
I.K. Olayemi
Department of Biological Sciences, Federal University of Technology, Minna, Nigeria
A.T. Ande
Department of Biological Sciences, Federal University of Technology, Minna, Nigeria
G. Danlami
Department of Zoology, University of Ilorin, Ilorin, Nigeria
U. Abdullahi
Department of Biological Sciences, Federal University of Technology, Minna, Nigeria
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 5 | Page No.: 459-467







ABSTRACT
This study elucidated the influence of blood meal type on reproductive performance of Anopheles gambiae s.s. Starting from three days old, An. gambiae mosquitoes, from a laboratory colony maintained following standard anopheline rearing techniques, were offered seven different blood meal types, through an artificial membrane. Thereafter, the mosquitoes were monitored for pre-larval and immature stage reproductive attributes. The results indicated that except for Embryony Period (EP) and Pupal Stage Duration (PSD), all attributes investigated varied significantly (p<0.05) among blood meals. While Blood Feeding Rate (BFR) was significantly highest and least in groups of mosquitoes fed with human (Mean = 94.69±3.36%) and pig (Mean = 48.54±10.13%) blood meals, respectively; the shortest Pre-oviposition Period (POP) were recorded among mosquitoes given human and cattle blood meals (Mean = 3.96±0.88 and 4.30±1.25 days, respectively). Also, human blood meal was the most ideal for Oviposition Rate (OR) (Mean = 79.20±12.39%) while chicken and pig blood meals were the least favourable (Mean = 46.80±9.55 and 48.05±6.34%, respectively). Fecundity (F) had similar distribution pattern as BFR while Egg Hatching Rate (HER) was generally low (<60%), irrespective of blood meal type. While Total Immature Stage Duration (TISD) varied widely among blood meal types, Total Larval Stage Duration (TLSD) was only significantly (p<0.05) longest in mosquitoes fed with pig blood meal (12.33±1.63 days). The epidemiological implications of these results were highlighted and discussed.
PDF Abstract XML References Citation
Received: February 28, 2011;
Accepted: April 19, 2011;
Published: May 30, 2011
How to cite this article
I.K. Olayemi, A.T. Ande, G. Danlami and U. Abdullahi, 2011. Influence of Blood Meal Type on Reproductive Performance of the Malaria Vector, Anopheles gambiae s.s. (Diptera: Culicidae). Journal of Entomology, 8: 459-467.
DOI: 10.3923/je.2011.459.467
URL: https://scialert.net/abstract/?doi=je.2011.459.467
DOI: 10.3923/je.2011.459.467
URL: https://scialert.net/abstract/?doi=je.2011.459.467
INTRODUCTION
Mosquito-borne diseases such as malaria, encephalitis, Rift Valley fever, dengue fever, etc are a major public health threat in the tropics (Azari-Hamidian et al., 2011). Malaria, for example, continues to ravage the tropical and sub-tropical regions of the world without a close end in sight; such that even under the most optimistic scenario, the disease may continue to be a major public health threat in these regions for decades. According to USAID (2005), malaria is endemic in over 100 countries where it threatens the lives of over 3 billion people worldwide and is responsible for an estimated 900 million clinical cases of illness and almost 3 million deaths annually, as well as, accounts for an estimated global loss of about 44.7 million Disability Adjusted Life Years (DALYs). Sub-Saharan Africa is the worst hit with malaria health and socio-economic burdens where about 90% of the world’s malaria-induced deaths are recorded and is responsible for 20% of Africa’s infant mortality and 10% overall disease burden (Roll-Back-Malaria, 2002; WHO, 2003; Njan-Nloga et al., 2007). Shah (2005) puts malaria-induced child mortality rate in Africa at one every 30 sec.
The development of an effective vaccine for malaria has remained a mirage and chemotherapy which proved very effective for more than three decades, is now being rendered virtually ineffective in most parts of the world. The failure of chemotherapy has been attributed to the emergence and rapid spread of drug-resistant plasmodial parasites. This development has renewed stakeholders’ interest in vector control, as the most viable strategy for reducing the present unacceptably high burden of malaria in its areas of distribution (McCaffery and Nauen, 2006). Effective malaria vector control however, requires a sound knowledge of the biology, ecology and behaviour of the anopheline species involved in transmission. In sub-Saharan Africa, the main malaria vectors are Anopheles gambiae and An. funestus complexes (Kelly-Hope, 2009; Craig et al., 1999; Appawu et al., 2004). Of these two anopheline complexes, the sibling species An. gambiae s.s. has been adjudged the most efficient vector of human malaria due to its highly anthropophilic, endophagic and endophilic tendencies, as well as its ability to support the development of Plasmodium parasites (Alavi et al., 2003). The vectorial capacity of this mosquito is influenced by factors including ecological, climatic, behavioural, etc. Though most of these factors have been well-studied (Bockarie et al., 1994; Awolola et al., 2003; Koenraadt et al., 2004; Olayemi and Ande, 2008), published information directly relating the behavior of mosquitoes, especially blood meal quality and quantity, to reproductive performance are scanty (Ferdouse and Islam, 2005; Basseri et al., 2010).
