
Nidhi Wadhwa
Laboratory of Bee Behaviour and Pollination Ecology, Department of Zoology and Aquaculture, CCS Haryana Agricultural University, Hisar-125 004, India
Ram C. Sihag
Laboratory of Bee Behaviour and Pollination Ecology, Department of Zoology and Aquaculture, CCS Haryana Agricultural University, Hisar-125 004, India
Journal of Entomology
Year: 2012 | Volume: 9 | Issue: 4 | Page No.: 187-207







ABSTRACT
Sarpagandha (Rauvolfia serpentina (Linn.) Benth., ex Kurz.) is an important medicinal plant. Flowers of this plant are protogynous and need cross-pollination for seed set. The characterization of its pollinators and pollination mode is, therefore, important. Entire ranges of flower visitors of sarpagandha were captured with hand net from its field and were identified. These visitors were characterized as pollinators and non-pollinators based on their mode of working on the flowers. Their relative abundances, foraging rates activity durations, number of pollen grains carried on the body, effect of multiple visits were used to determine their pollinating efficiencies. At Hisar (Haryana, India), nineteen insect species were found to visit the flower of sarpagandha. The insects visiting the flowers of sarpagandha showed two types of foraging modes. On the basis of these modes, the flower visitors were characterized as pollinators and non-pollinators. Some foragers of A. dorsata and A. mellifera were base-foragers (nectar thieves/non-pollinators). However, all the lepidopterous species and some of the hymenopterous species were exclusively front-foragers (pollinators). Most abundant were two species of butterflies viz. Papilio demoleus and Pieris brassicae and three species of bees viz. Mellisodes sp., Xylocopa fenestrata and Megachile sp. The peak activity of all the insect visitors was observed at 1300 h. Maximum duration of activity in the field was shown by Papilio demoleus. All the visitors carried excessively higher number of pollen grains on their bodies than required by the flower for fruit set. Incremental visits also did not have significant increase on the fruit set. Therefore, on the basis of population abundance, foraging mode, foraging rate and activity duration, pollinating efficiency of the visitors was determined, where in Papilio demoleus was ranked as the best pollinator of sarpagandha at Hisar, that confirmed our earlier belief about predominance of psychophilous mode of pollination (i.e., pollination by butterflies) in sarpagandha.
PDF Abstract XML References Citation
Received: November 05, 2011;
Accepted: January 26, 2012;
Published: March 06, 2012
How to cite this article
Nidhi Wadhwa and Ram C. Sihag, 2012. Psychophilous Mode of Pollination Predominates in Sarpagandha (Rauvolfia serpentina). Journal of Entomology, 9: 187-207.
DOI: 10.3923/je.2012.187.207
URL: https://scialert.net/abstract/?doi=je.2012.187.207
DOI: 10.3923/je.2012.187.207
URL: https://scialert.net/abstract/?doi=je.2012.187.207
INTRODUCTION
Sarpagandha (Rauvolfia serpentina (Linn.) Benth. ex Kurz.) is an important medicinal plant known in Indian medicinal literature. It belongs to the family Apocynaceae which is known for having several plants containing useful alkaloids. Five species of sarpagandha have been recorded in India, of which R. serpentina has attained a great reputation as a medicinal plant (Sahu, 1983; Goel et al., 2007; Dey and De, 2010). A large number of alkaloids have been isolated from various species of sarpagandha; reserpine is pharmacologically the most potent. Other important alkaloids are: reserpinine, deserpidine, deserpideine, serpentine, serpentinine, ajmaline, ajmalinine and rauwolfinine. The total alkaloid content of the root varies from 1.7 to 3.0% of which the bark alone accounts for nearly 90% (Blackwell, 1990). Powdered taproots of sarpagandha have, for centuries, been used medicinally in India for the treatment of moon disease or lunacy and also for the treatment of snakebites and insect stings. The drug is used as a sedative and hypnotic agent and for reducing blood pressure (Ebadi, 2002). The alkaloids in the drug stimulate the central nervous system. It is also largely used in insanity, intestinal disorders and cardiac diseases. It is more suitable for cases of mild anxiety or for the patients of chronic mental illness. The drug has tranquillizing effect (Dutta and Varmani, 1964). The decoction of roots is given to pregnant women during labour pains to increase uterine contraction. Juice of leaves of sarpagandha is used for the cure of corneal opacity of the eye. The flower buds are used in bronchitis. Flowers are also used to stop nose bleeding (Farooq, 2005).
The flowers of angiosperms attract a wide variety of visitors; the ultimate attraction is the floral reward constituted by nectar and pollen. The sweet floral reward, the nectar, is the primary attractant and its presentation pattern in the flower determines how easily it is available to a seeker (Abrol and Sihag, 1997). A pollinator is an agent that brings about pollination by transferring pollen grains from the anthers of a flower to the stigma of the flower (s) of the same species. In practice, however, not all the insect visitors to the flower of a plant are pollinators, eventhough they are defined as such by some research workers (Free and Ferguson, 1983; Kevan and Baker, 1983). The floral visitor may harvest the floral reward (nectar) through a legitimate or illegitimate route. The foraging mode of the visitors to the flowers of a plant would, therefore, reveal whether they are its pollinators or non-pollinators. It is, therefore, of great importance to distinguish between pollinating and non-pollinating visitors. The population abundance, activity duration, foraging rate, pollen load and multiple visits are other parameters that influence the pollinating efficiency of the flower visitors (Sihag and Rathi, 1994; Priti and Sihag, 1997, 1998, 1999, 2000a, b; Gahlawat et al., 2002a, b; Narwania et al., 2003). In fact, the best pollinator is one that realizes maximal reproductive potential in the plant. The visitor’s behaviour, therefore, has great bearing on the reproductive success of a plant it visits. The latter is often pollinators limited and may be impaired if the flowers are not visited by a suitable pollinator, as well as if their number is not adequate. That is why, characterization of the appropriate pollinators of a plant species is important. With this aim the present study was undertaken on the pollinators of sarpagandha, a medicinal plant par excellence.
