
A. H. Fouly
College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah-51452, Saudi Arabia
M. A. Al-Deghairi
College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah-51452, Saudi Arabia
N.F. Abdel Baky
College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah-51452, Saudi Arabia
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 1 | Page No.: 52-62







ABSTRACT
Development, feeding capacity and life table parameters of the predatory mite Typhlodromips swirskii (Athias-Henriot) fed three different levels of whitefly eggs Bemisia tabaci Genn. were studied under laboratory conditions of 26°C and 70% RH. Both organisms were collected from eggplant leaves growing in an greenhouse in Al-Qassim region, center of Saudi Arabia in 2009. In leaf disc assays, increasing number of prey eggs accelerated the development of T. swirskii. Duration of developmental stages and feeding capacity of T. swirskii were significantly affected by the number of whitefly eggs offered. During an ovipositional period of 13.1, 18.9 and 19.5 days, adult female of T. swirskii consumed an average of 41.6, 128.4 and 140.1 prey eggs when each female was provided with 4, 8 and 12 prey eggs day-1 and laid an average of 10.5; 12.8 and 15.7 eggs day-1, respectively. Offering the predatory mite with 4, 8 and 12 prey eggs day-1 caused a mean generation time (T) of T. swirskii was 17.15; 17.66 and 15.45 days while, the net reproductive rate (Ro) averaged 10.86, 20.98 and 22.12 female progeny/female and the intrinsic rate of increase (rm) of T. swirskii was 0.22, 0.17 and 0.14 female progeny/female/day, respectively. The finite rate of increase erm (λ) was at its highest level (1.23) when T. swirskii individuals was provided with 12 eggs day-1 and declined to 1.19 and 1.15 after feeding on 8 and 4 prey eggs day-1, respectively. Crowding of T. swirskii significantly decreased feeding capacity and egg deposition.
PDF Abstract XML References Citation
Received: April 07, 2010;
Accepted: May 19, 2010;
Published: July 17, 2010
How to cite this article
A. H. Fouly, M. A. Al-Deghairi and N.F. Abdel Baky, 2011. Biological Aspects and Life Tables of Typhlodromips swirskii (Acari: Phytoseiidae) Fed Bemisia tabaci (Hemiptera: Aleyroididae). Journal of Entomology, 8: 52-62.
DOI: 10.3923/je.2011.52.62
URL: https://scialert.net/abstract/?doi=je.2011.52.62
DOI: 10.3923/je.2011.52.62
URL: https://scialert.net/abstract/?doi=je.2011.52.62
INTRODUCTION
The whitefly, Bemisia tabaci Gennadius (Homoptera: Aleyrodidae), is one of the most intractable and worldwide damaging pest within the agricultural and horticultural production systems (Perring, 2001; Carabali et al., 2005). Since, late 1980's, the insect has risen from relative obscurity to become one of the primary insect pests of agricultural crops (Lin et al., 2007). Many biotypes of this pest have been identified around the world (Dong et al., 2007; Touhidul Islam and Shunxiang, 2007), which suggest that B. tabaci is a species complex (Brown et al., 1995; Oliveira et al., 2001). Therefore, it is considered one of the worst world’s top 100 invasive agricultural pest species. During the last decades, chemical control was the most efficient method to minimize whitefly damages to crop production, although such practice is hazardous to water, soil, environment and human health. Therefore, new control tactics such as biological control using parasites and predators became necessary (Al-Rehiayani and Fouly, 2005, 2006; Abdel-Baky and Al-Deghairi, 2008; Al-Deghairi, 2009; Fouly and Al-Rehiayani, 2010). Among predaceous organisms that can suppress insect populations, phytoseiid mites which considered effective biological agents against wide range of harmful insect and mite pests that are commercially used in biological control and IPM programs (McMurtry and Croft, 1997; Fouly et al., 1995; Fouly, 1997; Nomikou et al., 2002, 2004; Inbar and Gerling, 2008; Fouly and Al-Rehiayani, 2010). Some of phytoseiid mites feed mainly on tetranychid mites(specialists) but others can feed on mite preys and as well as insects species such as whiteflies, thrips and plant pollens (generalists) (Fouly and Hassan, 1992; Fouly, 1997; McMurtry and Croft, 1997; Messelink et al., 2006; Al-Shammery, 2010).