According to Briegel (1990), blood-feeding is pivotal to a female mosquito’s reproductive success, as most species require blood meal to obtain amino acids from erythrocytes and plasma protein digestion to synthesize yolk proteins for egg production (Hurd, 2003; Roitberg and Gordon, 2005). Laboratory studies have shown fecundity in mosquitoes to vary with source and size of the blood meal with different blood meals resulting in significant variations in the number of females that developed eggs and number of eggs per female (Taylor and Hurd, 2001; Roitberg and Gordon, 2005). These variations have been attributed to differences in proteins and their constituent amino acids, in the blood meals (Prasad, 1987).
However, blood feeding may result in the ingestion of plasmodial parasites which the mosquito can transmit during subsequent blood feeding. Thus, hosts with superior blood quality for egg production may be preferred by anophelines thereby increasing the frequency at which they feed on such hosts and hence, malaria transmission. Burkot (1988) observed that the prevalence of malaria in an area is greatly influenced by the process of host selection by anopheline vectors which in turn, is influenced by many factors including host preferences of the vectors, dictated by the quality of the host’s blood. In Africa where An. gambiae complex is highly anthropophilic and An. arabiensis is an opportunistic species feeding predominantly on man but diverts to animals when the need arises, the latter species has significantly lower sporozoite rates than the former (Taylor et al., 1990; Githeko et al., 1993, 1994).
From the foregoing, it is clear that an investigation into the influence of blood meal types on reproductive performance of An. gambiae s.s. is imperative, especially, as the quantitative relationships are still poorly understood. These information will, no doubts, shed more light on the evolution of blood meal preferences in anophelines, as well as, the epidemiological implications of such behaviour. This study was therefore, carried out to elucidate the influence of conventional anopheline blood meal sources on pre-larval and immature stage reproductive attributes of An. gambiae s.s.
MATERIALS AND METHODS
Source of mosquitoes and insectary environmental conditions: Day old adult An. gambiae s.s. mosquitoes were obtained from a colony, in its 8th generation, maintained in the laboratory of the Department of Biological Sciences, Federal University of Technology, Minna, Nigeria. The mosquitoes were held in adult rearing cages (60x60x60), stocked at the rate of approximately 150 males and 150 females per cage and transferred to the insectary. Prio to transfer to the insectary, the mosquitoes were maintained in a humid and cool environment (El-Badry and Al-Ali, 2010). The Mean temperature and relative humidity of the insectary, during the study period were 28.57±1.06°C and 63.29±2.82%, respectively. The whole study was carried out and replicated between the months of August 2008 and March 2009.
Mosquito rearing and maintenance: The mosquitoes were reared following standard techniques (Das et al., 2007; Olayemi and Ande, 2009). Blood meal administration were according to the techniques of Lutomiah et al. (2007) and Moutailler et al. (2007) though with slight modifications. Briefly, the blood types were preserved by the addition of 0.3% Sodium Citrate and stored at 5°C and later fed to the adult mosquitoes using a prophylactic membrane (Gold Circle). The membranes were stretched and fastened to a 10 cm hood over a supporting platform of netting material. The hood was then placed on top of the mosquito cages and the preserved blood maintained at 40°C was poured into the membrane for the mosquitoes to feed on, between the hours of 19:00 and 22:00. After each blood meal feeding exercise, the female mosquitoes that successfully fed, evidenced by the red-coloration engorgement of the abdomen were removed from the cages and maintained individually in 20 mL plastic vials, with about 5 mL of distilled water at the bottom to act as oviposition site. Also, similar number of male mosquitoes, as those of the blood fed females were removed from the cages after each feeding exercise, to ensure a constant sex ratio of 1:1 (Male: Female).
