
A. Jeyasankar
Department of Zoology, Arignar Anna Government Arts College, Musiri-621 201, Tamil Nadu, India
N. Raja
Department of Biology, Faculty of Natural and Computational Sciences, Post Box 196, University of Gondar, Gondar, Ethiopia
S. Ignacimuthu
Entomology Research Institute, Loyola College, Chennai 600 034, India
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 4 | Page No.: 231-238







ABSTRACT
Impact of new crystal compound 2, 5-diacetoxy-2-benzyl-4, 4, 6, 6-tetramethyl-1, 3-cyclohexanedione on feeding physiology, developmental indices and quantitative changes in haemolymph protein on the larvae of armyworm, Spodoptera litura was studied. The novel compound was isolated from the leaves of medicinal plant, Syzygium lineare. The percentage of feeding physiology parameter such as Consumption Index (CI), Growth Rate (GR), Approximate Digestibility (AD), Efficiency of Conversion of Digested Food (ECD) and Efficiency of Conversion of Ingested Food (ECI) and weight gained by the larva was decreased significantly (p<0.05 by LSD). The developmental indices such as time taken for pupation, pupal weight, adult longevity, eggs laid per moths and the percentage of egg hatchability were also decreased significantly (p<0.05 by LSD) compared to control. Statistically significant (p<0.05 by LSD) reduction in total haemolymph protein was recorded at 100 ppm concentration of crystal compound treated larvae. This compound has potential to serve as an alternative botanical pesticide in the management of agricultural important filed insect pests.
PDF Abstract XML References Citation
Received: November 16, 2011;
Accepted: February 16, 2012;
Published: March 06, 2012
How to cite this article
A. Jeyasankar, N. Raja and S. Ignacimuthu, 2012. Impact of Novel Crystal Compound 2, 5-diacetoxy-2-benzyl-4,4,6,6-tetramethyl-1,3- Cyclohexanedione on Feeding Physiology and Developmental Indices of Armyworm, Spodoptera litura Fab. (Lepidoptera: Noctuidae). Journal of Entomology, 9: 231-238.
DOI: 10.3923/je.2012.231.238
URL: https://scialert.net/abstract/?doi=je.2012.231.238
DOI: 10.3923/je.2012.231.238
URL: https://scialert.net/abstract/?doi=je.2012.231.238
INTRODUCTION
Syzygium lineare Wall (Myrtaceae) is a shrub or small tree with white flowers, linear leaves, thick bark and elongated, slender pedicles found in river banks of Tirunelveli hills of Western Ghats, Tamil Nadu, India (Manickam et al., 2004). The leaf powder is used for body cooling and the paste of the fruit is used for increasing stamina. In addition it is used to treat fungal and bacterial infection. The bark of the plant is having astringent, refrigerant and diuretic properties (Duraipandiyan et al., 2008). The antifeedant and growth inhibitory activities of various crude extracts and purified fractions of the plant were evaluated well against economically important polyphagous pest Spodoptera litura (Jeyasankar et al., 2010). Recently, characterization of novel crystal compound and their insecticidal activities against Spodoptera litura was also reported (Jeyasankar et al., 2011). The Department of Science and Technology, New Delhi, India provide the patent right for the process of novel crystal compound isolation (Ignacimuthu et al., 2004; patent No. 236541). There is no other further work in this crystal compound so far. Therefore, in this study impact of crystal compound on feeding physiology, developmental indices and quantitative changes in haemolymph protein of S. litura was reported for the first time.
MATERIALS AND METHODS
Plant materials: Fresh leaves of Syzygium lineare were collected from Kalakad Mundanthurai Tiger Reserve Forest, Western Ghats of Tamil Nadu, India. Plant nomenclature was confirmed by plant taxonomist Dr. D. Narasimhan, Department of Botany, Madras Christian College, Chennai, India. The voucher specimen (MPH No. 170) was prepared and deposited at the Entomology Research Institute, Loyola College, Chennai, India.
