C). Despite the creation of adult mortality by both isolates at 45% RH, no sporulation occurred normally at this relative humidity. Our results have been confirmed by other researchers that pathogenesis occurred at much lower ambient values probably because of high humidity in the microclimate at the insect cuticle. It is certain, however that the external sporulation never occurs on the killed insect, if relative humidity is too low (Ferron, 1978; Ramoska, 1982; Talwar, 2005).
As most house fly activity occurs in places such as the inside poultry houses, livestock houses and human dwellings and that these places are somewhat favorable for fungal activity in the terms of temperature, relative humidity and light, therefore the killing potential of the M. anisopliae IRAN 437 C may be supported by these conditions.
The rapidity of mycelial development and therefore, the rapidity of the evolution of infection depend on temperature. In general, optimum values fall between 20 and 30°C with limits between 5 and 35°C. Temperatures lower than the optima distinctly retard the development of mycosis without necessarily affecting total mortality (Ferron, 1978).
The optimum temperature for development of the fungus is not necessarily the same as for development of disease. However, the influence of temperature on the host insect must be taken into consideration, since very short periods between moultings resulting from high temperature may reduce, for example, the duration of the instar to an extent that penetration of the fungus through the integument is impeded (Talwar, 2005).
Overall, the results showed that relatively good control was created by M. anisopliae Iran 437C on the house fly population in a wider range of temperature (20-30°C) and relative humidity (45-75%) compared to Bb Iran 187C. However, if fungal spore is released for adult control in low ambient humidity, no cadaver sporulation will occur and so the necessary inoculums to create epizootiology in adult population will not grow. As the usual method for controlling insect pests by entomopathogenic fungi is the release of abundant spores and the ultimate goal is to reduce the adult house fly population by means of the release of spores in to the environment, then relatively satisfactory insect control will be achieved.
M. anisopliae IRAN 437 C induced more mortality in the larva and adult population of house fly and higher cadaver sporulation in the wider range of temperature and humidity. Furthermore, synergistic effect of this fungal isolate with spinosad insecticide as a microbial and safe insecticide (Sharififard et al., 2011b) and growth potential of mycelia in low ambient humidity (45-50% RH) which are the reasons that this isolate would be recommended as a successful and promising biological agent for development as mycoinsecticide for the control of house fly.
ACKNOWLEDGMENT
The authors wish to thanks Iranian Research Institute of Plant Protection for providing the fungal isolates used in the study.
REFERENCES
- Barson, G., N. Renn and A.F. Bywater, 1994. Laboratory evaluation of six species of entomopathogenic fungi for control of house fly, Musca domestica L., a pest of intensive animal units. J. Invertebr. Pathol., 64: 107-113.
PubMed - Ferron, P., 1978. Biological control of insect pests by entomogenous fungi. Annu. Rev. Entomol., 23: 409-442.
CrossRef - Geden, C.G., D.A. Rutz and D.C. Steinkraus, 1995. Virulence of different isolates and formulation of Beauveria bassiana for house flies and the Parasitoid muscidofurax raptor. J. Bio. Conf., 5: 615-621.
CrossRef - Kaufman, P.E., C. Reasor, D.A. Rutz, J.K. Ketzis and J.J. Arends, 2005. Evaluation of Beauveria bassiana applications against adult house fly, Musca domestica, in commercial caged-layer poultry facilities in New York State. Bio. Control, 33: 360-367.
CrossRefDirect Link - Lecouna, R.E., M. Turica, F. Tarocco and D.C. Crespo, 2005. Microbial control of musca domestica (Diptera: Musciadae) with selected strains of beauveria bassiana. J. Med. Entomol., 42: 332-336.
PubMed - McCammon, S.A. and A.C. Rath, 1994. Separation of Metarhizium anisopliae strains by temperature dependent germination rates. Mycol. Res., 98: 1253-1257.
Direct Link - Ramoska, W.A., 1982. An examination of the long term epizooti potential of various artificially introduced mosquito larval pathogens. Mosq. News, 42: 603-607.
Direct Link - Sharififard, M., M.S. Mossadegh, B. Vazirianzadhe and A.Z. Mahmoudabadi, 2011. Interactions between Entomopathogenic Fungus, Metarhizium anisopliae and Sublethal Doses of Spinosad for Control of House Fly, Musca domestica. Iran J. Arthropod-Borne Dis., 5: 28-36.
Direct Link - Renn, N. and A.F. Bywater and G. Barson, 1999. A bait formulation with metarhizium anisopliae for the control of Musca domestica (Diptera: Musciadae) assessed in Large scale Laboratory enclosures. J. Applied Entomol., 123: 309-314.
CrossRef - Steinkraus, D.C., C.J. Geden, D.A. Rutz and J.P. Kramer, 1990. First report of the natural occurrence of Beauveria bassiana (Moillales: Moniliaceae) in Musca domestica (Diptera: Musciadae). J. Med. Entomol., 27: 309-312.
Direct Link - Tang, L.C. and R.F. Hou, 2001. Effects of environmental factors on virulence of the entomopathogenic fungus, Nomuraea rileyi, against the corn earworm, Helicoverpa armigera (Lep., Noctuidae). J. Applied Entomol., 125: 243-248.
CrossRef - Watson, D.W., C.G. Geden, S.J. Long and D.A. Rutz, 1995. Efficacy of Beauveria bassiana for controlling the house fly and stable fly (Diptera: Musciadae). J. Bio. Conf., 5: 405-411.
CrossRef - Watson, D.W., D.A. Rutz and S.J. Long, 1996. Beauveria bassiana and sawdust bedding for the management of the house fly, Musca domestica (Diptera: Muscidae) in calf hutches. Bio. Control, 7: 221-227.
CrossRef - Sharififard, M., M.S. Mossadegh, B. Vazirianzadeh and A. Zarei Mahmoudabadi, 2011. Laboratory evaluation of pathogenicity of entomopathogenic fungi, Beauveria bassiana (Bals.) Vuill. and Metarhizium anisopliae (Metch.) Sorok. to larvae and adults of the house fly, Musca domestica L. (Diptera: Muscidae). Asian J. Biol. Sci., 4: 128-137.
CrossRefDirect Link