
Nesrin A. El-Basha
Department of Biological Control, Plant Protection Research Institute, Agriculture Research Center, Giza, Egypt
M.S. Salman
Department of Vegetable and Aromatic Plant Mites, Plant Protection Research Institute, Agriculture Research Center, Giza, Egypt
M.A. Osman
Department of Agricultural Zoology, Faculty of Agriculture, Mansoura University, Mansoura, Egypt
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 5 | Page No.: 248-256







ABSTRACT
The objective of this study was to determine the functional response of the predator Orius albidipennis (Reuter) fed on egg of Tetranychus urticae Koch. We conducted a logistic regression of the proportion of prey consumed as a function of initial prey density to identify functional response types and used nonlinear least-squares regression and the random predator equation to estimate attack rates and handling times. Overall, all stages of O. albidipennis exhibited a type-I functional response to T. urticae. Whereas, attack rate (a) and handling time (Th) of O. albidipennis female and male recorded 1.267, 0.828, 0.0122 and 0.0141 when offering eggs of T. urticae, respectively.
PDF Abstract XML References Citation
Received: December 05, 2011;
Accepted: February 15, 2012;
Published: March 27, 2012
How to cite this article
Nesrin A. El-Basha, M.S. Salman and M.A. Osman, 2012. Functional Response of Orius albidipennis (Hemiptera: Anthocoridae) to the Two-spotted Spider Mite Tetranychus urticae (Acari: Tetranychidae). Journal of Entomology, 9: 248-256.
DOI: 10.3923/je.2012.248.256
URL: https://scialert.net/abstract/?doi=je.2012.248.256
DOI: 10.3923/je.2012.248.256
URL: https://scialert.net/abstract/?doi=je.2012.248.256
INTRODUCTION
The two spotted spider mite Tetranychus urticae Koch. (Acari: Tetranychidae), considers one of the most important pests around the world in various ornamental and vegetable crops (Zaher, 1986; Childers, 1994; Takafuji et al., 2000; Zhang, 2003; Alatawi et al., 2005; Abd-Elhady and Heikal, 2011). On other hand, Insect predators are most effective regulators of pest populations (Cardoso and Lazzari, 2003; Padmalatha et al., 2003), which has led their increasing use in insect pest management programs (Wiedenmann and Smith, 1997; Riudavents and Castane, 1998). Due to, the current concerns to reduce the excessive use of chemicals insecticides for pest control, as well as to avoid increments in doses or utilization of highly toxic compounds (Gravena, 1989) biological control stands as a profitable alternative to the use of chemicals in the agroecosystem (Lester et al., 2000).
Predation is supposed to be one of the great biotic mortality factors reducing insect pest populations and using them in insect pest management programs has been receiving better consideration because of the recent need to reduce the exclusive apply of insecticides for pest control (Sarmento et al., 2007).
Members of the genus Orius (Heteroptera: Anthocoridae) are important predators of certain arthropods like aphids, aleyrodids, eggs and larvae of lepidopterous, thrips and mites (Pericart, 1972; Vacante et al., 1997; Reitz et al., 2006; Butler and O’Neil, 2007; Fathi and Nouri-Ganbalani, 2009). However, thrips and mites are believed to represent imperative diets for Orius spp. (Wright, 1994). Furthermore, several species of Orius have been received considerable attention in biological control programs due to their efficiency in number of agricultural ecosystems (Cocuzza et al., 1997). Additionally the previous literatures stated that Orius spp. could effectively suppress spider mite populations in the field (Oatman and McMurthry, 1966; Du and Yan, 1995; Wittmann and Leather, 1997; Sanderson et al., 2005; Rosenheim, 2005; Xu and Enkegaard, 2009). Of them, Orius albidipennis (Reuter) (Heteroptera: Anthocoridae) was found in the south Mediterranean basin, in the Canary Islands and East Africa (Salim et al., 1987; Chyzik et al., 1995; Fritsche and Tamo, 2000). In Egypt, O. albidipennis is very common in cultivated areas, especially in corn and cotton fields. It is generally establish in flowers of plants infested with thrips, lepidopteran eggs or other small arthropods. O. albidipennis does not occur in the field earlier than March, but its activity increases from April until the end of November (Tawfik and Atta, 1974; Zaki, 1989; Sobhi et al., 2010).