Extraction and isolation: Two kilo grams of leaf powder was soaked in ethyl acetate for 48 h and extracted by following the method described earlier (Baskar et al., 2009). The residue of the extract was isolated by filtering through a Buchner Funnel with Whatman number 1 filter paper. The solvent from the filtrate was evaporated by using rotary vacuum evaporator. The fraction from the crude ethyl acetate extract (30 g) was separated by using silica gel (500 g-acme’s 100-200 mesh) column chromatography, eluted with n-hexane-ethyl acetate (30:70) and allowed for the crystal formation. The crystal compound structure was confirmed by XRD crystallographic data.
Experiment on feeding physiology: The second generation larvae of S. litura were collected from the insectory at Entomology Research Institute, Loyola College, Chennai, Tamil Nadu, India. The impact of crystal compound on the developmental physiology and food utilization characters of S. litura was evaluated at optimum concentration of 50 ppm and computed by gravimetric method (Waldbauer, 1968). The castor leaves were treated with the optimum concentration of crystal compound and 10 numbers of IV instar larvae was released to feed on 24 h. After that the larvae were transferred to normal castor leaves and reared up to VI instar stage. Acetone and water treated leaves was considered as control and pure azadirachtin (50 ppm) was used as a standard. The body weight gained by the larva was estimated by subtracting the initial weight of the larva from the final weight and the remaining feeding physiology parameters was calculated by using the following formula:
| (1) |
Where:
| F | = | Weight of leaf eaten |
| A | = | Mean wt. of larva during the feeding period |
| T | = | Duration of the feeding period |
| (2) |
| (3) |
| (4) |
| (5) |
Experiment on developmental indices: The developmental indices such as, time taken for pupation, pupal duration, pupal weight, adult longevity, fecundity and egg hatchability was studied on S. litura. The castor leaves were treated with crystal compound at 50 ppm concentration and released 10 larvae of IVth instar and allowed to feed for 24 h. After that larvae were transferred to fresh castor leaves and reared up to adult emergence and egg lying of newly emerged adults. The various developmental indices was recorded and calculated by the method of Sen and Sehgal (1993). Acetone and water treated leaves were considered as control and pure azadirachtin (50 ppm) was used as a standard. The experiment was replicated five times and the data was subjected to Least Significant Difference test (LSD p<0.05) for statistical significant.
Quantitative estimation of haemolymph protein: The impact of crystal compound on haemolymph protein was studied in S. litura. In this experiment fourth instar larvae (10 for each concentration) was released in to castor leaves treated with 25, 50, 75 and 100 ppm concentration of crystal compound and allowed to feed for 24 h and then transferred to normal leaves. The haemolymph was collected from 10 larvae for each treatment separately three days after treatment. The haemolymph was drawn by pricking the second proleg of the larva with sterilized needle and collected into prechilled Eppendorf tubes with few crystals of phenylthiocarbamide (1-phenyl-2-thiourea). Positive and negative control was maintained as that of previous experiment. The sample was collected from three replicates, pooled and centrifuged in a refrigerated centrifuge at 10,000 rpm for 10 min at 4°C. The clear supernatant solution was collected and subjected to quantitative estimation of protein according to the method of Lowry et al. (1951) by using bovine serum albumin as standard.
RESULTS
Crystal compound isolated from the leaves of S. lineare was cyclohexanedione derivatives of 2, 5-diacetoxy-2-benzyl-4, 4, 6, 6-tetramethyl-1, 3-cyclohexanedione. The molecular formula is C21H26O6 and mass weight is 315 (m+ -CH3-C-O) base peak, 91 (C6H5CH2) or C7H7+. Details of the characterization of the crystal compound were already published by Jeyasankar et al. (2011).