This paper examines and compares the values of different pollination attributes of flower visitors of sarpagandha and identifies its most effective pollinator(s) in semi-arid environment of North-west India. The knowledge of reproductive biology of a plant is a prerequisite for attempting any breeding programme (Singh et al., 2010). In our earlier study, we observed low reproductive potential in sarpagandha and we anticipated psychophilous mode of pollination in this plant (Sihag and Wadhwa, 2011). That view point has been confirmed in this paper.
MATERIALS AND METHODS
Sarpagandha plants were raised at the Research Farm of CCS Haryana Agricultural University, Hisar (India) as described by Sihag and Wadhwa (2011). To confirm the pollination mechanism in sarpagandha (R. serpentina), following studies were made:
Flower visitors of sarpagandha: The different insect species visiting the flowers of sarpagandha at Hisar (during its blooming period) were collected by hand net, got these identified in the Department of Entomology, CCS HAU, Hisar and a record was maintained.
Foraging behaviour of flower visitors of sarpagandha
Characterization of pollinators of sarpagandha: The flower visitors of sarpagandha were characterized as pollinators and non-pollinators on the basis of their foraging modes (method of working by a forager on a flower while harvesting pollen and/or nectar reward). The visitors collecting nectar and/or pollen through legitimate route were characterized as pollinators whereas, those through illegitimate route as non-pollinators (Inouye, 1980; Sihag, 1988; Sihag and Shivrana, 1997).
Foraging population abundance of flower visitors of sarpagandha: Three plots of 3x3 m size were randomly selected in the sarpagandha field. On a day, population abundances of different insect visitors on each plot were recorded by line transact method. The abundances were recorded at two hourly intervals, starting from commencement to the cessation of insect activity and were repeated at weekly intervals starting from commencement to the cessation of the flowering on the experimental plots following Sihag (1986). The recorded data were analyzed in “Completely Randomized Design” (Snedecor and Cochran, 1989) and the results were compared.
Foraging activity duration of the flower visitors of sarpagandha: The activity duration of pollinators was derived from the abundance data using following statistical formula suggested by Sihag and Rathi (1994):
Where:
| i | = | 1 to x, taking positive, whole numbers and finite values |
| T | = | Mean activity duration of pollinators |
| ni | = | Total number of insects of a species at ith hour of the day |
| ti | = | Total foraging activity duration of the visitors of a species active at ith hour of the day |
| N | = | Total number of visitors of a species through the course of the day |
Foraging rates: Foraging rate of a visitor was recorded in terms of flowers visited per minute. Time was recorded with the help of a stopwatch (chronometer) with accuracy of 0.1 sec. Observations were recorded at two hourly intervals on a day and were repeated at an interval of one week when the plant was in bloom. At any time, observations were recorded on ten insects of a species. The data were analyzed in “Completely Randomized Design” (Snedecor and Cochran, 1989) and foraging rates of different visitor species were compared.
Characterization of most efficient pollinator of sarpagandha
Number of loose pollen grains on the body of the pollinators: Loose pollen grains carried on the body of a forager of a species were counted as per method given by Traynor (1981). Insects foraging on the flowers of sarpagandha were captured from the field with the help of a forceps between 1000 and 1400 h. In case of bees, their hind legs were amputated and then put individually in glass vials containing 5 mL of 70% alcohol for counting the loose pollen grains sticking to their body. For this, an aliquot of 0.1 mL (replicated three times) was placed on haemocytometer slide and total numbers of pollen grains were counted under compound microscope. Ten samples were taken for each pollinator species and loose pollen grains present on each species were calculated and compared using “Completely Randomized Design” (Snedecor and Cochran, 1989).
Effect of single versus multiple visits of pollinators on seed set of sarpagandha: Newly opened flowers of sarpagandha were separately selected and guarded to receive one, two and three visits of a pollinator. The individual flowers were bagged till seed set and the numbers of seed set were recorded. One hundred flowers were taken for each ‘insect x visit’ and seed data were compared using “Completely Randomized Design” (Snedecor and Cochran, 1989).
Pollinating efficiency of the visitors of sarpagandha: When multiple visits have no incremental effect on seed set, then visitation frequency is the true measure of the pollinating efficiency of the visitor. Visitation frequency depends upon abundance, activity duration and foraging rate of the pollinator. For different pollination attributes (viz., population abundance of the flower visitors, mean activity duration and their foraging rates) the performance scores (PS) were derived for each species using following formula suggested by Sihag and Rathi (1994):
Where:
| i | = | 1 to x and j =1 to r, both taking positive, whole number and finite values |
| Psij | = | Performance-score of ith species for jth attribute |
| Nij | = | Importance value of ith species for jth attribute |
| Nj | = | Total importance values of all the species for jth attribute |
| s | = | Total number of species |
Performance-scores (PSs) tell about the relative performance of a species for a particular kind of foraging attributes e.g., abundance, activity duration, foraging rate etc. From various performance scores for different attributes of a species, Pollinating Index (PI) was derived by multiplying all the PSs of that species. The PIs so derived were then compared and on the basis of their values different species were ranked for their pollinating efficiency. The PIs so derived though are not the actual values of pollinating efficiency, yet these present an exact pattern of ranking of the pollinators of a plant species.