Prior to the release of a natural enemy in a biological control program, it is essential to evaluate its efficiency under laboratory conditions. One useful method for evaluating the efficiency of a natural enemy is to assess of their behavioral characteristics including functional response and searching rates (Fathipour et al., 2006; Bayoumy et al., 2009). The relationship among the number of prey consumed per predator individual and prey density was defined as functional response (Solomon, 1949; Holling, 1959a, b). It plays a critical role in the perspective of prey-predator interactions and their ecological and evolutionary consequences (Tully et al., 2005). Holling (1959a) identified three basic types of functional responses in general. The Type I response is characterized by a linear rise with a constant attack rate over all prey densities until satiation is reached. In the Type II response the attack rate decreases as prey density increases. Type III is represented by a sigmoid curve, where the attack rate increases with increasing prey density. Holling (1961) divided the functional response into several basic and subsidiary components. The attack rate (a) can be considered as a function of: (1) the reaction distance of the predator, i.e., the maximum distance at which the predator will react by attacking prey, (2) the speed movement of predator and prey and (3) the proportion of attacks that are successful. The handling time (Th) can be considered to be a function of: (1) the time spent pursuing an individual prey, (2) the time spent investigating and probing each prey and (3) the time spent drilling each prey. The time as prey and predator exposure (T) can be considered to be a function of: (1) time in non-feeding activities and (2) time in feeding-related activities (i.e., Th). The objective of this study was to investigate the functional response of O. albidipennis stages when preying on egg of T. urticae to improve our understanding of prey-predator interaction and get a better strategy for the biological control of T. urticae using O. albidipennis.
MATERIALS AND METHODS
Mite colony: The stock colony of Tetranychus urticae was collected from eggplant at the experimental farm, Ismailia Agricultural Research Station. T. urticae was collected from the same locations and host plants used for the collection of Orius albidipennis. The colony of T. urticae was kept on detached sweet potato branches kept with their upper part of the stem in contact with water in glass vials at 25±1°C, 50-80% RH and photoperiod was 14L/10D h.
Rearing of Orius albidipennis: A colony of O. albidipennis was established from nymphs and adults collected on eggplant plants (Solanum melongena L.) infested with T. urticae at the experimental farm, Ismailia Agricultural Research Station, Ismailia, Egypt. Adults and nymphs were maintained in plastic jars (10 cm diameterx20 cm height), which were covered with muslin that was held in place by rubber bands. Each jar was provided with sufficient quantities of T. urticae as a food supply, a piece of cotton that had been soaked in a 10% honey solution and bean pods (Phaseolus vulgaris L.) as an oviposition substrate. Bean pods with newly deposited eggs were removed and replaced daily, kept in previous jars. Jars were examined daily for hatching, after hatching nymphs were provided with T. urticae and small balls of foam to reduce cannibalism. Upon eclosion, adult males and females were sexed and placed in new plastic jars, provided with the same time of prey and oviposition substrates. Colonies were maintained at 26±1°C and 60±10% RH.
Functional response: The predator O. albidipennis larvae (4 h post molting) were collected from the colony and starved for 4 h in glass vials (7 cmx2 cm) containing small wet cotton with water without preys. Adult males and females 7 days old were collected in the same size of glass vials and starved 24 h before being used. Small discs 1.5 cm of sweet potato leaves harboring eggs of T. urticae was prepared and number of eggs was counted and put in Huffaker cells. Starved predators were singly transferred to modified Huffaker cells. T. urticae eggs were introduced as prey into modified Huffaker cell at six densities increased gradually to be synchronize with the developmental stage of the predators. Whereas, six prey densities of T. urticae were evaluated: 30, 40, 50, 60, 70 and 80 eggs for the fourth instar, fifth instar and adult males and females, respectively. Each predator individuals (larva and adult) was replicated ten times. After 24 h, numbers of consumed eggs of T. urticae were recorded.