After oviposition, the eggs from the vials were transferred into plastic bowls where they were held for 12 to 72 h for hatching. Twenty four hours post-hatching, the larvae were transferred to larval rearing troughs (30x25x5 cm) half-filled with well water, at the rate of 50 larvae/trough. The larvae were fed with fish feed (Tetramin) at the rate of 0.32 mg/larva every other day when the water from the larval rearing troughs were changed carefully until pupation. The pupae were separated from the larvae daily and placed in plastic bowls (5 height and 20 cm diameter) half-filled with well water, at the rate of 50 pupae/bowl. The pupae plastic bowls were placed in adult-holding cages for imago emergence.
Investigation of reproductive performance: Blood Feeding Rate (BFR) of the mosquitoes, defined as the proportion of female mosquitoes that successfully blood-fed was determined as the proportion of female mosquitoes presented with a blood meal type that successfully fed. Pre-Oviposition Period (POP) defined as the interval preceding egg deposition, was estimated as the interval between adult emergence and commencement of egg-laying. Oviposition Rate (OR) defined as the proportion of blood-fed female mosquitoes that oviposited was determined as the proportion of engorged mosquitoes that eventually laid eggs. Fecundity (F) was determined as the total number of eggs laid per mosquito. Embryony Period (EP), defined as egg incubation period was estimated as the interval between oviposition and larval emergence. Egg Hatching Rate (EHR) was estimated from the proportion of eggs hatched per mosquito.
Duration of the immature stages was determined according to the techniques of Edillo et al. (2004) and Olayemi and Ande (2009). Mean larval instar duration and pupal stage duration, in days were estimated using the formula:
where, Di is duration of life stage, Ti is present Mean age and ti-1 is previous mean age at molting.
Statistical analyses: Statistical analyses for data collected were carried out using SPSS computer software for Windows, Version 15.00 (Install Shield Corporation, Inc.). One way Analysis of Variance (ANOVA) was done to calculate the significance level among means of reproductive attributes, due to blood meal types.
RESULTS
Pre-larval development reproductive performance: Table 1 shows the pre-larval development responses of An. gambiae s.s. to different blood meal types. Blood Feeding Rate (BFR) by the mosquitoes varied significantly (p<0.05) with the mosquitoes showing distinct preference for human blood meal (Mean = 94.61±3.36%) while blood from pig was the least favoured (Mean = 48.54±10.13%). The Pre-Oviposition Period (POP) was significantly shorter in mosquitoes fed with human and cattle blood meals (Mean = 3.96±0.88 and 4.30±1.25 days, respectively) but not significantly (p>0.05) in individuals fed the other five types of blood meal (range = 5.13±1.77 to 6.82±2.00 days). Oviposition Rate (OR) also varied significantly (p<0.05) across the blood meal types. As the case with the previous reproductive attributes, OR was most favoured by human blood meal (Mean = 79.20±12.39%) although this was not significantly different from the OR recorded for mosquitoes exposed to dog blood meal. On the other hand, the groups of mosquitoes fed with chicken and pig blood meals had the least number of individuals that oviposited (Mean = 46.80±9.55 and 48.05±6.34%, respectively). As recorded for POP, OR was not significantly different in the groups of mosquitoes given bovine blood meals namely, cattle, goat and sheep meals laid the highest number of eggs (Mean = 103.86±22.91 eggs/female) followed by mosquitoes (range = 69.49±18.00 to 75.00±8.00%).
| Table 1: | Influence of blood meal type on pre-larval reproductive performance of Anopheles gambiae s.s |
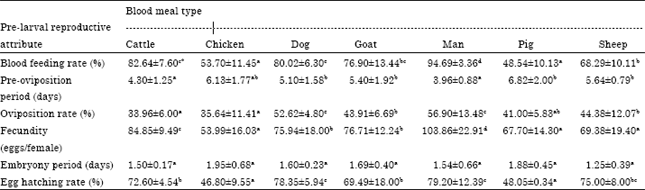 | |
| Values followed by same superscript letter in a row are not significantly different at p = 0.05 level of significance | |
| Table 2: | Mean duration of immature life stages of Anopheles gambiae s.s. in response to blood meal type |
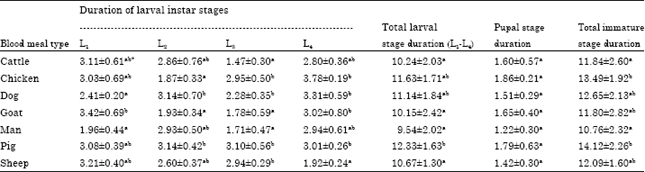 | |
| Values followed by same superscript letter in a row are not significantly different at p = 0.05 level of significance. All values were measured in days | |
Fecundity (F) had a similar pattern of distribution across the blood meal types, as that observed for BFR. Significantly, mosquitoes exposed to human blood fed with cattle, goat and dog blood meals (range = 75.94±18.00 to 84.85±9.49 eggs/female) while mosquitoes given chicken, pig and sheep blood were the least productive (range = 53.99±16.03 to 76.71±12.24 eggs/female). Embryony Period (EP) had a distinct pattern of distribution across the blood meal types, been the only pre-larval reproductive attribute that did not differ significantly (p>0.05) among the blood meal types and where human blood meal was not the best performing type; i.e., 1.54±0.60 days as against 1.25±0.39 days recorded for sheep blood meal type. Egg Hatching Rate (EHR) in the mosquitoes was generally low (<60%) irrespective of blood meal type. However, OR was significantly highest in mosquitoes fed with human and dog blood meals (Mean = 56.90±13.48 and 52.62±4.80%, respectively) and lowest in mosquitoes given cattle and chicken blood meals (Mean = 33.96±6.00 and 35.64±11.41%, respectively).