RESULTS
Flower visitors of sarpagandha: During its flowering period, ninteen insect species belonging to seven families under two orders were observed visiting the blossoms of sarpagandha at Hisar (Table 1). Among these visitors, 11 were hymenopterous and 8 were lepidopterous insect species.
| Table 1: | Different insects visiting the flowers of sarpagandha (R. serpentina) crop at Hisar |
 | |
 | |
| Plate 1: | Hymenopterous insect visitors of sarpagandha (R. serpentina) |
Among hymenopterous visitors, three species belonged to the family Apidae i.e., Apis dorsata, A. mellifera and A. florea (Plate 1; 1-3), five species belonged to family Anthophoridae i.e., Mellisodes sp., Pithitis smargdula, Xylocopa fenestrata, Xylocopa pubescens and Xylocopa sp. (Plate 1; 5,7, 8-10, 12 ( 8 and 9 are X. pubescens male and female, respectively), two species belonged to the family Megachilidae i.e., Megachile bicolor and Megachile sp. (Plate 1; 4, 6) and one species belonged to the family Vespidae i.e., Polistes hebraeus (Plate 1; 11).
 | |
| Plate 2: | Lepidopterous insect visitors of sarpagandha (R. serpentina) |
Among lepidopterous visitors, two species belonged to the family Papilionidae i.e., Papilio demoleus and Papilio polytes (Plate 2; 1, 3), two species belonged to the family Danaidae i.e., Danais chrysippus and Danais aglea creamer (Plate 2; 2, 4), three species belonged to the family Pieridae i.e., Pieris brassicae, cabbage butterfly and Eurema hecabe (Plate 2; 5-7) and one species belonged to the family Hesperiidae i.e., Pelopidas mathius (Plate 2; 8).
Characterization of pollinators of sarpagandha
Foraging modes: Two types of foragers were observed among the insects visiting the blossoms of sarpagandha. These were:
Side/base foragers: These foragers collected only nectar and that too from the side/base of the flower. Such foragers landed on the base of the flower, gripped it with their legs and inserted proboscis between petals using force to take nectar from the flower. In this mode, therefore, no part of the insect body could touch the reproductive parts of the flower. These foragers were categorized as nectar robbers. Since, these visitors did not come in contact with the reproductive organs of the flower; they did not play any role in the process of pollination of the flower. These visitors were designated as the n on-pollinators. Among all insect visitors of sarpagandha, Apis dorsata, A. mellifera, A. florea, Pithitis smargdula and Polistes hebraeus resorted to this foraging mode. Therefore, these visitors were not the pollinators of sarpagandha (Plate 3, 4).
Front/top foragers: These insects visited the flowers from its front/top. That is why, a part of the visitor’s body always came in contact with anthers and stigma of the flower while working on a flower, resulting in transfer of pollen and hence accomplishing the pollination activity. Such flower visitors were characterized as pollinators. Front foragers collected only pollen or both nectar and pollen in the same foraging attempt, voluntarily or involuntarily, butterflies were involuntary pollen gatherers. They visited the flowers of sarpagandha for nectar, but during their foraging attempt, they involuntarily transferred pollen from one flower to the other. All the lepidopterous insects visiting sarpagandha adopted this type of foraging mode.
 | |
| Plate 3: | A. dorsata visiting the flower of sarpagandha (R. serpentina) |
 | |
| Plate 4: | A. mellifera visiting the flower of sarpagandha (R. serpentina) |
These included: Papilio demoleus, Papilio polytes, Danais aglea creamer, Pieris brassicae, Danais chrysippus, Eurema hecabe, Pedis skipper and cabbage butterfly. Some hymenopterous insects visiting sarpagandha flowers, on the other hand, intentionally gathered both pollen and nectar in each foraging attempt. These were voluntary pollen gatherers and were designated as pollinators. Among the hymenopterous insects, Megachile bicolor, Megachile sp., Mellisodes sp., Xylocopa fenestrata, Xylocopa pubescens and Xylocopa sp. adopted this type of foraging mode and hence acted as pollinators of sarpagandha (Plate 5-9).
Foraging population abundance of flower visitors of sarpagandha: Among the hymenopterous insects, Mellisodes sp. was most abundant (24.71%), followed by Xylocopa fenestrata (18.52%) and Megachile sp. (7.64%). Among the lepidopterous insects, Papilio demoleus was most abundant (32.17%) followed by Pieris brassicae (9.66%). The proportion of remaining 14 species was very low (7.27%) (Table 2, 3). Average numbers of all the insect visitors, irrespective of their species, were low at the time of commencement of flowering.
 | |
| Plate 5: | Xylocopa fenestrata visiting the flower of sarpagandha (R. serpentina) |
 | |
| Plate 6: | Xylocopa pubescens visiting the flower of sarpagandha (R. serpentina) |
These numbers increased gradually and peaks were observed during peak flowering period, when the crop was in full bloom. The number of visitors then decreased gradually and drastically and these were very low at the time of cessation of flowering. In the beginning and at the end of the flowering, the numbers of insect visitors were significantly lower than those at the time of peak flowering. Differences in abundance among the insect species as well as the days were significant. At six observation hours (i.e., 0700, 0900, 1100, 1300, 1500, 1700 h) the number of different insect visitors i.e., Papilio demoleus, Mellisodes sp., Xylocopa fenestrata, Megachile sp., Pieris brassicae and others were 20.96, 16.08, 11.87, 4.98, 6.94 and 3.95 insects/m/min, respectively (Table 3). The peak numbers of insects were observed at 1100 and 1300 h which were significantly higher than those at all other observational hours.
 | |
| Plate 7: | Papilio demoleus visiting the flower of sarpagandha (R. serpentina) |
 | |
| Plate 8: | Papilio polytes visiting the flower of sarpagandha (R. serpentina) |
 | |
| Plate 9: | Mellisodes sp. visiting the flower of sarpagandha (R. serpentina) |
| Table 2: | Foraging rates of different insect pollinators of sarpagandha (R. serpentina) at different observation hours of the day |
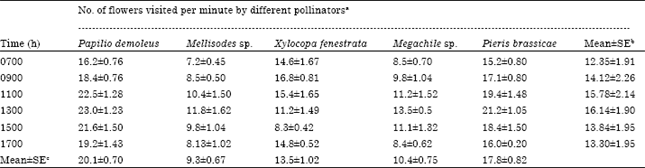 | |
| Mean±SE of a; 30 observation, b = 5 observations, c = 6 observations | |
| Table 3: | Relative abundance of insects pollinators of sarpagandha (R. serpentina) during blooming period |
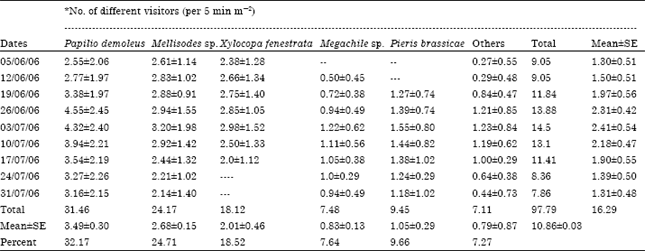 | |
| *Mean±SE of 54 observations | |
| Table 4: | Mean activity duration of pollinators of sarpagandha |
 | |
However, Xylocopa fenestrata, showed bimodal activity exhibiting two peaks, one during the morning hours and other during the evening hours; in between the abundance was low.