Data analysis: The approach developed by Juliano (2001) was used to analyze the predator’s functional response. Initially, the type (shape) of response was determined by seeing if the data pit a type I, II or III response, using a polynomial logistic regression of the proportion of prey consumed (Na/No) versus the initial number of prey offered (No) as follows:
| (1) |
where, Na is the number of prey consumed, N0 is the initial number of prey density and P0, P1, P2 and P3 are the intercept, linear, quadratic and cubic coefficients, respectively. These parameters can be estimated using the CATMOD procedure in SAS (Juliano, 2001). The logistic regression was used to obtain the maximum likelihood estimates of parameters P0 to P3. The functional response type was determined by the sign of the linear coefficient from Eq. 1 and the significance of the parameters from the logistic model was evaluated by log likelihood tests. For a type I, the curve of Na/No versus No has a linear shape if the linear term from Eq. 1 was not significantly different from 0, a type I functional response was indicated , whereas a significant negative value indicated a type II response and a significant positive value indicated a type III (Juliano, 2001). The second part of the analyses used Holling disk equation (Holling 1959a) to estimate the parameter values of type I as follows:
where, Ha defines the number of prey attacked by a predator per time unit, a is attack rate of a predator, H is the original number of prey items offered to each predator at the beginning of the experiment, T is the total time of exposure time (1 day in this experiment) and Th is handling time for each prey caught (proportion of the exposure time that a predator spends in identifying, pursuing, killing, consuming and digesting prey).
The parameters a (the rate of successful attack) and Th (the time required to handle a prey item) were calculated using least-squares non-linear regression. Whereas, Th values were used to calculate maximum attack rate as T/Th (Hassell, 1978), this represent the maximal number of prey individuals that could be consumed by O. albidipennis during 24 h.
RESULTS AND DISCUSSION
Functional response, though an important tool, cannot only be attributed to report success and failures in biocontrol programs. For instance, other factors, such as intrinsic growth rates, host patchiness, predation and competition, host traits and environmental complexities (abiotic and biotic factors) also have a major influence on the efficiency of predator in managing the prey population (Pervez and Omkar, 2005).
The average number of T. urticae attacked by O. albidipennis increased with prey density during a 24 h period. Prey consumption by 4th, 5th, male and female of O. albidipennis increased 20.4 to 33.4, 23 to 38.4, 18.2 to 32.4 and 26 to 43.4 individual with increase in density of eggs of T. urticae, respectively. While, the proportion of killed preys by 4th, 5th, male and female of O. albidipennis decreased from 0.68 to 0.41, 0.76 to 0.48, 0.6 to 0.4 and 0.86 to 0.54 with increase in density of eggs of T. urticae, respectively (Fig. 1a, b). Decreasing in proportion of prey consume with increasing prey density is common for arthropods predator Holling (1961). Jalalizand et al. (2011) showed that functional response of Orius niger niger Take type II when fed on adult of T. urticae while it take type III when fed on egg of T. urticae. Gitonga et al. (2002) reported a functional Type I response curve of O. albidipennis preying on Megalurothrips sjostedti Trybom second instar larvae and adults at various densities.