The Consumption Index (CI), Growth Rate (GR), Approximate Digestibility (AD), comparative efficiency of conversion of digested (ECD) food in to body mass and comparative efficiency of conversion of ingested (ECI) food in to body mass showed statistical significant (p<0.05; by LSD) on crystal compound treatment compared to control (Table 1). The consumption index and the growth rate were decreased at the maximum of 52.7 and 61.2%, respectively in crystal compound treatment compared to control. The value of AD, ECD and ECI was observed at the maximum of 70.27, 31.24 and 20.15, respectively in positive control but when compare to crystal compound treatment the values are not statistically significant (p>0.05; by LSD).
| Table 1: | Effect of crystal compound of S. lineare on feeding physiology of S. litura |
 | |
| Values are mean and standard deviation of five replications. Similar alphabets within the column are statistically not significant (p>0.05 by LSD) | |
| Table 2: | Effect crystal compound of S. lineare on development physiology of S. litura |
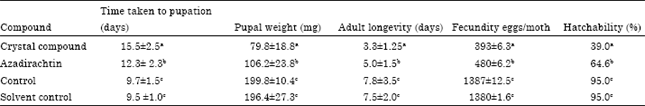 | |
| Values are mean and standard deviation of five replications. Similar alphabets within the column are statistically not significant (p>0.05 by LSD) | |
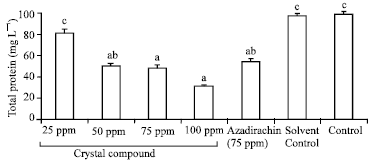 | |
| Fig. 1: | Effect of crystal compound of S. lineare on haemolymph protein of fourth instar larvae of S. litura Values are mean of five replications. Similar alphabets are statistically not significant (p>0.05 by LSD) |
The reduction of larval weight in crystal compound treatment was 53.9% and the azadirachtin treatment was 41.9% which is statistically significant at p<0.05 by LSD. All the parameters in solvent and aqueous treatment did not show any statistical significant.
In various developmental indices (Table 2), time taken for the completion of pupation was increased at 15.5 days in crystal compound treatment compared to control and the result was statistically significant (p<0.05; LSD). The significant reduction in pupal weight (79.8), adult longevity (3.3 days), fecundity (393 eggs/month) and percentage of egg hatchability (39%) was recorded in crystal compound treatment.
Total haemolymph protein content of S. litura was 99 99.8 mg mL-1, respectively in solvent control and control (Fig. 1). The results were statistical not significant (p>0.05; by LSD). However, crystal compound at 100 ppm showed statistically significant (p<0.05; by LSD) drastic reduction (70%) compared to control.
DISCUSSION
Decreased consumption index in crystal compound treatment was adversely affecting the growth rate of the larvae. Similar results have been reported earlier by Prasad (1973), Ramdev and Rao (1979), Prabhakar (1991), Chitra and Rao (1996) and Kubo and Klocke (1993). The increased rate of AD values reflects that the insect attempts to compensate reduced consumption and utilization of treated diet in order to maintain the growth (Blaney et al., 1990). Barnby and Klocke (1987) reported that the value may have increased in response to a decreased rate of passage of food through the gut was shown to occur in H. virescens and also in the cabbage webworm, Crocidolomia binotalis (Fagoonee and Lange, 1981). It is reasonable to associate such increase in AD of treated insects to compensate for their reduced food consumption and utilization efficiencies to meet energy requirements to maintain growth and developmental processes (Tanzubil, 1989; Tanzubil and McCaffery, 1990).
In crystal compound treatment the larvae consumed less food and ultimately gained less body weight and also decreased the efficiency in converting ingested and digested food into biomass (ECI and ECD). Furthermore, negative ECI and ECD values might be due to the adverse effect on the hormonal system, which controls the utilization of food. These results are corroborate to the earlier reports on the larvae of H. virescens (Barnby and Klocke, 1987) and Eligma narcissus indica (Joseph, 2000) and the larvae of African armyworm, S. exempta (Tanzubil, 1995). Fagoonee, (1983) also suggested that the high ECD and ECI values reflect compensation by the larvae since they were consuming much less food than the larvae exposed to non-treated leaf discs. This high utilization efficiency declined presumably because the detoxification mechanisms were exhausted. A similar mechanism of diversion of energy from the production of biomass to detoxification could account for the low utilization efficiencies in H. armigera (Barnby and Klocke, 1987). Sahayaraj (1998) also reported that significantly negative ECI and ECD values in plant extracts treated categories and this indicated that the utilization of food for conservation of energy for various life activities was challenged by the chemicals present in the plant extracts. Barnby and Klocke (1987) reported that when 0.25 μg of azadirachtin was introduced into the gut of Heliothis virescens larvae, they gained weight for the first two days and subsequently lost their weight prior to pupation. The inhibition of larval development resulted at least in part, from a reduced physiological age brought about by reduced food intake and weight gain as it has been suggested by many workers (Arnason et al., 1987; Rembold et al., 1982; Ladd et al., 1984).
