
Dinesh Kumar
Silkworm Physiology Laboratory, Central Tasar Research and Training Institute (Central Silk Board), P.O. Piska-Nagri, Ranchi 835303, India
J.P. Pandey
Silkworm Physiology Laboratory, Central Tasar Research and Training Institute (Central Silk Board), P.O. Piska-Nagri, Ranchi 835303, India
Ragini
Silkworm Physiology Laboratory, Central Tasar Research and Training Institute (Central Silk Board), P.O. Piska-Nagri, Ranchi 835303, India
A.K. Sinha
Silkworm Physiology Laboratory, Central Tasar Research and Training Institute (Central Silk Board), P.O. Piska-Nagri, Ranchi 835303, India
B.C. Prasad
Silkworm Physiology Laboratory, Central Tasar Research and Training Institute (Central Silk Board), P.O. Piska-Nagri, Ranchi 835303, India
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 4 | Page No.: 220-230







ABSTRACT
The tropical tasar silkworm, Antheraea mylitta Drury is sericigenous wild insect has own dictation on its life cycle stages. Being a poikilothermic organism, temperature decides the fate of A. mylitta during embryonic and postembryonic development. Silkworm eggs (seed) is very vital input of tasar silk industry but due to global warming elevation in temperature with less humidity during 1st crop grainage of A. mylitta negatively affects the quality and quantity of egg hatching. In the present study, impact of temperature stress on embryonic development and biochemical profile of A. mylitta eggs has been investigated. Data reveals that considerable alteration in temperature used to hamper the protein and carbohydrate profile which leads to affect the embryonic development and hatching of eggs. Fluctuation in temperature with low humidity causes delay in egg hatching and decrease in hatching percentage. But higher temperature with low humidity during embryonic development of eggs resulted in death of embryo during early age. The depressed eggs with fully formed dead larvae inside the eggs were found after high temperature stress. The concentration of the protein and carbohydrate steadily decreases during subsequent larval differentiation until hatching. Protein and carbohydrate profile also confirm the death of embryo during early stage. In un-hatched eggs, due to less metabolized utilization higher concentration of these nutrients are available. Our initial information indicates that protein and carbohydrate profile can be utilized as biochemical marker for testing appropriate embryonic development and hatchability of eggs.
PDF Abstract XML References Citation
Received: November 03, 2011;
Accepted: January 24, 2012;
Published: March 06, 2012
How to cite this article
Dinesh Kumar, J.P. Pandey, Ragini, A.K. Sinha and B.C. Prasad, 2012. Temperature Discerns Fate of Antheraea mylitta Drury Eggs During Embryonic Development. Journal of Entomology, 9: 220-230.
DOI: 10.3923/je.2012.220.230
URL: https://scialert.net/abstract/?doi=je.2012.220.230
DOI: 10.3923/je.2012.220.230
URL: https://scialert.net/abstract/?doi=je.2012.220.230
INTRODUCTION
Due to global warming, impact of temperature on living organisms has been core of scientific investigation in recent past. In insect, all species has its own choice of temperatures for its usual growth and very high temperature slows down the growth, may leads to developmental malfunction, such as larval ecdysis and adult emergence (Chapman, 2002). Being a poikilothermic organism, temperature decides the fate of insect, during embryonic and postembryonic development. It is well understood that, temperature affects the insect life-cycle plasticity, physiological thermal traits, immune responses and gene expression (Neven, 2000; Kiuchi et al., 2008; Arias et al., 2011; Beckett, 2011; Catalan et al., 2011). It also affects the metabolic rate and oxidative stress status (Contreras and Bradley, 2010; Lalouette et al., 2011). Woods and Bonnecaze (2006) studied the impact of temperature, water and oxygen on insect eggs at a transition between diffusion and reaction limitation. Even oxygen consumption profiles of insect cells get affected due to temperature (Gotoh et al., 2004). In a number of insect species, the growth rate is improved and the developmental period and immune cells (hemocytes) are abridged as the temperature raises (Wigglesworth, 1972; Kiuchi et al., 2008; Pandey et al., 2010).