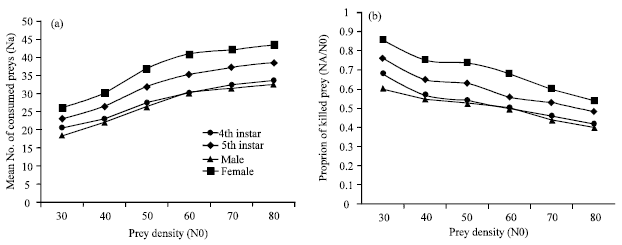 | |
| Fig. 1 (a, b): | Observed functional response of Orius albidipennis 4th, 5th, male and female to Tetranychus urticae egg densities |
| Table 1: | Results of logistic regression analyses, indicating estimates and standard errors of linear, quadratic and cubic coefficient for the proportion of prey eaten by O. albidipennis against initial prey number offered at 25°C |
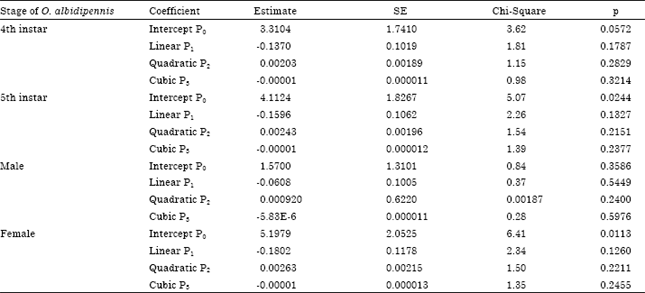 | |
Data presented in Table 1 showed that the outcome of the logistic regression 4th, 5th instars, male and female to densities of eggs of Tetranychus urticae reflected a type I functional response, in all cases the sign of the linear term was negative and p>0.05 (Table 1).Whereas, the type of functional response can be determined based on the sign of the linear coefficient (Juliano, 1993). The type of functional response and estimated parameters for a natural enemy could be affected by some factors such as host plant. Temperature and type of prey or host and prey stages (Wang and Ferro, 1998; De Clercq et al., 2000; Mohaghegh et al., 2001). Xu and Enkegaard (2009) noticed that female of Orius sauteri (Poppius) reflected a type I functional response to densities of deutonymph of T. urticae. Whereas, Zamani et al. (2009) showed that the functional response of O. albidipennis to densities of female of T. urticae take type II. Although, Jalalizand et al. (2011) reported that O. niger female exhibited a type II and III functional response in their predation of T. urticae female and eggs, respectively.
The functional response data of O. albidipennis 4th, 5th, male and female to densities of eggs of Tetranychus urticae were successfully fitted to the Holling disk equation (Holling, 1959a) (Fig. 2, Table 2).
Attack rate and handling time were the parameters used to determine the magnitude of functional responses Pervez and Omkar (2005). The attack rate of O. albidipennis 4th and 5th was 1.613 and 1.148 whereas, male and female of O. albidipennis attack rate was 0.828 and 1.267 (Table 2).
Xu and Enkegaard (2009) showed that attack rate of O. saurteri female on T. urticae was 0.042. Whereas, Jalalizand et al. (2011) reported that attack rates of O. niger on both cucumber and strawberry for T. urticae female was 0.021 and 0.045, respectively. While, attack rates of O. niger on the same host for T. urticae egg was 0.001 and 0.003, respectively. On other hand, handling time is a good indicator of consumption rate and predator efficacy because it reflects the cumulative effect of time taken during capturing, killing, subduing and digesting the prey(Veeravel and Baskaran, 1997). Handling time (Th) O. albidipennis 4th, 5th, male and female between 0.0122 and 0.0225.
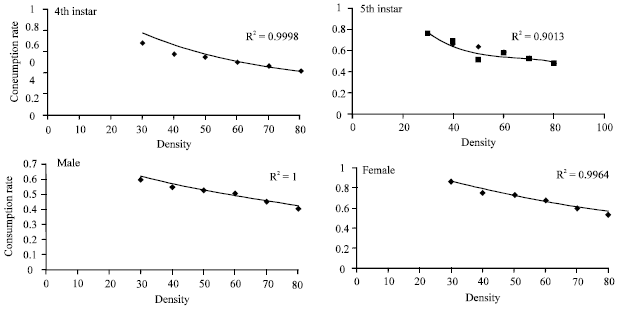 | |
| Fig. 2: | Consumption rate of Orius albidipennis ,4th, 5th, male and female on eggs of Tetranychus urticae at 25±1°C, The fitted lines are predictions of the Holling (1959a) model |
| Table 2: | Effect of Tetranychus urticae densities on the attack rate (a), handling time (Th) and maximum number of consumption (T/Th) on O. albidipennis derived from random predator equation |
 | |
Whereas, The expected maximum consumption (T/Th) of O. albidipennis 4th, 5th, male and female between 44.44 and 81.96 eggs per day of T. urticae (Table 2). Xu and Enkegaard (2009) reported that handling time (Th) of O. saurteri female on T. urticae deutonymph was 1.217. Whereas, Zamani et al. (2009) showed that handling time (Th) for the O. albidipennis when reared on T. urticae ranged between 0.005 to 0.012 days and the highest theoretical maximum predation rate was (80 adult).