Foraging activity duration: All the insects started their activity in the morning and remained active till evening hours. Among all the insect visitors, Papilio demoleus remained active for the longest duration (7.6 h) on the blossoms of sarpagandha flower followed by Mellisodes sp. (6.8 h), Xylocopa fenestrata (3.3 h), Pieris brassicae (3.1 h) and Megachile sp. (2.5 h), respectively (Table 4).
Foraging rates: Higher foraging rate of a pollinator means more pollen transfer in many flowers, in a short duration of time that helps hastening the pollination process.
| Table 5: | Foraging rates of different insect pollinators of sarpagandha at different observation hours of the day |
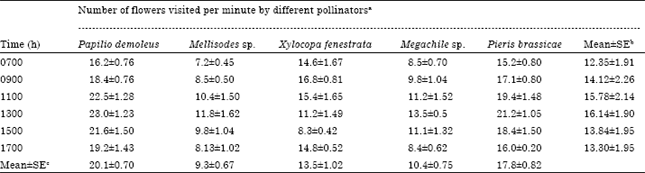 | |
| Mean±SE of a: 30 observation , b = 5 observations, c = 6 observations | |
| Table 6: | Number of loose pollen grains carried by the insect pollinators of sarpagandha |
 | |
| a: Mean±SD of 10 observations | |
The foraging rates of butterflies were significantly higher than those of the bees. Papilio demoleus had maximal foraging rate (20.1 flowers min-1) whereas Pieris brassicae was second (17.8 flowers min-1). Among the bees, Xylocopa visited maximal number of flower per minute (13.5 flowers min-1), followed by Megachile sp. (10.4 flowers min-1) and Mellisodes sp. (9.3 flowers min-1) (Table 5). On this basis of this attribute, butterflies seemed to contribute more than bees towards the pollination process of sarpagandha.
Characterization of most efficient pollinator of sarpagandha
Number of loose pollen grain on the body of pollinators: Number of loose pollen grains carried by different insects varied among species (Table 6). Mellisodes species carried maximal number of pollen grains (5087.66), followed by Megachile sp. (4780.33), Xylocopa fenestrata (4471), Papilio demoleus (2730) and Pieris brassicae (2513) and differences were significant (p = 0.05, ANOVA, Table 6).
Effect of single versus multiple visits of pollinators on seed set of sarpagandha: The differences among seed set and quality (test weight and germination) due to multiple visits were non-significant (p = 0.05, ANOVA, Table 7). This indicated that even a single visit of a pollinator was sufficient to pollinate the flower of sarpagandha. As explained earlier, this is due to the presence of very small number of ovules (only 4) in the ovary of sarpagandha and all the pollinators carried very large number of loose pollen grains in comparison to those required by the flower. Therefore, multiple visits data did not exactly confirm, whether sarpagandha is a butterfly or bee pollinated plant.
Visitation frequency as a measure of pollinating efficiency of the visitors and derivation of performance scores and efficiencies of pollinators of sarpagandha: Loose pollen grains carried on the body of the pollinators of sarpagandha can’t be taken as one of the parameters for determining pollinating efficiency.
| Table 7: | Effect of Multiple visits of different pollinators on seed set of sarpagandha |
 | |
| a: On the basis of 25 flowers with 100 number of expected seed set | |
| Table 8: | Pollinating efficiency of different pollinators of sarpagandha |
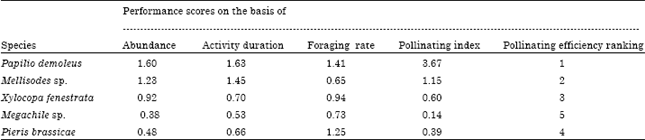 | |
This is because ovary of sarpagandha flower carried 4 ovules. All the pollinators were capable of transferring this number, as each carried much higher number of pollen grains on their body. Therefore, on the basis of abundance, foraging rate and mean activity duration insect pollinators were tentatively ranked for pollinating efficiency which could determine the most efficient pollinators among the several visitors of sarpagandha blossoms (Table 8). Among the 5 insect pollinators Papilio demoleus came out to be on the first place and seemed to be the best pollinator of this crop. Other pollinators with relatively lower ranking in pollinating efficiency were the long tongued bees- Mellisodes sp., Xylocopa fenestrata and Megachile sp. in descending order. Therefore, sarpagandha flowers seemed to rely for their pollination mainly on butterflies.
DISCUSSION
Seed/fruit production in plants depends upon several factors and inputs (Ahmad et al., 2003; Akhtar and Khaliq, 2003; Mari et al., 2005; Ansari, 2007; Goel et al., 2007; Macit et al., 2007; Iqbal et al., 2007; Solangi and Lohar, 2007; Talebi et al., 2007; Kumar et al., 2011); the managed pollination is one of these factors/inputs (Free, 1993; Sihag, 2001). For managed pollination of a plant, there is essential need to know about its pollinators. This article presents information on this aspect of sarpagandha.
Flower visitors of sarpagandha: There is no earlier report on the flower visitors of sarpagandha in semi-arid sub-tropics of north-west India, though many reports are available on other plants (Arya et al., 1994; Sihag and Rathi, 1994; Priti and Sihag, 1997, 1998, 1999, 2000a, b; Gahlawat et al., 2002a, b; Chaudhary and Sihag, 2003; Narwania et al., 2003; Bhattacharya et al., 2005). The entire ranges of flower visitors do tell about the diversity of species and groups (Table 1, Plate 1,2). However, these do not tell about as to how many of these are pollinators or non-pollinators. To ascertain this, their characterization is important.