Field observations showed that Typhlodromips swirskii (Athia-Henriot) is frequently found in association with phytophagous tetranychid mites, scale insects and whiteflies (Nomikou et al., 2001, 2002, 2004; Inbar and Dan Gerling, 2008; Messelink et al., 2006; Momen et al., 2009). This predatory mite is considered one of the generalist phytoseiid predators (McMurtry and Croft, 1997). Fouly and Al-Rehiayani (2010) surveyed predatory mites in Qassim region, central of Saudi Arabia and observed T. swirsiki feeding on different immature stages of whitefly infesting eggplants growing in greenhouses. So far, there were no considerable attempts to use phytoseiid predators for control of whiteflies, despite the fact that effective biological control for this insect pest is much needed (Nomikou et al., 2002, 2004).
The present protocol aims to throw some light on different biological aspects of T. swirskii provided with three levels of eggs of B. tabaci under laboratory conditions. Influence of the predator crowding on its egg production and feeding capacity was also studied.
MATERIALS AND METHODS
Whitefly culture: Sweet potato whitefly adults (B. tabaci) were collected by aspirator from insecticide free squash plants at Al-Qassim region, central of Saudi Arabia and maintained in large screen cages planted with white kidney beans (Phaseolus vulgaris L.) in pots during the growing season in 2009. Whitefly colony was kept under laboratory conditions at the room temperature 26±1°C, 65±3% RH with a 14:10 (L:D) photoperiod. At least three successive generations of B. tabaci was gained under these conditions, to serve for other biological experiments. To produce plants with heavy infestation of whitefly, newly white kidney pots were infested with whitefly adults and left to deposit eggs for three days, resulting in at least 50-100 eggs leaf-1. After ten days, the colony had a mixed whitefly eggs and immature stages to serve as a food source for predaceous mites.
Mite culture: Individuals of T. swirskii were collected from eggplant leaves grown at the experimental station, College of Agriculture and Veterinary Medicine, Qassim University in 2009. A pure culture of the predatory mite was maintained on the two-spotted spider mite Tetranychus urticae Koch, reared on eggplant leaves under laboratory conditions of 26±1°C, 65±3% RH.
Biology of T. swirskii: For rearing experiments, leaf discs of eggplant (2.5 cm in diameter) were used and bordered with tangle foot of a mixture of canada balsam, citronella oil and castor oil to prevent mites from escaping (Krantz, 1978; Fouly, 1997). Five of these discs were placed together on a layer of cotton wool soaked in water in an Petri-dish. Suitable moisture was maintained daily to the cotton layer. Adults of T. swirskii were sexed where mite females were left to deposit eggs. Newly deposited eggs of the same age were divided into three groups (treatments) each of which was approximately 30 eggs, where the newly hatched larvae of the first group was singly fed on 4 eggs of B. tabaci per day, while the second and third groups were offered 8 and 12 eggs day-1 during their life span. Consumed insect eggs were daily counted and replaced by new ones. Duration and rate of mortality of immature stages as well as number of consumed prey eggs and number of deposited eggs by adult females of T. swirskii were recorded daily.
Concerning the crowding effect of T. swirskii on its feeding capacity and egg production during 10 days in oviposition period of mite females, other three groups of 1, 2 and 4 newly emerged and mated adult females (similar in age), each was placed on similar leaf discs and subjected to a surplus amount of insect eggs (approximately 8 eggs each). Consumed whitefly eggs were daily counted and replaced by new ones. Total rate of consumed eggs of B. tabaci as well as total number of deposited eggs of T. swirskii and daily rate/female were counted daily.
Statistical analysis: Data was statistically analyzed by ANOVA test to compare means (LSD test, where p>0.05). Duration of immature stages, mortality, sex ratio and total number of deposited eggs/females of T. swirskii were counted daily and used in calculation of life table parameters. Life table parameters of T. swirskii fed on three level of prey eggs were calculated according to Birch (1948) and by using the Basic Computer Program (Abou Setta et al., 1986) where the intrinsic rate of natural increase, rm was estimated from the equation:
where, x is the age in days, lx the age-specific survival rate (proportion of females alive at age x) x (survival rate during the immature stage) X (hatchability%) and mx the oviposition rate at age x {(age-specific oviposition) x (proportion of females)}.
The net reproductive rate (Ro), is given as:
The mean generation time (T), in days, is given by:
The hatchability and developmental rate at lab conditions of 26°C and 65% RH were used for lx. The proportions of females (number of females/females+males) were used for calculating the mx values.