The time taken for the pupation and pupal duration was increased in the treatment of crystal compound. It is due to the effect of active compound on the juvenile hormone thereby arresting the development ultimately the larvae take more time for pupation. Due to the influence of compound on insect metamorphosis pupa will take longer time compared to control. These findings are similar with earlier reports of Slama and Williams (1966) who has reported that plant extracts/compound have effect on juvenile hormone and ecdysone thereby development was delayed. Bowers et al. (1976) also reported that plant substances disrupt the growth by antagonizing juvenile hormone action. Decreased pupal weight, adult longevity, fecundity and percent hatchability of eggs was due to reduced feeding rate of the larvae on treated plant leaves or the negative impact of plant compound on juvenile hormone and ecdysone. When the feeding rate was decreased, pupal weight was also decreased and the development of oozyte was also reduced. The number of eggs laid by adults was reduced or normal adult emergence was suppressed or sometimes adults laid eggs before copulation because of improper development of oozyte caused by the crystal compound treatment. Because of the infertile egg the hatchability was reduced. Several workers have reported that the reduced rate of pupal weight was caused due to the influence of plant substances (Fagoonee and Lange, 1981; Schmutterer et al., 1983; Mukherjee and Sharma, 1993).
Proteins constitute the basic entities in the living being which is reduced in crystal compound treatment. The reduction is depended on the concentration of plant extracts, when the concentration is increased the reduction rate also increased. It may be due to the toxic stress of active compound related to reduction of protein synthesis by deranging the protein machinery or it might be due to faster rate of proteolysis. Similar observations were reported by Bhagwan et al. (1992), Reddy et al. (1993), Krishnayya and Rao (1995), Chitra and Rao (1996) and Anitha et al. (1999). Decreased protein content due to the botanical treatment was reported by many workers (De Man et al., 1981; Jadhav and Ghule, 2003; Kaur and Rup, 2003). Ayyangar and Rao (1990) reported that haemolymph protein of S. litura was decreased due to the impact of azadirachtin. Due to reduced feeding and growth inhibitory effect of novel crystal compound 2, 5-diacetoxy-2-benzyl-4, 4, 6, 6-tetramethyl-1, 3-cyclohexanedione, feeding physiology, various developmental indices and protein concentration of S. litura was decreased. This compound may be useful to development of novel botanical pesticide to management of agricultural important insect pests.
ACKNOWLEDGMENT
The confirmation and identification of plant species by Dr. D. Narasimhan, a plant taxonomist, Department of Botany, Madras Christian College, Chennai, India was gratefully acknowledged.
REFERENCES
- Anitha, B., M. Arivalagan, S.M.S. Nalaini and G. Durairaj, 1999. Effect of alkaloid abrine, isolated from Abrus precatorius (Linn.) seeds on mealy bug Maconellicoccus hirsutus green. Ind. J. Exp. Biol., 34: 415-417.
Direct Link - Arnason, J.T., B.J.R. Philogene, N. Donsker and L. Kubo, 1987. Limonoids from Meliaceae and Rutaceae reduce feeding, growth and development of Ostrinia nubilalis. Entomol. Exp. Applicata, 43: 221-226.
CrossRef - Barnby, M.A. and J.A. Klocke, 1987. Effects of azadirachtin on the nutrition and development of the tobacco budworm, Heliothis virescens (Fabr.) (Noctuidae). J. Insect. Physiol., 33: 69-75.