In recent past, several physiological studied have been undertaken in different aspects of tropical tasar silkworm, Antheraea mylitta (Kumar et al., 2011; Pandey et al., 2010, 2011, 2012). Although, embryonic development of insects in relation to various stresses have been studied by several researchers (Khattak et al., 2001; Duraimurugan and Regupathy, 2005; Temerak, 2006; Ali and Rizvi, 2007; Seidavi et al., 2008; Ahmad et al., 2008; Sujatha and Joseph, 2011; Bajaj and Sharma, 2011) but impact of temperature in relation to embryonic development of tasar silkworm eggs (seed) is not studied much which is backbone of tasar silk industry.
The tropical tasar silkworm, A. mylitta is produces tasar silk of commercial importance at industry level and it gives direct supplementary livelihood to poor people of the society. The basic reproductive unit in the tasar silk industry is the silkworm eggs and temperature plays very crucial role during embryonic development. Embryonic development starts within few hours of egg lying and it needs proper incubation and care. It reported that, as the development of the embryo proceeds, both lipids and carbohydrates gets utilized, resulting in increase in the concentration of citrate, pyruvate and lactate (Pant et al., 1979). Due to global warming elevation in temperature with less humidity was recorded during 1st crop grainage which not only affect the egg hatching but also adversely affects the seed sector of tasar industry which is very essential input of sericulture. Although, embryology of several sericigenous insects have been studied by researchers (Tazima, 1978; Jolly et al., 1974; Kumar et al., 1998). But biochemical and morphological changes occurs in tasar silkworm eggs under temperature stress conditions are not understood well. Hence, the present study has been conducted to investigate impact of temperature stress on embryonic development and biochemical profile of the A. mylitta eggs.
MATERIALS AND METHODS
Insect culture: Stock of tropical tasar silkworm, A. mylitta Drury (Daba ecorace) was maintained in outdoor condition and larvae were fed on fresh leaves of Asan, Terminalia tomentosa W&A. Disease free laying was obtained from our laboratory grainage and utilized for different set of experiments (Kumar et al., 2011). The 72 h old eggs were used for hatching percentage experiment. For embryonic development study 24 h old eggs were used. Present research work was carried out during the year 2010-2011 in Silkworm Physiology Laboratory, Central Tasar Research and Training Institute, Ranchi, India. Our rearing farm and laboratory are located at Latitude-23°.21’N, Longitude- 85°.20’E and an altitude of 652 m MSL, in the state of Jharkhand, India (Kumar et al., 2011).
Embryonic development: For proper embryonic development, tasar silkworm eggs were incubated after standardisation, at optimum temperature and relative humidity i.e., 26-28°C and 75-80%, respectively. Different age groups of samples were collected daily for embryonic development and biochemical study.
Temperature stresses: To study the impact of temperature stresses on hatching of tasar silkworm eggs, 200 eggs in five replications were kept in egg hatching boxes having transparent top to permit light and perforated sides for aeration (5.5x4x2 cm) at four temperature stress conditions (T1 to T4) along with control (T5). For treatment-1 (T1) temperature 37°C and 40% RH in Vista Biocell environmental chamber incubator (This condition was observed during first crop grainage), for treatment-2 (T2) temperature 27°C and 60% RH in Vista Biocell environmental chamber followed by one higher temperature stress at 37°C daily, for treatment-3 (T3) temperature at 25°C and 60-70% RH at room temperature and treatment-4 (T4) cyclic temperature 38 and 18°C and 50-85% RH under outdoor direct sun light covered with polythene sheet. In treatment-5 (T5) respective control were maintained and considered as controls. After completion of treatment the eggs samples of experimental and control groups were kept at -80°C for biochemical analysis. Hatching percentage was recorded after 1st, 2nd and 3rd day separately by using following formula:
Eggs samples collection: Eggs samples of different treatments were taken in Eppendorf tube and crushed and homogenized in 1 mL, 50 mm phosphate buffer (pH 6.9). These samples were centrifuged at 5000 rpm for 10 min and supernatant was collected in to pre-chilled Eppendorf tube and utilized for present study.