RESULTS
Development and feeding capacity of T. swirskii: Incubation period of eggs ranged from 1.6 to 2.4 days under laboratory conditions of 26°C and 70% RH. The response of immature stages and adults of T. swirskii to three food level of B. tabaci was studied. Data in Table 1 showed that feeding on surplus amount of prey eggs (8-12 prey eggs day-1) shortened the duration of the predator's developmental stages, which significantly differed from those fed on 4 eggs day-1 (Table 1).
| Table 1: | Duration (days) and feeding capacity of female immature stages of Typhlodromips swirskii fed on 4, 8 and 12 eggs day-1 of whitefly Bemisia tabaci at 26°C and 70% RH |
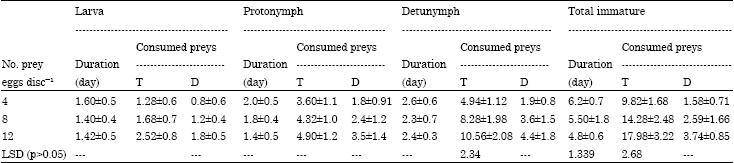 | |
| Values are present as Mean±SE; T: Total consumed preys; D: Daily rate of consumed preys | |
| Table 2: | Duration (days) and feeding capacity during preoviposition, postoviposition, longevity and life span of Typhlodromips swirskii fed on 4, 8 and 12 eggs day-1 of whitefly Bemisia tabaci at 26°C and 70% RH |
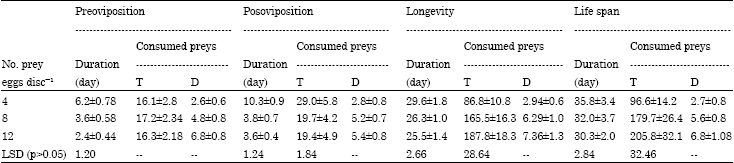 | |
| Values are present as Mean±SE; T: Total consumed preys; D: Daily rate of consumed preys | |
Data also showed that there were no significant differences between total developmental time of immature stages offered neither with 4 and 8 eggs; nor 8 and 12 eggs while the differences were significant between those provided with 4 and 12 prey eggs. On the other hand, data proved that there were no significant differences between the number of devoured prey eggs by larvae and protonymphs of T. swirskii, while deutonymphs significantly ate higher quantity of prey eggs. Similar trend was observed for the total number of consumed prey eggs during the whole total duration of immature development (Table 1). Duration and feeding capacity of T. swirskii during preoviposition and postoviposition periods were significantly affected by number of prey eggs offered. Data in Table 2 showed that during adult female longevity of 29.6, 26.3 and 25.5 days, predatory mite consumed an average of 86.8, 165.5 and 187.8 prey eggs with a daily rate of 2.94, 6.29 and 7.36 prey eggs when each adult female was provided with 4, 8 and 12 prey eggs day-1, respectively. Therefore, the results clearly showed that the duration and feeding capacity during adult female longevity were significantly differed between predatory females provided with 4 prey eggs and those provided with both levels of 8 and 12 prey eggs day-1 (LSD = 2.66 and 28.64). Similar trend was observed during life span of female predatory mite as shown in Table 2.
Table 3 showed that food level offered to the predatory female significantly affected the duration, feeding capacity and fecundity during the ovipositional period. During a period of 13.1, 18.9 and 19.5 days, adult female of T. swirskii consumed an average of 3.2; 6.8 and 7.2 of B. tabaci eggs day-1 when each female was provided with 4, 8 and 12 prey eggs day-1, respectively. Table 3 showed that increasing number of prey eggs offered to the predatory adult female resulted in a higher rate of fertility or egg production.
Life table parameters of T. swirskii: Calculating the life tables of T. swirskii, Table 4 clearly proved that food quantity offered highly affected all life table parameters of the predatory mite under laboratory conditions.