CrossRefDirect Link - Baskar, K., S. Kingsley, S.E. Vendan, M.G. Paulraj, V. Duraipandiyan and S. Ignacimuthu, 2009. Antifeedant, larvicidal and pupicidal activities of Atalantia monophylla (L) correa against Helicoverpa armigera Hubner (Lepidoptera: Noctuidae). Chemosphere, 75: 355-359.
CrossRefDirect Link - Blaney, W.M., M.S.J. Simmonds, S.V. Ley, J.C. Anderson and P.L. Toogood, 1990. Antifeedant effects of Azadirachtin and structurally related compounds on lepidopterous larvae. Entomol. Exp. Et Applicata, 55: 149-160.
Direct Link - Fagoonee, I. and G. Lange, 1981. Noxious effects of neem extract on Crocidolonia bionotalis. Phytoparasitica, 9: 111-118.
CrossRefDirect Link - Jeyasankar, A., N. Raja and S. Ignacimuthu, 2010. Antifeedant and growth inhibitory activities of Syzygium lineare Wall (Myrtaceae) against Spodoptera litura Fab. (Lepidoptera: Noctuidae). Cur. Res. J. Biol. Sci., 2: 173-177.
Direct Link - Jeyasankar, A., N. Raja and S. Ignacimuthu, 2011. Insecticidal compound isolated from Syzygium lineare Wall. (Myrtaceae) against Spodoptera litura (Lepidoptera: Noctuidae). Soudi J. Biol. Sci., 18: 329-332.
CrossRefDirect Link - Kaur, R. and P.J. Rup, 2003. Influence of some Plant Growth Regulators (PGR) on biochemical profile in the larvae of melon fruit fly Bactrcera cucurbitae (Coquillett) (Diptera: Trypetidae). Entomon, 28: 89-96.
Direct Link - Kubo, I. and J.A. Klocke, 1993. Azadirachtin insect ecdysis inhibitor. Agri. Biol. Chem., 46: 1951-1953.
Direct Link - Ladd, T.L., J.D. Warthen and M.G. Klein, 1984. Japanese beetle (Coleoptera: Scarabaeidae): The effects of azadirachtin on the growth and development of the immature forms. J. Econ. Entomol., 77: 903-905.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Reddy, K.D., C.N. Bhagwan and K. Sukumar, 1993. Depletion of amino acids and protein in the nymphal haemolymph of the red cotton bug, Dysdercus koenigii (Fab.) (Hemiptera: Pyrrhocoridae) treated with Annona squamosa extract. Phytophaga, 5: 45-50.
Direct Link - Rembold, H., G.K. Sharma, Czoppelt and C. Schmutterer, 1982. Azadirachtin: A potent insect growth regulator of plant origin. Zeitschrift Angewandte Entomol., 93: 12-17.
CrossRef - Schmutterer, H., R.C. Saxena and J. von der Hyke, 1983. Morphogenetic effects of some partially-purified fractions and methanolic extracts of neem seeds on Mythimna separata (Walker) and Cnaphalocrocis medinalis (Guenee). J. Applied Entomol., 95: 230-237.
CrossRefDirect Link - Slama, K. and C.M. Williams, 1966. Paper factor as an inhibitor of the embryonic development of the European bug, Pyrrhocoris apterous. Nature, 210: 329-330.
CrossRefDirect Link - Tanzubil, P.B. and A.R. McCaffery, 1990. Effects of azadirachtin and aqueous neem seed extracts on survival, growth and development of the African armyworm, Spodoptera exempta. Crop Prot., 9: 383-386.
CrossRefDirect Link - Tanzubil, P.B., 1995. Effects of neem (Azadirachta indica A. Juss) extract on food intake and utilization in the African armyworm, Spodoptera exempta. Int. J. Trop. Insect Sci., 16: 167-170.
CrossRefDirect Link
savaliya dinesh Reply
how to calculate the relative growth rate.?
pls give me immediately