Foraging behaviour of flower visitors of sarpagandha
Characterization of pollinators of sarpagandha: Nectar robbery can be facilitating or inhibitory in the reproductive fitness of a plant. Sampson et al. (2004) observed nectar robbery by bees in rabbit eye blueberry (Vaccinium ashei). Xylocopa virginica L. and Apis mellifera were the flower visitors of this plant. A. mellifera probed for nectar from robbery slit in the flowers previously made by male carpenter bees (Xylocopa virginica). This relationship between primary nectar robbers (carpenter bees) and secondary nectar thieves (honey bees) is poorly understood but only seemingly is unfavorable for pollination of this plant. Observations demonstrated that male carpenter bees were benign or even potentially beneficial floral visitors of blueberry (V. ashei). Their robbery of blueberry flowers in the southeast U.S.A. may attract more honeybee pollinators to the crop, thus enhancing its pollination potential. Here, nectar robbery is facilitating the reproductive fitness of rabbit eye blueberry. Nectar robbery in many other plants has been stated to be ecologically disadvantageous to the plant species, as this has a negative impact on the reproductive fitness of the plant (Koeman-Kwak, 1973; Barrows, 1976; Rust, 1979; Wyatt, 1980; McDade and Kinsman, 1980; Inouye, 1981; Roubik, 1982; Young, 1983; Roubik et al., 1985; Zimmerman and Cook, 1985; Addicott, 1986; Tyre and Addicott, 1993; Addicott and Tyre, 1995). The nectar robbers in sarpagandha, however, were of no use to this plant as these did not facilitate the visits of other visitors to the flowers of this plant to effect pollination.
In the present study, the insects visiting the blossoms of sarpagandha showed both types of foraging modes i.e., side foraging and front foraging (Plates 3-9). In side foraging mode, the foragers collected only nectar and that too from the sides of the flower. Such foragers landed on the base of the flower, gripped it with their legs and inserted proboscis between petals using force to take nectar from the flower. In this mode, therefore, no part of the insect body could touch the reproductive parts of the flower. These foragers were categorized as nectar robbers. Since, these visitors did not come in contact with the reproductive organs of the flower; they did not play any role in the process of pollination of the flower. These visitors were designated as the non-pollinators. Among all insect visitors of sarpagandha, Apis dorsata, A. mellifera, A. florea, Pithitis smargdula and Polistes hebraeus resorted to this foraging mode. In the front mode of foraging, a part of the flower visitor’s body always came in contact with anthers and stigma while working on a flower, resulting in transfer of pollen and hence accomplishing the pollination activity. Such flower visitors were characterized as pollinator’s (Sihag, 1988, Sihag and Shivrana, 1997). Front foragers collected both nectar and pollen or only pollen in the same foraging attempt, voluntarily or involuntarily. Bees were voluntary pollen gatherers. All the lepidopterous: Papilio demoleus, Papilio polytes, Danais aglea creamer, Pieris brassicae, Danais chrysippus, Eurema hecabe, Pedis skipper, cabbage butterfly and most of the hymenopterous insects viz. Megachile bicolor, Megachile sp., Mellisodes sp., Xylocopa fenestrata, Xylocopa pubescens and Xylocopa sp. visiting sarpagandha adopted this type of foraging mode. These pollinators, while working on sarpagandha flowers as top foragers alighted on the petals, approached through the mouth of the flower and inserted their proboscis for nectar only, or they alighted on the petals probed the flower from its front and while doing so, touched the reproductive parts of the flower and transferred pollen. Therefore, the insects which are front foragers always acted as pollinators. These visitors are beneficial to the protogynous flowers of sarpagandha in bringing out cross-pollination.
Foraging population abundance of flower visitors of sarpagandha: Abundance of the pollinators presents the full spectrum of different species that visit the flowers of a crop during the full span of its flowering period (Table 2, 3). Higher the visitation frequencies more are the chances of a flower to be visited repeatedly by a species. That should ensure more pollination in the flowers and more will be the contribution towards the reproductive success of the plant. The latter is measured in terms of degree of seed set and seed/ fruit yield (Free, 1993).
Pollination process is more efficient when there are more flowers on a plant. At peak flowering, the availability of flowers is more than commencement and cessation and large number of insect visit the crop during this period to help maximize the pollination process. Therefore, flower number clearly influences the pollinator abundance and in turn level of pollination; plants with many flowers often attract more floral visitors than those with fever (Free, 1993). In the present investigation, a fluctuation in visits of insect pollinators on different days was observed. The visits were low at the time of commencement and cessation of the flowering but these remained high during mid flowering period (Table 2). This difference should be due to variation in the floral density during the span of blooming on sarpagandha crop (Sihag and Wadhwa, 2011). In this study, the abundance of foragers followed the same pattern as did the floral density. At the time of commencement of flowering period, the abundance of all the foragers was less because there was less number of flowers (and lower availability of nectar amount) to attract the insect visitors. Abundance increased gradually as the flowering period progressed, the peak of abundance was recorded when the crop was in full bloom. Abundance of all the foragers again decreased at the cessation of the flowering period because at that time again there was less number of flowers (Sihag and Wadhwa, 2011). These findings are in agreement with those of Pyke et al. (1977) who reported that during the time of peak flowering, more flowers were on the plant and they provided more nectar and pollen simultaneously on the same plant to attract more pollinators.
The entire organisms in the community are not equally important in determining the nature and function of the whole community. The latter are classified on the basis of the dominating organisms. The latter strongly affect the environment of all other species and these are called ecological dominants (Odum, 1971). In an ecosystem, pollination is accomplished by a large number of pollinator species. However, only a few of them largely control the pollination process and are said to be the dominant pollinators. Removal of the dominant pollinator species would adversely affect the pollination process in the reference plant. Though, the removal of a non-dominant species would produce much less change on the pollination process, yet it will affect the species diversity. However, populations of the entire organisms in the ecosystem are extremely important to regulate and stabilize the ecosystem (Odum, 1971). This is very much true in case of pollinator-plant systems too, where large numbers of pollinator species are present. This is because, the diverse populations of insect pollinators are extremely important to regulate and stabilize the pollination process of entomophilous plants.