| Table 3: | Duration (days), feeding capacity and egg production during oviposition period of Typhlodromips swirskii fed on eggs of Bemisia tabaci at 26°C and 70% RH |
 | |
| Values are present as Mean±SE; T: Total consumed preys; D: Daily rate of consumed preys | |
| Table 4: | Life table parameters of Typhlodromips swirskii fed on 4, 8 and 12 eggs of Bemisia tabaci at 26°C and 70% RH |
 | |
Data in Table 3 and Fig. 1a-c clearly showed that sex ratio was not affected by food quantity, where females proportion (No. females/No. females+males) averaged 0.68; 0.72 and 0.70 when predatory mites were provided with 4, 8 and 12 prey eggs of B. tabaci, respectively. These values were subsequently used to calculate the specific rate of fecundity (Mx). It is necessary to note that survival curves (Lx) of T. swirskii fed on the aforementioned prey quantities followed a type I in which most eggs developed to maturity (87, 92 and 91%, respectively) and most female death occurred gradually after extended ovipositional period. Table 3 also showed that the mean generation time (T) of predatory mite was 17.15 and 17.66 days when mites were provided with 4 and 8 prey eggs day-1, while it was shorter when the predatory mite was subjected to 12 prey eggs day-1 (15.45 days), respectively. As shown in Table 4 and Fig. 1a-c, it was clear that offering the predatory mite with 12 prey egg day-1 caused the highest value of net reproductive rate (Ro = ΣLx Mx) of 22.12 expected female daughters per female. Feeding on 8 prey eggs day-1 resulted in a slightly lower Ro value of 20.98 and then sharply decreased to reach 10.86 expected female progeny/female, respectively. That means providing the predatory mite with 4 prey eggs day-1 resulted in Ro about 50% of that obtained by feeding on either 8 or 12 prey eggs day-1. Concerning the intrinsic rate of increase (rm) (female/female/day), quantity of prey mites tested here obviously affected the intrinsic rate of increase of T. swirskii where, it was 0.22 female progeny/female/day when predatory mite was provide with 12 prey eggs day-1 and then declined to 0.17 and 0.14 when predatory mites were provided with 8 and 4 prey eggs day-1, respectively (Table 4). The finite rate of increase erm (λ), which means the population multiplications of predatory mite in a unit of time, data showed that it was at its highest level (1.23) when T. swirskii individuals were provided with 12 eggs day-1 and declined to 1.19 and 1.15 after feeding on 8 and 4 prey eggs day-1, respectively.
Crowding effect of the T. swirskii on its feeding capacity and fecundity: Data showed that crowding of the predatory mite T. swirskii decreased the predaceous potentiality. A single female of T. swirskii devoured an average of 38.6 eggs of B. tabaci with a daily rate of 3.8 eggs. These values were significantly reduced to 24.4 and 22.1 prey eggs with a daily rate of 2.4 and 2.2 eggs when 2 and 4 females of T. swirskii lived together for 10 days of the experiment.
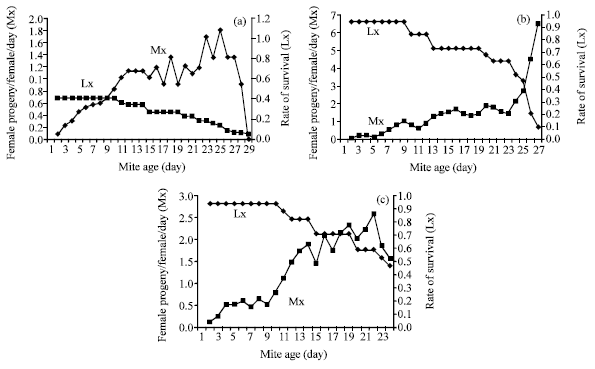 | |
| Fig. 1: | (a-c) Age-specific fecundity (Mx) and survival (Lx) of fed on Typhlodromips swirskii fed on 4, 8 and 12 eggs day-1 of Bemisia tabaci at 26°C and 70% RH |
| Table 5: | Effect of crowding of adult females of Typhlodromips swirskii offered with the same amount of whitefly Bemicia tabaci eggs for 10 days on their feeding capacity and egg production at 26°C and 70% RH |
 | |
| Values are present as Mean±SE; T: Total consumed preys; D: Daily rate of consumed preys | |
Therefore, the egg production was clearly affected by feeding capacity as shown in Table 5. When a single female was kept with a surplus B. tabaci eggs, it laid an average of 12.4 eggs during 10 days. The number of deposited eggs was significantly declined to 6.4 and 4.2 eggs female-1 when 2 and 4 adult females of T. swirskii were together and provided with the same number of prey eggs, respectively. Accordingly, it is recommended to provide the predator's culture with surplus amount of food to avoid the competition effect on different biological aspects of the predatory mite.