Estimation of protein and carbohydrate in eggs: Samples were collected daily for biochemical study. Eggs samples of different temperature stress conditions and control were analyzed for protein and carbohydrate after third day of egg hatching. The protein estimation was carried according to Bradford’s micro protein assay (Bradford, 1976) with slight modifications. Concentration of the protein sample was determined from a standard curve drawn using bovine serum albumin and OD was taken at 595 nm. Colorimetric determination of carbohydrate was performed by using phenol-sulfuric acid based reaction absorbance at 490 nm to see colored aromatic complex formed between phenol and the carbohydrate (Fournier, 2001). UV 5704-SS spectrophotometer was used in the present experiment.
Microscopic observation: For proper embryonic development, eggs were incubated at optimum temperature and relative humidity i.e., 26-28°C and 75-80%, respectively. Different temperature stress was given to eggs and samples were collected daily for microscopic observation. To see the impact of temperature stress on embryonic development, eggs samples of different age groups of control and experimental were dissected out by opening the eggs shell and subjected to microscopic morphological observation by using stereo zoom microscope. Comparative morphological evaluation of pictures was done and temperature stress impact on embryonic development was analysed.
Data analysis: The data was subjected to the statistical analysis by using ANOVA. The Microsoft Excel 2003 software was used for data analysis assistance.
RESULTS
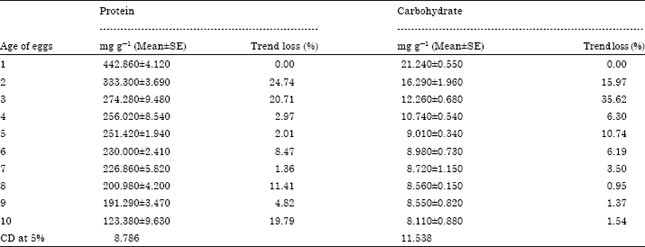
Temporal changes in protein and carbohydrate profile of A. mylitta eggs: Throughout embryonic development of egg, the concentration of protein was recorded maximum (442.86±4.120 mg g-1) on day 1 and its level decreases on advancement of age of eggs or progression of embryonic development up to 123.380±9.630 mg g-1. The utilization of protein was observed highest on day 10 (35.50%) followed by day 2 (24.74%), day 3 (17.71%) and day 8 (11.41%). The utilization of protein was observed lowest on day 5 (1.80%) followed by day 7 (1.36%) and day 1 (0.00%). Regression equation shows Y = -26.332x+397.87 and regression trend shows R2 = 0.8476.
During embryonic development of egg, the concentration of carbohydrate was recorded maximum (21.240 mg g-1) on day 1 and its level decreases on advancement of age of eggs or progression of embryonic development up to 8.110±0.880 mg g-1. The utilization of carbohydrate was observed highest on day 3 (24.73%) followed by day 2 (23.27%), day 5 (16.06%) and day 4 (12.45%). Whereas, the utilization of carbohydrate was observed lowest on day 7 (2.93%) followed by day 8 (1.81%), day 6 (0.35%) and day 9 (0.16%). (y = -26.332x+397.87 and R2 = 0.8476 (Table 1).
Impact of temperature stress on protein concentration: The concentration of total protein in eggs kept in different temperature stress conditions reveals that differences were highly significant among all treatments (p<0.1). Total protein concentration was higher in T1 (230.62±5.65 mg g-1) followed by 197.10±3.40 mg g-1 in T4 whereas, lowest ranged from 163.31±4.85 to 171.47±5.30 mg g-1 in T2, T3 and T5. It was evidence that higher concentration of protein in T1 temperature stress condition due to unfavourable condition for embryonic development and block the embryogenesis and not utilization of protein (Fig. 1). For incubation of tasar eggs, 26-28°C temperature and 75-80% RH is found appropriate to acquire proper embryonic development as well as uniform and timely hatching.