The results of this study revealed that pollination process in sarpagandha might be destabilized; if a dominant species of pollinator (e.g., Papilio demoleus) is removed, though other non-dominant pollinator species might carry out some pollination. So, dominant insect pollinator plays an important role in the pollination process in the agro-ecosystem. In this study many insect species were found to visit the sarpagandha crop at sub-tropical Hisar (Table 1). The numbers of insect species visiting the blossoms of sarpagandha were Nineteen. Eleven species belonged to the order Hymenoptera and eight species belonged to the order Lepidoptera. However, all these species were not equally efficient and suitable pollinators of this crop; only lepidopterous butterflies and some of hymenopterous bees were important pollinators. Throughout the flowering period of sarpagandha, Papilio demoleus was most abundant followed by Mellisodes sp., Xylocopa fenestrata, Pieris brassicae and Megachile sp. (Table 1). The difference in visitor’s spectrum was perhaps due to their suitability to the flowers.
Pollinators’ abundance has earlier been taken as a measure of their pollinating efficiency (Schemske and Horvitz, 1984). Though individually, P. demoleus was most abundant, yet as a group, bees out numbered the butterflies. Among the pollinators, about 42% of the total visitors were butterflies whereas, about 51% were bees. Therefore, structural evidence notwithstanding (Sihag and Wadhwa, 2011), the abundance data did not support that sarpagandha depends predominantly on butterflies for pollination of its flowers.
Foraging activity duration: Duration of foraging activity mainly depends upon tolerance and threshold values of temperature. Insects are cold blooded animals and they start their activities, when the temperature crosses its lower threshold values. As the temperature increases, insect activity also increases due to increase in metabolism in their bodies (Table 3). In fact, the best adapted pollinators should remain active for a longer time on the blossom of a crop. Longer duration of the mean foraging activity of the visitors would be useful in enhancing the pollination process in a plant species. Insect pollinators will pollinate more flowers if they remain active for a longer duration. Foraging activity duration of insect pollinators vary from species to species and plant to plant (Free, 1993). In the present study, on the blossom of sarpagandha, Papilio demoleus remained active for longest duration followed by Mellisodes sp., Xylocopa fenestrate, Pieris brassicae and Megachile sp., respectively (Table 4). Again, sarpagandha showed main dependence on psychophilous pollination.
Foraging rates: Foraging rate is the number of flowers visited per minute by an insect. Higher the foraging rate of an insect pollinator means more the pollen transfer and in many flowers, in a short duration of time that helps hastening the pollination process. It is influenced by complex system of factors like, insect body size, flower shape and size, air temperature and relative humidity etc. In this study, air temperature was the most important. This is because, the individual insects are poikilothermal and ambient temperature influences their body temperature and body metabolism to a greater degree. That is why, rising temperature significantly increases the speed of flower visitors (Benedek and Prenner, 1972).
The foraging rates of different insect species were low in the morning and evening (i.e., 0700-800 and 1600-1700 h) and high during mid day (1100-1500 h). All insect species showed maximum activity during 1100-1500 h. They visited more number of flowers during mid day hours because of rise in temperature (Table 5). Arya (1993) also observed that the foraging rates of the bees were more during mid-day hours. This indicates that maximum pollination in the crops was effected during/around mid day.
In the present study, the foraging rates of lepidopterous insects were significantly higher than those of the hymenopterous insects, Papilio demoleus had highest foraging rate followed by Pieris brassicae, Xylocopa fenestrata, Megachile sp. and Mellisodes sp. (Table 5). On this basis too, psychophilous mode of pollination seemed to be more prevalent in sarpagandha.
Characterization of most efficient pollinator of sarpagandha: The efficiency of an insect to pollinate the flowers of a plant is its pollinating efficiency. A large number of insects visit the flowers of a plant. However, all are not equally useful to the crop as its pollinators. It is not possible to determine the best pollinator of a plant without studying the relative pollinating efficiency of its flower visitors. There are a number of ways to find whether a flower visitor is a pollinator or not (Sihag, 1988). The pollinating efficiency is determined by studying the various pollinating attributes like visitation frequency of the pollinators, their foraging modes and foraging rates, activity duration and number of pollen grains carried on the body. All these attributes add to the determination of relative pollination efficiencies of the pollinators and help characterization and differentiation between best and supplementary pollinators of a crop (Sihag and Rathi, 1994).
Number of loose pollen grain on the body of pollinators: Different visitor species carry different number of pollen grain depending upon their body size, number of hairs on the body and pollen display pattern of the flower. For example, in glory bower (Clerodendron trichotomum Thunb.), the number of pollen grains carried by two butterflies visiting the flowers were 1,776 for P. helenus and 2,817 for P. protenor (Suzuki et al., 1987).
Suzuki et al. (1987) observed that the butterflies (Papilio helenus and P. protenor) were found to feed mainly on the nectar of the glory bower (Clerodendron trichotomum) which was the most abundant nectar plant in summer in the study area. Both the species were found to have a proboscis longer than 24 mm corresponding to the length of the corolla tube of C. trichotomum. Since, the flower of C. trichotomum has long protruding sexual organs, its pollen grains were found to adhere efficiently to the bodies of butterflies, mainly the thorax, during nectar feeding. Most of the butterflies became loaded with C. trichotomum pollen. The flowers opened at any time of day but more frequently in the morning. The nectar was secreted throughout the day. In the maturation of the protandrous flower of C. trichotomum, the duration of the pistillate phase was about twice as long as the staminate phase. The long flowering period and the short duration of the staminate phase resulted in asynchrony of the flowering stages even within a single cyme on a tree. Such asynchrony and the abundance of attractive flowers on a tree facilitate efficient pollination by the butterflies.