DISCUSSION
Development and feeding capacity of T. swirskii: The phytoseiid mite T. swirskii individuals were frequently observed feeding on eggs and immature stages of whiteflies as well as the two spotted spider mite T. urticae. These observations are supported by the findings of Fouly and Al-Rehiayani (2010) at the same area. The present results showed that all immature stages including larvae of T. swirskii feed on a few number of prey eggs, despite the availability of prey number offered. Incorporated results were obtained by El-Laithy and Fouly (1992), Fouly and El-Laithy (1992), Fouly et al. (1995), Abou-Setta et al. (1997), Steiner et al. (2003) and Nomikou et al. (2004) who found that larvae of phytoseiid mites feed on different mite and insect preys. Moreover, McMurtry et al. (1970) stated that larval feeding depended on the species studied. Thus, the present observations suggest that non-feeding larval behavior may be an adaptation to avoid sib-cannibalism, which occurs when eggs are oviposited closer together. This phenomenon was intensively studied by Chittenden and Saito (2001). The present study showed that 87, 92 and 91% of the tested newly hatched larvae reached maturity after feeding on 4, 8 and 12 prey eggs, respectively. Similar results were obtained by Mochizuki (2009) who mentioned that 95.7% to 100% of eggs of the phytoseiid mite Gynaeseius liturivorus (Ehara) fed on diets of Frankliniella occidentalis and Thrips palmi hatched and 75 to 95% of hatched larvae developed to adults. During the development of immature stages of T. swirskii, it was found that increasing food quantity shortened the duration time (LSD = 1.339). These results incorporated with those of Fouly and Hassan (1992) who reared Euseius scutalis (A.-H.) on different immature stages of B. tabaci. They found that increasing B. tabaci preys significantly shortened the development of immature stages of E. scuatalis, where it lasted 7.4, 6.2 and 4.9 days when they were provided with 1, 2 and 4 eggs, respectively. Moreover, food level offered to the predatory adult female significantly affected the duration, feeding capacity and fecundity of T. swirskii especially during the oviposition period. In other words, increasing number of prey eggs resulted in a higher rate of fertility or egg production. Therefore, it can be concluded that the duration and feeding capacity during longevity and life span of adult females were significantly differed according to the quantity of food provided. Ovipositional period of T. swirskii was about 2-4 times the generation time according to the number of prey eggs with a maximum ovipositional rate of 1.5 eggs/female/day. Incorporated results were obtained by Yue et al. (1994), Fouly et al. (1995), Escudero and Ferragut (2005), Momen and El-Sawi (2008) and Al-Shammery (2010) who found that ovipositional period of phytoseiid mites E. mesembrins, Neoseiulus californicus, Phytoseiulus persimilis and E. scutalis didn't exceed 3-4 times the generation time. Contradictory, Abou-Setta et al. (1997) found that the ovipositional period was about 5 times greater than generation time of Proprioseiopsis rotendus (Muma) fed on 4 different food sources.
An adult female of T. swirskii consumed an approximately of 47.79, 77.58 and 74.60% only during oviposition period as compared with 89.85, 92.09 and 91.25% food consumption in its entire longevity when the predatory mite was provided with 4, 8 and 12 eggs, respectively. These results agree with those obtained by Sabelis (1981), Fouly et al. (1995), Fouly (1997) and Steiner et al. (2003) who found that an approximately of 65-70% of the total prey protein consumed by a phytoseiid mite were eaten during ovipositional period. This may be due to prey protein required for egg production. In other word, Sabelis (1981) stated that the prey consumption formed a ratio of about 70% of the biomass of deposited eggs by the predatory mite. Thus, the total eggs deposited during the ovipositional period represented about 70% from total protein of consumed preys.