Impact of temperature stress carbohydrate concentration: Total carbohydrate concentration in eggs of different temperature stress conditions is presented in Fig. 2. The level of carbohydrate was observed significantly higher in T1 (p<0.1) than other conditions. The maximum concentration of carbohydrate in eggs kept in temperature stress condition of T1 (103.51±11.27 mg g-1) followed by T4 (72.22±5.12 mg g-1) and in T2, T3 and T5 it’s ranged between 48.67±0.83 to 55.94±4.24 mg g-1 (Fig. 2).
| Table 1: | Temporal changes in protein and carbohydrate profile of A. mylitta eggs during embryonic development |
 | |
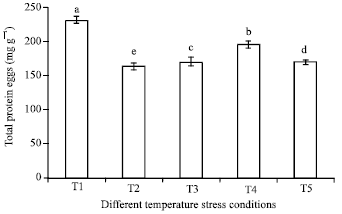 | |
| Fig. 1: | Protein profile of A. mylitta eggs kept in different temperature stress conditions. Mean followed by different alphabets are significantly different at p<0.1 |
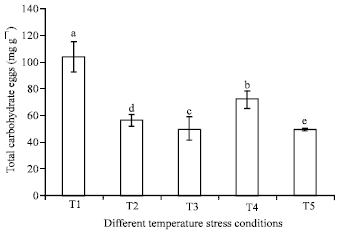 | |
| Fig. 2: | Carbohydrate profile of A. mylitta eggs kept in different temperature stress conditions. Mean followed by different alphabets are significantly different at p<0.1 |
Temporal changes during embryonic development of normal eggs: The eggs of A. mylitta contain yolk and cleavage takes place in superficial nature. Chronological variation during embryonic development of normal eggs was found from 24 h to till eggs hatching (Fig. 3a-h). Within 6 to 10 h of egg deposition after repeated division, cleavage energids originate as aggregated cytoplasm of the cytoplasmic reticulum. During 10-16 h as a result of increased cell division in ventral region the ventral plate is formed and by 22 h trough shaped embryonic primodium floats in yolk and the protocephalon and protocorn are separated by transverse furrows. Throughout 22-28 h, the germ band becomes slender and elongated and by 28-34 h along the mid portion of the germ band on upper side a long narrow depression called the primitive groove or streak or median plate is formed. By 24-36 h the development of embryo was well differentiated into head and trunk region. After 48 h, segmentation of the body is clearly visible and in 72 h embryo showed three distinct regions of the body i.e., head, thorax and abdomen were well differentiated. After 96 h the embryo became C-shaped. Although, mouthparts are yet not developed completely but they were at the advanced stage of development. The thoracic region is clearly divisible into three thoracic segments. Interestingly after 120 h organogenesis in embryo takes place. The antennae bear segments and head region was detachable into three segments. The head capsule formation completed in after 144 h and mouthparts got matured. Three segmented antennae with antennal setae, the mandibles and labrum are well developed.
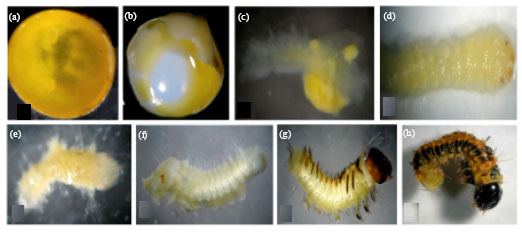 | |
| Fig. 3(a-h): | Temporal morphological changes during embryonic development of A. mylitta eggs, (a) 24 h old embryo, (b) 48 h old embryo, (c) 72 h old embryo, (d) 96 h old embryo, (e) 120 h old embryo, (f) 144 h old embryo, (g) 168 h old embryo and (h) 192 h old embryo |
After 168 h old embryo well pigmented integument were found and coloration of head capsule takes place. After 192 h embryo was fully formed larva, which is ready to cut off the egg shell or to hatch out.