More pollen is expected to be transferred by an insect if it carries larger number of loose pollen grains and may pollinate several flowers one after the other (Free, 1993). However, this is not always true. Loose pollen grains carried on the body of the pollinators of sarpagandha can’t be taken as one of the parameters for determining pollinating efficiency. This is because; ovary of sarpagandha flower carried only four ovules (Sihag and Wadhwa, 2011). All the pollinators were capable of transferring this number, as each carried much higher number of pollen grains on their body than the actually required by a flower (Table 6).
Effect of single versus multiple visits of pollinators on seed set of sarpagandha: The differences among seed set due to multiple visits were non-significant (p = 0.05, ANOVA, Table 7). This indicated that even a single visit of a pollinator was sufficient to pollinate the flower of sarpagandha. As explained earlier, this is due to the presence of very small number of ovules (only 4) in the ovary of sarpagandha and all the pollinators carried very large number of loose pollen grains in comparison to those required by the flower. Therefore, multiple visits data did not exactly confirm whether sarpagandha is a butterfly or bee pollinated plant.
Role of multiple visits on seed set has been reported in plants where ovaries in their flowers have large number of ovules. The important examples are watermelon (Adlerz, 1966) and kiwifruit (Donovan and Read, 1991). In such cases, the individual flower is benefited from the larger number of pollen grains carried by a visitor. Therefore, visitors transferring more pollen grains in a single visit or making multiple visits should be more useful to the plant. This situation does not exist in sarpagandha. That is why; the differences in seed set data are non-significant.
Pollinating efficiency of the visitors of sarpagandha: Higher the visitation frequency more is the chances of a flower to be visited repeatedly by a pollinator species. That should ensure more pollination in the flowers and more will be the contribution towards the reproductive success of the plant. The latter is measured in terms of degree of seed set and seed/fruit yield (Free, 1993). This contention is true in case of sarpagandha, where single visit of the pollinator is sufficient to set all the four seeds in the flower. Visitation frequency depends upon abundance, activity duration and foraging rates of the visitors.
On the basis of these attributes, pollinators of sarpagandha were tentatively ranked for their efficiencies (Table 8). Among the five pollinators, Papilio demoleus came out to be on the first place and seemed to be the best pollinator of this crop. Other pollinators with relatively lower ranking in pollinating efficiency were the long tongued bees Mellisodes sp., Xylocopa fenestrata and Megachile sp. in descending order. Therefore, sarpagandha flowers seemed to rely for their pollination mainly on butterflies.
The efficiency of an insect to pollinate the flowers of a plant is its pollinating efficiency. A large number of anthophiles visit the flowers of a plant. However, all are not equally useful to the plant as its pollinators. It is not possible to determine the best pollinator of a plant without studying their relative pollinating efficiency. There are a number of ways to find whether a flower visitor is a pollinator or not (Sihag, 1988). The pollinating efficiency is determined by studying the various pollinating attributes like visitation frequency of the pollinators, their foraging modes and foraging rates, activity duration and number of pollen grains carried on the body. All these attributes add to the determination of relative efficiencies of the pollinators and help characterization and differentiation between best and supplementary pollinators of a plant (Sihag and Rathi, 1994). Among the nineteen insect species that visited the flowers of sarpagandha, only five were major pollinators of this plant (Table 8). Among these, Papilio demoleus was the most efficient pollinator. Others in this ranking were Mellisodes sp., Xylocopa fenestrata and Megachile sp. in descending order (Table 8). The butterfly, Papilio demoleus could bring out 61.69% pollination in this plant that was maximal among all the visitors butterflies together contributed to about 68.25% pollination whereas bees alone could do only 31.75% (Table 8). This confirmed the major dependence of sarpagandha on butterflies for pollination. This overall revealed the predominance of psychophilous mode of pollination in sarpagandha in semi-arid environment of North-west India.
CONCLUSION
The blossoms of sarpagandha attracted 19 insect species. Out of these, honeybees and wasps were the nectar robbers and non-pollinators. Three long tongued bees were, however, pollinators but were at a lower rank in pollinating efficiency. The butterflies were the most efficient pollinators of this plant. This confirms our earlier belief of predominance of psychophilous mode of pollination in sarpagandha. Therefore, for the conservation of sarpagandha, conservation of its butterfly pollinators is most important.
ACKNOWLEDGMENTS
We are thankful to Dr. C.S. Tyagi, Sectional Head, Medicinal Plants, for raising the sarpagandha plants and providing the field facilities. The financial assistance received by NW as Merit Stipend from CCS HAU is gratefully acknowledged.
REFERENCES
- Addicott, J.F., 1986. Variation in the costs and benefits of mutualism: The interaction between yuccas and yucca moths. Oecologia, 70: 486-494.
Direct Link - Addicott, J.F. and A.J. Tyre, 1995. Cheating in an obligate mutualism: How often do yucca moths benefit yuccas? Oikos, 72: 382-394.
Direct Link - Adlerz, W.C., 1966. Honeybee visit numbers and watermelon pollination. J. Econ. Entomol., 59: 28-30.
Direct Link - Ahmad, S., T. Muhammad, A. Karim, A. Jabbar and G. Ahmad, 2003. Effect of pollinators and insecticides on seed reduction of onion (Allium cepa L.). Asian J. Plant Sci., 2: 653-654.
CrossRefDirect Link - Ansari, N.A., 2007. Effect of density, cultivars and sowing date on onion sets production. Asian J. Plant Sci., 6: 1147-1150.
CrossRefDirect Link - Arya, D.R., R.C. Sihag and P.R. Yadav, 1994. Diversity, abundance and foraging activity of insect pollinators of sunflower (Helianthus annuus L.) at Hisar. Indian Bee J., 56: 172-178.
Direct Link - Barrows, E.M., 1976. Nectar robbing and pollination of Lantana camara (Verbanaceae). Biotropica, 8: 132-135.
Direct Link - Benedek, P. and J. Prenner, 1972. Effect of temperature on the behaviour and pollinating efficiency of honey bees on winter rape flowers. Zeitschrift Angewandte Entomol., 71: 120-124.