In conclusion, the present study has improved our understanding of the O. albidipennis behavior to T. urticae. However, functional responses studies in small laboratory arenas have been criticized as being unrepresentative of natural conditions (Kareiva, 1990) and should be interpreted with care. We believe they may have some value as a first step in estimating predatory capacity but recommend that additional studies be conduct.
REFERENCES
- Abd-Elhady, H.K. and H.M.M. Heikal, 2011. Selective toxicity of three acaricides to the two-spotted spider mite Tetranychus urticae and predatory mite Phytoseuilus persimilis in apple orchards. J. Entomol., 8: 574-580.
CrossRefDirect Link - Alatawi, F.J., G.P. Opit, D.C. Margolies and J.R. Nechols, 2005. Within-plant distribution of two-spotted spider mites (acari: Tetranychidae) on impatiens: Development of a presence-absence sampling plan. J. Econ. Entomol., 98: 1040-1047.
PubMed - Cardoso, J.T. and S.M.N. Lazzari, 2003. Comparative biology of Cycloneda sanguine (Linnaeus, 1763) and Hippodamia convergens (Guerin-Meneville, 1842) (Coleoptera, Coccinellidae) focusing on the control of Cinara spp. (Hemiptera, Aphididae). Revista Brasileira de Entomol., 47: 443-446.
Direct Link - Chyzik, R., M. Klein and Y. Ben-Dov, 1995. Reproduction and survival of the predatory bug Orius albidipennis on various arthropod prey. Entomologia Experimentalis Applicata, 75: 27-31.
CrossRefDirect Link - Cocuzza, G.E., P. de Clercq, M. van de Veire, A. de Cock, D. Degheele and V. Vacante, 1997. Reproduction of Orius laevigatus and Orius albidipennis on pollen and Ephestia kuehniella eggs. Entomol. Exp. Appl., 82: 101-104.
Direct Link - De Clercq, P., J. Mohaghegh and L. Tirry, 2000. Effect of host plant on the functional response of the predator Podisus nigrispinus (Heteroptera: Pentatomidae). Biol. Cont., 18: 65-70.
Direct Link - Du, X.G. and Y.H. Yan, 1995. Manipulating the ground cover of apple orchard to increase the number of Orius sauteri in apple trees. Chinese J. Biol. Control, 11: 1-4.
Direct Link - Fathi, S.A.A. and G. Nouri-Ganbalani, 2009. Prey preference of Orius niger (Wolf.) and O. minutus (L.) from Thrips tabaci (Lind.) and Tetranychus urticae (Koch.). J. Entomol., 6: 42-48.
CrossRefDirect Link - Fritsche, M.E. and M. Tamo, 2000. Influence of thrips prey species on the life-history and behaviour of Orius albidipennis. Entomologia Experimentalis Applicata, 96: 111-118.
Direct Link - Gitonga, L.M., W.A. Overholt, B. Lohr, J.K. Magambo and J.M. Mueke, 2002. Functional response of Orius albidipennis (Hemiptera: Anthocoridae) to megalurothrips sjostedt (Thysanoptera: Thripidae). Biol. Control, 24: 1-6.
CrossRef - Holling, C.S., 1959. Some characteristics of simple types of predation and parasitism 1. Can. Entomol., 91: 385-389.
CrossRef - Holling, C.S., 1959. The components of predation as revealed by a study of small mammal predation of the European pine sawfly. Can. Entomol., 91: 293-320.