Impact of temperature stress on embryonic development: For incubation of tasar eggs, 26-28°C temperature and 75-80% RH is found appropriate to acquire proper embryonic development. Marked variation in temperature and humidity negatively affects the embryonic development of tasar silkworm eggs (Fig. 4a-f). Data reveals considerable alteration in temperature used to hamper the protein and carbohydrate profile which leads to affect the embryonic development hatching of eggs. The depressed eggs with fully formed dead larvae inside the eggs were found after high temperature stress (Fig. 4a, b). But higher temperature with low humidity during embryonic development of eggs resulted in death of embryo during early age (Fig. 4c-f).
Impact of temperature stress on hatching percentage: When stress was given to tasar silkworm eggs, it showed great deal of variation in hatching percentage (Table 2). On the 1st day-maximum hatching percentage was recorded in T5 (63.50) followed by T2 (42.50) and T3 (27.67). Highly significance different in hatching percentage was observed between T2 (42.50) and T3 (27.67); between T2 (42.50) and T5 (63.50); between T3 (27.67) and T5 (63.50). On the 2nd day-maximum hatching percentage was observed in T3 (42.00) followed by T2 (30.67); T4 (38.33) and T5 (29.50) shows less hatching percentage. Significant differense in hatching percentage was also observed between T2 (30.67) and T3 (42.00); between T2 (30.67) and T4 (38.33); between T3 (42.00) and T5 (29.50); between T4 (38.33) and T5 (29.50). No significant difference was observed between T2 (30.67) and T5 (29.50).
In day 3, maximum hatching percentage was recorded in T4 (17.50) followed by T3 (14.00), T2 (9.33) and T5 (0.833) and T5 shows less hatching percentage (0.833). No significant difference in hatching percentage was observed between T2 (9.33) and T3 (14.00), between T3 (14.00) and T4 (17.50) but highly significance different was observed between T2 (9.33) and T5 (0.833), between T3 (14.00) and T5 (0.833) and between T4 (17.50) and T5 (0.833).
 | |
| Fig. 4(a-f): | Impact of temperature stress on embryonic development of A. mylitta eggs (a-b) Fresh and depressed eggs, (c-f) Larvae died inside egg shell due to higher temperature stress |
| Table 2: | Impact of temperature stress on A. mylitta egg hatching |
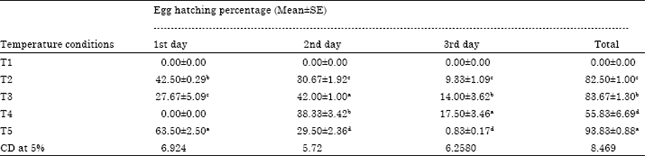 | |
| Note: T1: Temperature at 37°C and 40% RH in incubator (This condition was observed during first crop grainage), T2: Temperature at 27°C ant 60% RH in incubator followed by one higher temperature stress at 37°C daily, T3: Temperature at 25°C and 60-70% RH at room, T4: Max. temperature. 38°C and mini. temperature. 18°C and 50-85% RH under outdoor direct sun light covered with polythene sheet, T5: Control: Mean followed by different letters with in column are significantly different at p<0.1 | |
In total, maximum hatching percentage was recorded in T5 (93.83) followed by T2 (82.50) and T3 (83.67) whereas, T4 shows less hatching percentage (55.83). No significant difference in hatching percentage was observed between T2 (82.50) and T3 (83.67), between T2 (82.50) and T5 (93.83), between T3 (83.67) and T5 (93.83), but highly significant difference was observed between T2 (82.50) and T4 (55.83), between T3 (83.67) and T4 (55.83). It is observed that high temperature and less humidity are not good for larval hatching (Table 2).