Life table parameters of T. swirskii: Sex ratio of T. swirskii (No. females/No. females+males) was not affected by food quantity where it averaged 0.68, 0.72 and 0.70 when the predatory mite was provided with 4, 8 and 12 prey eggs, respectively. Female proportion values were subsequently used to calculate the specific rate of fecundity (Mx). Similar results were obtained by Mochizuki (2009) who found that the proportion of females of G. liturivorus was 0.759 among the 311 individuals tested after feeding on diets of F. occidentalis eggs and T. palmi larvae. These findings are not incorporated with Abou-Setta et al. (1997) who noticed lower female proportions of 0.57, 0.56, 0.56 and 0.58 when P. rotendus fed on four different food sources. Also, Fouly (1997) found that female progeny in the first generation of Proprioseiopsis asetus (Chant) presented only 52% when fed on E. orientalis. The present results proved that life table parameters of T. swirskii were highly affected by food quantity offered under laboratory conditions. On the other hand, it is necessary to note that survival curves (Lx) of T. swirskii fed on the aforementioned prey quantities followed a type I in which most eggs developed to maturity (87, 92 and 91%, respectively) and most female death occurred gradually after extended ovipositional period. The mean generation time (T) of predatory mite was sub-equal (17.15 and 17.66 days) when mites were provided with 4 and 8 prey eggs day-1, while it sharply decreased when the predatory mite was subjected to 12 prey eggs day-1 (15.45 days). Similar results were obtained by Al-Shammery (2010) who noticed that T. urticae and Euteranychus orientalis Klein as preys caused T time of E. scutalis of 14.70 and 14.88 days, while it was longer when the predatory mite was subjected to date palm red spider mite Olygonychus afrasiaticus Mc Gregor (16.14 days), respectively. Contradictory, Fouly (1997) found that T time averaged only 12.49 days when P. asetus subjected to E. orientalis under laboratory conditions. The greatest number of prey eggs offered, the highest value of net reproductive rate (RO). That means providing the predatory mite T. swirskii with 4 prey eggs day-1 resulted in RO about 50% of that obtained by feeding on either 8 or 12 prey eggs day-1. These observations agree with those obtained by Al-Shemmary (2010) who found that feeding on E. orientalis and O. afrasiaticus resulted in RO values of the phytoseiid mite E. scutalis were about 50% of that obtained by feeding on T. urticae. Moreover, El-Laithy and Fouly (1992) found that RO of E. scutalis and T. swirskii were 17.22 and 22.97. These findings incorporated with those of Emmert et al. (2008) who found that T. urticae was not the favorable prey mite where it caused Ro value of P. asetus 50% lower than feeding on thrips Frankkliniella occidentalis (Pergande) or cattail pollen Typha latifolia L. Contradictory, Escudero and Ferragut (2005) found a significant higher Ro value with Neoseiulus californicus (McGregor) and Phytoseiulus persimilis A.-H. where it reached 49.25 and 45.61 expected female progeny female-1 after feeding on the two-spotted spider mite T. urticae. The present results are not incorporated with Momen and El-Sawi (2008) who provided E. scutalis with eggs of two lepidopteraous insects and obtained the lowest Ro value among phytoseiid species, where it only averaged 10.94 and 5.40 expected females female-1 after feeding on eggs of Spodoptera littoralis Boisduval and Agrotis ipsillon (Hufnagel), respectively. Contradictory, Momen et al. (2009) found that Ro value was 29.04 when T. negevi Swirski was provided with B. tabaci eggs in laboratory experiments.
Concerning the intrinsic rate of increase (rm) (female/female/day), Birch (1948) stated that rm value is the rate of increase of an insect or mite species under specific physical conditions, in unlimited environment where the effects of increasing density don't need to be considered. The present study proved that quantity of prey mites obviously affected the intrinsic rate of increase rm of T. swirskii where it was at its highest value (0.22 female progeny/female/day) when predatory mite was provided with 12 prey eggs day-1 and then declined to 0.17 and 0.14 when predatory mites were provided with 8 and 4 prey eggs day-1, respectively. Contradictory, Fouly (1997) found that rm value of P. asetus fed on E. orientalis was higher and reached to 0.28. Similar results were obtained by Nomikou et al. (2001, 2002) who reported that rm of E. scutalis fed citrus red spider mite Panonychus citri averaged 0.23-0.29 according to the temperature. Kasap and Lu (2004), contradictory, found that rm value of E. scutalis fed P. citri was lower than the previous one where it averaged between 0.16 and increased to 0.29 female/female/day by increasing temperature. While, Momen and El-Sawi (2008) found that rm value didn't exceed 0.14 when the same predatory mite fed eggs of cotton leaf worm S. littoralis. In general, Abou-Setta et al. (1997), mentioned that the number of deposited eggs of P. rotendus over a period equal to one or two generation times was responsible for most of the calculated rm value.
The finite rate of increase erm (λ), which means the population multiplications of predatory mite in a unit of time, the present results showed that erm ranged from 1.15 to 1.23 according the number of prey eggs offered to T. swirskii. These results incorporated with those obtained by El-Laithy and Fouly (1992), who found that erm of E. scutalis fed T. urticae was 1.15, while this value was only 1.106 when Momen and El-Sawi (2008) reared E. scutalis on eggs of the cotton leaf worm S. ipsillon. In 1997, Fouly found that erm of P. asetus was 1.33 after feeding on E. orientalis, while Al-Shammery (2010) obtained erm value of 1.24 when E. scutalis individuals fed on immature stages of T. urticae and declined to 1.192 and 1.175 after feeding on E. orientalis and O. afrasiaticus, respectively.