Immature stage duration: Table 2 shows the duration of immature life stages of An. gambiae s.s. fed with different blood meal types. Total Immature Stage Duration (TISD) varied significantly (p<0.05) across blood meal types, ranging from 10.76±2.32 days in mosquitoes given human blood meal to 14.12±2.26 days in those exposed to pig blood meal. Again, TISD elicited by the bovine blood meals were insignificantly different (p>0.05) and likewise those of human and dog blood meals. Pupal Stage Duration (PSD) was not significantly different among the blood meal types, ranging from 1.22±0.30 days in mosquitoes fed with human blood meal to 1.86±0.21 days in those given chicken blood meal. On the other hand, Total Larval Stage Duration (TLSD) was significantly longest in mosquitoes fed with pig blood meal (12.33±1.63 days) while those of the other blood meal types were insignificantly different (range = 9.54±2.02 days in human blood meal to 11.63±1.71 days in chicken blood meal). The distribution of the individual larval life stage duration across the blood meal types also varied significantly (p<0.05) with the L1 stage been the longest for most of the blood meal types.
DISCUSSION
The results of this study revealed that blood meal is indispensable for successful reproduction in An. gambiae s.s., as none of the mosquitoes denied access to blood meal oviposited. This result agrees with those of Takken et al. (1998) and Klowden and Russell (2004) and confirms that the species is truly an autogenous, i.e., depends on a blood meal for the necessary nutrients to produce eggs. Also, the mosquito showed distinct preferences for certain blood meal types, especially, human blood. Similar results have been obtained elsewhere thus, resulting in the classification of the species as pre-dominantly anthropophagic (Takken and Lindsay, 2009). This feeding behavior stands An. gambiae s.s. in good stead as a formidable vector of malaria and other human diseases such as filariasis, as studies have shown that anopheline vectorial capacity is greatly enhanced by the species’ propensity to feed on man (Basseri et al., 2010). However, the fact that the mosquito fed, at least fairly well, on all the other blood types presented to it, indicates plasticity in its feeding behavior. Thus, it may be feasible to successfully divert the species to blood sources other human, as aimed with the widespread distribution of insecticide-treated bed nets with its attendant potential to reduce malaria transmission. The Pre-oviposition Period in the mosquitoes responded commonly to certain blood meal types, with blood meals from the larger mammals namely, human and cattle, resulting in significantly shorter durations. This finding, perhaps, explains why female anophline mosquitoes fly long distance in search of blood meal and commonly breed and rest close to human habitations and cattle sheds thus, having serious epidemiological implications.
The reproductive performance of An. gambiae s.s. was significantly influenced by blood meal type. This finding agrees with previous reports which shows that a strong positive correlation exists between blood meal type and mosquito reproductive output, especially, fecundity and egg-hatching rates (Clements, 1992; Islam and Ferdousi, 1999). Oviposition Rate (OR) was significantly highest in the groups of mosquitoes given human and dog blood meals. This finding, perhaps, confirms the superiority of human blood for oogenesis in mosquitoes. Though, the dog blood meal achieved, more or less, a similar high level of OR as the human blood meal, the former may not enjoy the same status as the latter, since it elicited significantly lower BFR and longer POP in the mosquitoes. The similar influence of bovine blood meals on OR may be due to the close evolutionary relationships among this group of mammals (Vrba and Schaller, 2000) thus, resulting in the sharing of identical blood quality. Fecundity (F) and BFR had more or less similar patterns of distribution, an observation not entirely surprising. If mosquitoes could discriminate among larval breeding habitats as a result of differential prospects for immature developmental and survival rates (Edgerly et al., 1998) they may as well possess innate ability to identify and select blood meal sources with better potentials of supporting egg development and ultimately higher fecundity.