DISCUSSION
In insect, all species has its own choice of temperatures for its usual growth and very high temperature slows down the growth, may leads to developmental malfunction, such as larval ecdysis and adult emergence (Chapman, 2002). Being a poikilothermic organism, temperature discerns the fate of A. mylitta, during embryonic and postembryonic development. It is reported that temperature affects the insect life-cycle plasticity, physiological thermal traits, immune responses and gene expression (Neven, 2000; Arias et al., 2011; Beckett, 2011; Ma and Ma, 2011). It also affects the metabolic rate and oxidative stress status (Contreras and Bradley, 2010; Lalouette et al., 2011). Temperature, water and oxygen impact on insect eggs have been also reported (Woods and Bonnecaze, 2006). Proper incubation of eggs plays a vital role in during embryonic development of silkworm leading to timely hatching. Based on our biochemical and morphological evidences, the appropriate condition (26-28°C temperature and 75-80% relative humidity) for incubation A. mylitta eggs has been worked out. Our finding corroborates the result several investigators i.e., Irie and Yamashita (1980) reported changes in the amounts of vitellin and other yolk proteins of the Bombyx mori eggs during embryonic development by using polyacrylamide gel electrophoresis and immuno-titration techniques. It is found that yolk proteins of the silkworm, B. mori are important source of nutrition and energy during embryo development. Xin-Pei et al. (2005) stated that the silkworm embryo does not utilize yolk proteins selectively by absorbing one or some special proteins, but by absorbing the yolk protein grains one by one during development. The role of temperature and relative humidity on oviposition and incubation of eggs was reported by several workers (Ahsan et al., 1974; Yamaoka and Hirao, 1981; Katti et al., 2007). In a number of insect species, the growth rate is improved and the developmental period and immune cells (hemocytes) are abridged as the temperature raises (Wigglesworth, 1972; Kiuchi et al., 2008; Pandey et al., 2010). Developmental response and fertility under various conditions have studied in different insects (Ali and Rizvi, 2007; Ahmad et al., 2008). Its relation with endocrine causes of early embryonic death has been also studied (Bajaj and Sharma, 2011). Likewise, impact of insecticides on embryonic development of Helicoverpa armigera (Duraimurugan and Regupathy, 2005) and its comparative effects on maize weevil, Sitophilus zeamais is also investigated (Khattak et al., 2001).
Present finding indicates that, the embryonic development starts within few hours of egg laying and it requires proper incubation. Any change in temperature can hamper the development, hatching and rearing performance. There was no hatching in temperature stress condition T1 (37°C and 40% RH), because high temperature and low humidity were unfavorable condition for embryonic development. Due to global warming this type of condition prevails during first crop grainage (June-July) of A. mylitta. Comparatively less reduction in hatching was recorded at 27°C and 60% RH in comparison to one hour higher temperature stress at 37°C daily (T2) and 25°C and 60-70% RH at room (T3). Hatching was noticed up to 55 and 40-45% depressed eggs at 18- 38°C and 50-85% RH under outdoor direct sun light covered with polythene sheet (T4) and hatching was above 90% Control (T5). The synchronization on hatching has been also worked out (Ahsan et al., 1974). Recently (Seidavi et al., 2008) impact of individual selection in 3P generation on resistance, production and reproduction improvement of three Japanese pure lines of silkworm Bombyx mori have been observed. Impact of oligidic diet and its effects of mass rearing performance on Spodoptera litura (Fab.) have also made (Sujatha and Joseph, 2011) and role of moth scales remain on the top of egg masses of Spodoptera littoralis and it’s interference with the performance of the natural bio-product was found (Temerak, 2006).
Total protein and carbohydrate concentration was very high in temperature stress condition of T1. It is clearly substantiate that non-utilization of protein and carbohydrate for development of embryo due to fatality. So it was concluded that T1 condition is lethal for embryonic development. It is reported that the as development of the embryo proceeds, carbohydrates were metabolized during early embryogenesis, lipids were catabolised at the later stages (Pant et al., 1979). Temporal changes in embryonic development in A. mylitta eggs were similar as earlier reported by Kumar et al., (1998). The protein and carbohydrate profile of eggs of A. mylitta during embryonic development from 1st day to 8th day showed utilization of protein and carbohydrate whereas, peak of trend during early and late phase of embryonic development. (Pant et al., 1979) also reported a significant increase in both total carbohydrates and glycogen on days 5 and 6 and suggested the concurrent occurrence of both gluconeogenesis and glycogenesis. As the development of the embryo proceeds, both lipids and carbohydrates were utilized, resulting in the increase in the concentration of citrate, pyruvate and lactate. Irie and Yamashita (1980) found changes in the amounts of vitellin and other yolk proteins of the eggs of the silkworm, Bombyx mori during embryonic development using polyacrylamide gel electrophoresis and immunotitration techniques. In addition Xin-Pei et al. (2005) also reported that the yolk proteins of the silkworm, B. mori are important sources of nutrition and energy during embryo development.