Crowding effect of the T. Swirskii on its feeding capacity and fecundity: Crowding of T. swirskii significantly decreased feeding capacity and egg deposition. That may be due to the competition occurring between the predators on each tested leaf discs especially when they were provided with the same quantity of food. These findings incorporated with those obtained by Fouly and Hassan (1992), Fouly (1997), McMurtry and Croft (1997), Nomikou et al. (2004), Inbar and Gerling (2008) and Mochizuki (2009) who noticed that although, the amount of food was surplus, yet predator mite crowding highly decreased feeding capacity and fecundity of the phytoseiid mites E. scutalis (A.-H.); Typhlodromus athiasae (Porath and Swirski), N. barkeri Hughes, T. swirskii; Phytoseius finitimus Ribaga and Gynaeseius liturivorus, respectively. They concluded that this competition may usually happen especially in prey's scarcity in special circumstances in nature.
From the previous results, it can be concluded that comparison with rm value of the insect pest B. tabaci obviously indicates that T. swirskii should be capable to suppress local populations of whitefly under greenhouse or/and open field conditions. Therefore, this predatory mite could be a useful biological control agent against whiteflies under the local conditions in Al-Qassim region. More studies with regard to biology, feeding habits, host range of the predatory mites, especially those in family Phytoseiidae inhabit different localities Saudi Arabia, are needed.
REFERENCES
- Abdel-Baky, N.F. and M.A. Al-Deghairi, 2008. Role of host plants on the biological aspects and parasitism levels of Eretmocerus mundus mercet (Hymenoptera: Aphelinidae), a parasitoid of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). J. Entomol., 5: 356-368.
CrossRefDirect Link - Al-Deghairi, M.A., 2009. Combining effect of Beauveria bassaania (Bals.) and Ertemocerus mundus mercet (Hymenoptera: Aphelinidae) on sweetpotato whitefly, Bemisia tabaci gennadius (Aleyrodidae; Hemiptera). J. Entomol., 6: 72-81.
CrossRefDirect Link - Al Rehiayani, S.M. and A.H. Fouly, 2005. Cosmolaelaps simplex (Berlese), a polyphagous predatory mite feeding on root-knot nematode Meloidogyne javanica and citrus nematode Tylenchulus semipenetrans. Pak. J. Biol. Sci., 8: 168-174.
CrossRefDirect Link - Al-Shammery, K.A., 2010. Different biological aspects of the predaceous mite Euseius scutalis (Acari: Gamasida: Phytoseiidae) and the effects due to feeding on three tetranychid mite species in hail, Saudi Arabia. Asian J. Biol. Sci., 3: 77-84.
CrossRefDirect Link - Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26.
CrossRefDirect Link - Brown, J.K., D.R. Frohlich and R.C. Rosell, 1995. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol., 40: 511-534.
CrossRefDirect Link - Carabali, A., A.C. Bellotti, J. Montoya-Lerma and M.E. Cuellar, 2005. Adaptation of Bemisia tabaci biotype B (Gennadius) to cassava, Manihot esculenta (Crantz). Crop Prot., 24: 643-649.
CrossRef - Chittenden, A.R. and Y. Saito, 2001. Why are there feeding and nonfeeding larvae in phytoseiid mites (Acari: Phytoseiidae)? J. Ethol., 19: 55-62.
CrossRef - Dong, C., J. Zhang, W. Chen, H. Huang and Y. Hu, 2007. Characterization of a newly discovered China variety of M. anisopliae (M. anisopliae var. dcjhyium) for virulence to termites, isoenzyme and phylogenic analysis. Microbiol. Res., 162: 53-61.
PubMed - Emmert, C.J., F. Russell, I. Mizell, P.C. Andersen, J.H. Frank and J. Stimac, 2008. Effects of contrasting diets and temperatures on reproduction and prey consumption by Proprioseiopsis asetus (Acari: Phytoseiidae). Exp. Applied Acarol., 44: 11-26.
CrossRef - Escudero, L.A. and F. Ferragut, 2005. Life history of predatory mites Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae) on four spider mite species as prey, with special reference to Tetranychus evansi (Acari: Tetranychidae). Biol. Control, 32: 378-384.
CrossRef - Fouly, A.H., 1997. Effects of prey mites and pollen on the biology and life tables of Proprioseiopsis asetus (Chant) (Acari, Phytoseiidae). J. Applied Entomol., 121: 435-439.
CrossRefDirect Link - Fouly, A.H. and A.Y.M. El-Laithy, 1992. Immature stages and life history of the predatory mite species Amblyseius barkeri (Hughes, 1948) (Acarina, Gamasida, Phytoseiidae). Deutsche Entomol. Zeitschrift, 39: 427-435.