CrossRefDirect Link - Chaudhary, N. and R.C. Sihag, 2003. Diversity, foraging behaviour and foraging efficiency of different pollinators visiting onion (Allium cepa L.) blossoms. Korean J. Apicult., 18: 103-108.
Direct Link - Free, J.B., 1993. Insect Pollination of Crops. 2nd Edn., Academic Press, London, UK., ISBN: 9780122666513, Pages: 684.
Direct Link - Gahlawat, S.K., S.K. Narwania, R.C. Sihag and Ombir, 2002. Studies on the diversity, abundance, activity duration and foraging behaviour of insect pollinators of cucumber (Cucumis sativus L.) at Hisar. Korean J. Apicult., 17: 69-76.
Direct Link - Iqbal, M., N. Islam, K. Hayat and T. Muhammad, 2007. Management of cotton crop under high Cotton leaf curl virus attack. Asian J. Plant Sci., 6: 1125-1130.
CrossRefDirect Link - Kevan, P.G. and H.G. Baker, 1983. Insects as flower visitors and pollinators. Annu. Rev. Entomol., 28: 407-453.
CrossRefDirect Link - Kumar, P., A.K. Misra and D.R. Modi, 2011. Current status of mango malformation in India. Asian J. Plant Sci., 10: 1-23.
CrossRefDirect Link - Macit, I., A. Koc, S. Guler and I. Deligoz, 2007. Yield, quality and nutritional status of organically and conventionally-grown strawberry cultivars. Asian J. Plant Sci., 6: 1131-1136.
CrossRefDirect Link - Mari, J.M., N.H. Rizvi, S.M. Nizamani, K.H. Qureshi and M.K. LOhar, 2005. Predatory efficiency of Manochilus sexmaculatus Fa. And Coccinella undecimpunctata Lin. (Coccinellidae: Coleoptera) on alfalfa aphid, Therioaphis trifolii (Monell.). Asian J. Pl. Sci., 4: 365-369.
Direct Link - McDade, L.A. and S. Kinsman, 1980. The impact of floral parasitism in two neotropical hummingbird-pollinated plant species. Evolution, 34: 944-958.
Direct Link - Narwania, S.K., S.K. Gahlawat, Ombir and R.C. Sihag, 2003. Diversity, Abundance, foraging rates and pollinating efficiency of insects visiting wanga (Cucumis melo ssp. Melo) blossoms at Hisar. Korean J. Apicult., 18: 29-36.
Direct Link - Priti and R.C. Sihag, 2000. Diversity, Visitation Frequency, Foraging Behaviour and Pollinating Efficiency of Insect Pollinators Visiting Turnip Blossoms. In: Asian Bees and Beekeeping: Progress of Research and Development, Matsuka, M., L.R. Verma, S. Wongsiri, K.K. Shrestha and U. Partap (Eds.). Oxford and IBH Publ. Co. Pvt Ltd., New Delhi, India, ISBN-13: 9781578080847, pp: 190-192.
- Pyke, G.H., H.R. Pulliam and E.L. Charnov, 1977. Optimal foraging: A selective review of theory and tests. Q. Rev. Biol., 52: 137-154.
Direct Link - Roubik, D.W., 1982. The ecological impact of nectar-robbing bees and pollinating hummingbirds on a tropical shrub. Ecology, 63: 354-360.
Direct Link - Roubik, D.W., N.M. Holbrook and G.V. Parra, 1985. Roles of nectar robbers in reproduction of the tropical treelet Quassia amara (Simaroubaceae). Oecologia, 66: 161-167.
Direct Link - Rust, R.W., 1979. Pollination of Impatiens capensis: Pollinators and nectar robbers. J. Kansas Entomol. Soc., 52: 297-308.
Direct Link - Sampson, B.J, R.G. Danka and S.J. Stringer, 2004. Nectar robbery by bees Xylocopa viginica and Apis mellifera contribution to the pollinator of Rabbiteye blueberry. J. Econ. Entomol., 97: 735-740.
PubMedDirect Link - Schemske, D.W. and C.C. Horvitz, 1984. Variation among floral visitors in pollination ability: A precondition for mutualism specialization. Science, 225: 519-521.
CrossRefDirect Link - Sihag, R.C., 1986. Insect pollination increases seed production in cruciferous and umbelliferous crops. J. Apicult. Res., 25: 121-126.
CrossRefDirect Link - Sihag, R.C., 1988. Characterization of the pollinators of cultivated cruciferaus and leguminars crops of sub-tropical Hisar, India. Bee Wld., 69: 153-158.
Direct Link - Singh, K.P., Bhavana and G. Dhakre, 2010. Reproductive biology of Evolvulus alsinoides L. (Medicinal Herb). Int. J. Bot., 6: 304-309.
CrossRefDirect Link - Solangi, B.K. and M.K. Lohar, 2007. Effect of some insecticides on the population of insect pests and predators on okra. Asian J. Plant Sci., 6: 920-926.
CrossRefDirect Link - Suzuki, N.Y., A. Niizume and K. Kiritani, 1987. Studies on ecology and behaviour of Japanese black swallow tail butterflies. 6. Nectar feeding of Papilio helenus nicconicolens Butler and P. protenor demetrius Cramer as main pollinators of glory bower (Clerodendron trichotomum). J. Ecol. Res., 2: 41-52.
Direct Link - Talebi, R., F. Fayaz and N.A.B. Jelodar, 2007. Correlation and path coefficient analysis of yield and yield components of chickpea (Cicer arietinum L.) under dry land condition in the West of Iran. Asian J. Plant Sci., 6: 1151-1154.
CrossRefDirect Link - Traynor, 1981. Use of fast and accurate method for evaluating pollen production of alfalfa and almond flowers. Amer. Bee J., 121: 23-25.
Direct Link - Tyre, A.J. and J.F. Addicott, 1993. Facultative non-mutualistic behaviour by an obligate mutualist: Cheating, by yucca moths. Oecologia, 94: 173-175.
Direct Link