CONCLUSIONS
Temperature discerns the fate of A. mylitta eggs during embryonic development. Higher temperature and low humidity during embryonic development leads to death of embryo during early age. Protein and carbohydrate profile also affected after treatment. Temperature stress caused delay and poor hatching, depression of eggs and death of fully formed larvae inside egg. Our initial finding indicated the possible application protein and carbohydrate profile as biochemical marker for testing of eggs to ascertain proper embryonic development and hatchability. In addition, it will also helping in finding the reason for egg depression and seed quality in future. For development of technology to test eggs, further refinement and gel electrophoresis study is essential.
ACKNOWLEDGMENTS
Authors are thankful to training section of the Institute for suggestion and support during experiments.
REFERENCES
- Ahmad, S.K., A. Ali and P.Q. Rizvi, 2008. Influence of varying temperature on the development and fertility of Plutella xylostella (L.) (Lepidoptera: Yponomeutidae) on cabbage. Asian J. Agric. Res., 2: 25-31.
CrossRefDirect Link - Ali, A. and P.Q. Rizvi, 2007. Developmental response of cabbage butterfly, Pieris brassicae L. (Lepidoptera: Pieridae) on different cole crops under laboratory and field condition. Asian J. Plant Sci., 6: 1241-1245.
CrossRefDirect Link - Kiuchi, T., F. Aoki and M. Nagata, 2008. Effects of high temperature on the hemocyte cell cycle in silkworm larvae. J. Insect Physiol., 54: 454-461.
CrossRefDirect Link - Arias, M.B., M.J. Poupin and M.A. Lardies, 2011. Plasticity of life-cycle, physiological thermal traits and Hsp70 gene expression in an insect along the ontogeny: Effect of temperature variability. J. Thermal Biol., 36: 355-362.
CrossRefDirect Link - Bajaj, N.K. and N. Sharma, 2011. Endocrine causes of early embryonic death: An overview. Curr. Res. Dairy Sci., 3: 1-24.
CrossRefDirect Link - Beckett, S.J., 2011. Insect and mite control by manipulating temperature and moisture before and during chemical-free storage. J. Stored Prod. Res., 47: 284-292.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Catalan, T.P., A. Wozniak, H.M. Niemeyer, A.M. Kalergis and F. Bozinovic, 2011. Interplay between thermal and immune ecology: Effect of environmental temperature on insect immune response and energetic costs after an immune challenge. J. Insect Physiol.
CrossRef - Contreras, H.L. and T.J. Bradley, 2010. Transitions in insect respiratory patterns are controlled by changes in metabolic rate. J. Insect Physiol., 56: 522-528.
CrossRef - Duraimurugan, P. and A. Regupathy, 2005. Mitigation of insecticide resistance in Helicoverpa armigera (Hubner) (Lepidoptera:Noctuidae) by conjunctive use of trap crops, neem and Trichogramma chilonis ishii in cotton. Int. J. Zool. Res., 1: 53-58.
CrossRefDirect Link - Fournier, E., 2001. Colorimetric quantification of carbohydrates. Curr. Protocols Food Anal. Chem.
CrossRefDirect Link - Gotoh, T., K. Chiba and K.I. Kikuchi, 2004. Oxygen consumption profiles of Sf-9 insect cells and their culture at low temperature to circumvent oxygen starvation. Biochem. Engn. J., 17: 71-78.
CrossRef - Irie, K. and O. Yamashita, 1980. Changes in vitellin and other yolk proteins during embryonic development in the silkworm, Bombyx mori. J. Insect Physiol., 26: 811-813, 815-817.