CrossRefDirect Link - Fouly, A.H., M.M. Abou-Setia and C.C. Childers, 1995. Effects of diet on the biology and life tables of Typhlodromalus peregrinus (Acari: Phytoseiidae). Environ. Entomol., 24: 870-877.
Direct Link - Inbar, M. and D. Gerling, 2008. Plant-mediated interaction between whiteflies, herbivores and natural enemies. Ann. Rev. Entomol., 53: 431-448.
Direct Link - Kasap, I. and E.E. Lu, 2004. Life history of Euseius scutalis feeding on citrus red mite Panonychus citri at various temperatures. BiolControl, 49: 645-654.
Direct Link - Lin, K., K. Wu, Y. Zhang and Y. Guo, 2007. Overwintering and population dynamics of Bemisia tabaci biotype B in greenhouse during the spring in Northern China. Crop Prot., 26: 1831-1838.
CrossRef - McMurtry, J.A. and B.A. Croft, 1997. Life styles of phytoseiid mites and their roles in biological control. Ann. Rev. Entomol., 42: 291-321.
CrossRef - Messelink, G.J., S.E.F. van Steenpaal and P.M.J. Ramakers, 2006. Evaluation of phytoseiid predators for control of western flower thrips on greenhouse cucumber. Biol. Control, 51: 753-768.
CrossRef - Mochizuki, M., 2009. Development, reproduction and prey consumption of the thrips predator Gynaeseius liturivorus (Ehara) (Acari: Phytoseiidae). J. Acarol. Soc. Japan, 18: 73-84.
CrossRef - Momen, F.M. and S. El-Sawi, 2008. Life-history traits of the predacious mite Euseius scutalis (Athias-Henriot) (Acari: Phytoseiidae) on eggs of three insects (Lepidoptera: Noctuidae). Acta Phytopathol. Entomol. Hungarica, 43: 163-170.
CrossRefDirect Link - Momen, F., A. Abdel-Khalek and S. El-Sawi, 2009. Life tables of the predatory mite Typhlodromus negevi feeding on prey insect species and pollen diet (Acari: Phytoseiidae). Acta Phtopathol. Entomol. Hungarica, 44: 353-361.
CrossRef - Nomikou, M., A. Janssen, R. Schraag and M.W. Sabelis, 2001. Phytoseiid predators as biological control agents for Bemisia tabaci. Exp. Applied Acarol., 25: 271-291.
CrossRef - Nomikou, M., A. Janssen, R. Schraag and M.W. Sabelis, 2002. Phytoseiid predators suppress populations of Bemicia tabaci on cucumber plants with alternative food. Exp. Applied Acarol., 27: 57-68.
CrossRef - Nomikou, M., A. Janssen, R. Schraag and M.W. Sabelis, 2004. Vulnerability of Bemicia tabaci immature to phytoseiid predators: Consequences for oviposition and influence of alternative food. Entomol. Exp. App., 110: 95-102.
CrossRef - Oliveira, M.R.V., T.J. Henneberry and P. Anderson, 2001. History, current status and collaborative research projects for Bemisia tabaci. Crop Prot., 20: 709-723.
CrossRefDirect Link - Perring, T.M., 2001. The Bemisia tabaci species complex. Crop Prot., 20: 725-737.
CrossRefDirect Link - Steiner, M.Y., S. Goodwin, T.M. Wellham, I.M. Barchia and L.J. Spohr, 2003. Biological studies of the Australian predatory mite Typhlodromips montdorrensis (Schicha) (Acari: Phytoseiidae), a potential biocontrol agent for western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Aust. J. Entomol., 42: 124-130.
Direct Link - Islam, M.T. and R. Shunxiang, 2007. Development and reproduction of Bemisia tabaci on three tomato varieties. J. Entomol., 4: 231-236.
CrossRefDirect Link - Yue, B., C.C. Childers and A.H. Fouly, 1994. A comparison of selected plant pollens for rearing Euseius mesembrinus (Acari: Phytoseiidae). Int. J. Acarol., 20: 103-108.
CrossRefDirect Link - Abou-Setta, M.M., R.W. Sorrell and C.C. Childers, 1986. Life 48: A basic computer program to calculate life table parameters for an insect or mite species. Florida Entomol., 69: 690-697.
CrossRefDirect Link
Peter Dennis Msa Reply
Quite resourceful