CrossRef - Jalalizand, A., M. Modaresi, S.A. Tabeidian and A. Karimy, 2011. Functional response of Orius niger niger (Hemiptera: Anthocoridae) to Tetranychus urticae (Acari: Tetranychidae): Effect of host plant morphological feature. Int. Conf. Food Eng. Biotechnol., 9: 92-96.
Direct Link - Lester, P.J., H.M. Thistlewood and R. Harmsen, 2000. Some effects of pre-release host-plant on the biological control of Panonychus ulmi by the predatory mite Amblyseius fallacies. Exp. Applied Acarol., 24: 19-33.
PubMed - Mohaghegh, J., P. de Clercq and L. Tirry, 2001. Functional response of the predators Podisus maculiventris (say) and Podisus nigrispinus (Dallas) (Heteroptera: Pentatomidae) to the beet armyworm, Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae): Effect of temperature. J. Applied Ent., 125: 131-134.
Direct Link - Pervez, A. and Omkar, 2005. Functional responses of coccinellid predators: An illustration of a logistic approach. J. Insect Sci., 5: 5-5.
PubMedDirect Link - Reitz, S.R., J.E. Funderburk and S.M. Waring, 2006. Differential predation by the generalist predator Orius insidiosus on congeneric species of thrips that vary in size and behaviour. Entomol. Exp. Appl., 119: 179-188.
CrossRef - Riudavents. J. and C. Castane, 1998. Identification and evaluation of native predators of Frankliniella occidentalis (Thysanoptera: Thripidae) in the maditerranean. Envir. Entomol., 27: 86-93.
Direct Link - Rosenheim, J.N., 2005. Intraguild predation of Orius tristicolor by Geocoris spp. and the paradox of irruptive spider mite dynamics in California cotton. Biol. Control, 32: 172-179.
CrossRef - Salim, M., S.A. Masud and A.M. Khan, 1987. Orius albidipennis (Reut.) (Hemiptera: Anthocoridae), a predator of cotton pests. Philipp. Entomol., 7: 37-42.
Direct Link - Sobhy, I.S., A.A. Sarhan, A.A. Shoukry, G.A. El-Kady, N.S. Mandour and S.R. Reitz, 2010. Development, consumption rates and reproductive biology of Orius albidipennis reared on various prey. BioControl, 55: 753-765.
CrossRef - Solomon, M.E., 1949. The natural control of animal populations. J. Anim. Ecol., 18: 1-35.
CrossRefDirect Link - Takafuji, A., A. Ozawa, H. Nemoto and T. Gotoh, 2000. Spider mites of Japan: Their biology and control. Exp. Applied Acarol., 24: 319-335.
CrossRef - Tully, T., P. Cassey and R. Ferriere, 2005. Functional response: Rigorous estimation and sensitivity to genetic variation in prey. Oikos, 111: 479-487.
CrossRef - Vacante, V., G.E. Cocuzza, P. de Clercq, M. van de Veire and L. Tirry, 1997. Development and survival of Orius albidipennis and O. Laevigatus (Het.: Anthocoridae) on various diets. Entomophaga, 42: 493-498.
Direct Link - Veeravel, R. and P. Baskaran, 1997. Functional and numerical responses of Coccinella transversalis Fab. and Cheilomenes sexmaculatus Fab. feeding on the melon aphid, Aphis gossypii Glov. Insect Sci. Applied, 17: 335-339.
CrossRefDirect Link - Wittmann, E.J. and S.R. Leather, 1997. Compatibility of Orius laevigatus Fieber (Hemiptera: Anthocoridae) with Neoseiulus (Amblyseius) cucumeris Oudemans (Acari: Phytoseiidae) and Iphiseius (Amblyseius) degenerans Berlese (Acari: Phytoseiidae) in the biocontrol of Frankliniella occidentalis Pergande (Thysanoptera: Thripidae). Exper. Applied Acarol., 21: 523-538.
CrossRef - Wright, B., 1994. Know your friends: Minute pirate bugs. Midwest Biol. Control News Online, Vol. 1, No. 1.
Direct Link