CrossRef - Khattak, M.K., A.B. Broce and B. Dover, 2001. Comparative effects of neem or mineral oil on maize weevil, Sitophilus zeamais Motsch. and its parasitoid, Anisopteromalus calandrae (Howard). J. Biological Sci., 1: 378-381.
CrossRefDirect Link - Kumar, D., J.P. Pandey, J. Jain, P.K. Mishra and B.C. Prasad, 2011. Qualitative and quantitative changes in protein profile of various tissue of tropical tasar silkworm, Antheraea mylitta Drury. Int. J. Zool. Res., 7: 147-155.
CrossRef - Lalouette, L., C.M. Williams, F. Hervant, B.J. Sinclair and D. Renault, 2011. Metabolic rate and oxidative stress in insects exposed to low temperature thermal fluctuations. Comp. Biochem. Physiol. A. Mol. Integ. Physiol., 158: 229-234.
CrossRefPubMedDirect Link - Ma, G. and C.S. Ma, 2011. Effect of acclimation on heat-escape temperatures of two aphid species: Implications for estimating behavioural response of insects to climate warming. J. Insect Physiol., (In Press).
CrossRef - Neven, L.G., 2000. Physiological responses of insects to heat. Postharvest Biol. Technol., 21: 103-111.
CrossRef - Pandey, J.P., P.K. Mishra, D. Kumar, B.M.K. Singh and B.C. Prasad, 2010. Effect of temperature on hemocytic immune responses of tropical tasar silkworm, Antheraea mylitta D. Res. J. Immunol., 3: 169-177.
CrossRefDirect Link - Pandey, J.P., D. Kumar, S.K. Roy, P.K. Mishra, A.K. Sinha and B.C. Prasad, 2012. Hemocytes and enzyme-based route to evaluate the impact of seed cocoons preservation conditions on Antheraea mylitta pupae. Int. J. Zool. Res., 8: 1-11.
CrossRefDirect Link - Pandey, J.P., P.K. Mishra, D. Kumar, A.K. Sinha, B.C. Prasad, B.M.K. Singh and T.K. Paul, 2011. Possible-efficacy of 26 kDa Antheraea mylitta cocoonase in cocoon-cooking. Int. J. Biol. Chem., 5: 215-226.
CrossRef - Pant, R., S. Kumar and S.D. Singh, 1979. Changes in carbohydrates and lipids during embryonic development of Antheraea mylitta (Lepidoptera). J. Biosci., 1: 27-33.
Direct Link - Xin-Pei, Y., Z. Bo-Xiong, X. Meng-Kui, Y. Guo-Hua, C. Qi-Long and T. Xiao-Fen, 2005. Composition and changes of yolk proteins from silkworm Bombyx mori during embryonic development stages. Chinese J. Agric. Biotechnol., 2: 99-106.
CrossRef - Seidavi, A.R., A.R. Bizhannia, M. Ghanipoor and A.A.A. Qotbi, 2008. Investigation on improvement possibility of resistance, production and reproduction traits in 3p, 2p and p generations in three japanese pure lines of silkworm Bombyx mori L., using individual selection in 3P generation. Asian J. Anim. Vet. Adv., 3: 443-447.
CrossRefDirect Link - Sujatha, S. and B. Joseph, 2011. Development of tapioca based oligidic diet and its effects of mass rearing performance (biology) on lepidopteran pest of Spodoptera litura (Fab.) (Lepidoptera: Noctuidae). J. Entomol., 8: 174-182.
CrossRefDirect Link - Temerak, S.A., 2006. Role of moth scales remain on the top of egg masses of Spodoptera littoralis and its interference with the performance of the natural bio-product spinosad. J. Entomol., 3: 102-107.
Direct Link - Woods, H.A. and R.T. Bonnecaze, 2006. Insect eggs at a transition between diffusion and reaction limitation: Temperature, oxygen and water. J. Theoretical Biol., 243: 483-492.
CrossRef - Yamaoka, K. and T. Hirao, 1981. Mechanism of ovipositional behaviour of Bombyx mori, time gating and accumulation of the internal factor. Int. J. Invertbr. Reprod., 4: 169-180.
Direct Link