Embryony Period (EP) was not significantly influenced by blood meal type. This finding, perhaps, the conventional believe that blood meal is utilized by mosquitoes primarily for egg development and maturation (Hurd, 2003; Roitberg and Gordon, 2005). According to (WHO, 1975) embryony in mosquitoes is highly influenced by temperature; and this is generally not a serious problem in the Tropics (Paaijmans et al., 2010). The Egg Hatching Rate (EHR) recorded in this study was generally low, the highest been 56.90±13.48% in the mosquitoes fed with human blood. Earlier, Olayemi and Ande (2009) achieved a higher EHR of 63.19±23.91% with the species, by simulating a crepuscular dusk condition to encourage mating by the mosquitoes. However, despite the low EHR, human and dog blood meals elicited significantly better performance in this respect, perhaps, confirming the superiority of particularly human blood meal for egg development in mosquitoes.
The results of this study equally revealed that larval stage developmental rates of An. gambiae s.s. were significantly influenced by blood meal types and their responses were similar to those of pre-larval reproductive attributes. This finding indicates that while blood meal is of primary importance for egg development in An. gambiae s.s., it never-the-less secondarily influences immature stage developmental rates and hence, overall reproductive success in the species. This, no doubts, will influence the vectorial capacity of the mosquito with serious epidemiological consequences for malaria transmission.
CONCLUSION
The results of this study confirms that blood meal is inevitable for successful reproduction in An. gambiae s.s and the source significantly influences pre-larval and immature stage reproductive indices with human blood meal eliciting the highest level of productivity. This, perhaps, explains the dominance of anthropophagy among wild populations of An. gambiae s.s and perhaps, provides a justification for diverting the mosquito to blood meal sources other than man, especially with the use of insecticide-treated bed nets. While this strategy reduces malaria transmission in human populations by reducing vector-human host contacts, it ultimately may not reduce vector density, as the results of this study indicated that An. gambiae s.s. can, to some extent, reproduce successfully utilizing blood meal sources other than human hosts.
REFERENCES
- Alavi, Y., M. Arai, J. Mendoza, M. Tufet-Bayona and R. Sinha et al., 2003. The dynamics of interactions between Plasmodium and the mosquito: a study of the infectivity of Plasmodium berghei and Plasmodium gallinaceum and their transmission by Anopheles stephensi, Anopheles gambiae and Aedes aegypti. Int. J. Parasitol., 33: 933-943.
CrossRef - Azari-Hamidian, S., M.R. Abai, K. Arzamani, H. Bakhshi, H. Karami, H. Ladonni and R.E. Harbach, 2011. Mosquitoes (Diptera: Culicidae) of North Khorasan province, Northeastern Iran and the zoogeographic affinities of the Iranian and middle Asian mosquito fauna. J. Entomol., 8: 204-217.
CrossRefDirect Link - Basseri, H., A. Raeisi, M.R. Khakha, A. Pakarai and H. Abdolghafar, 2010. Seasonal abundance and host-feeding patterns of anopheline vectors in malaria endemic area of Iran. J. Parasitol. Res., 2010: 8-8.
CrossRef - Briegel, H., 1990. Fecundity, metabolism and body size in Anopheles (Diptera: Culicidae), vectors of malaria. J. Med. Entomol., 27: 839-850.
CrossRefPubMedDirect Link - Bockarie, M.J., M.W. Service, G. Barnish, G.H. Maude and B.M. Greenwood, 1994. Malaria in a rural area of Sierra Leone. III. Vector ecology and disease transmission. Ann. Trop. Med. Parasitol., 88: 251-262.
PubMed - Burkot, T.R., 1988. Non-random host selection by anopheline mosquitoes. Parasitology Today, 4: 156-162.
CrossRef - Craig, M.H., R.H. Snow and D. Le-Sueur, 1999. A climate-based distribution model of malaria transmission in sub-Saharan Africa. Parasitol. Today, 15: 105-111.
PubMed - Edillo, F.E., Y.T. Toure, G.C. Lanzaro, G. Dolo and C.E. Taylor, 2004. Survivorship and distribution of immature Anopheles gambiae s.l. (Diptera: Culicidae) in Banambani village, Mali. J. Med. Entomol, 41: 333-339.
PubMed - Edgerly, J.S., M. Mcfarland, P. Morgan and T. Livdahl, 1998. A seasonal shift in egg-laying behaviour in response to cues of future competition in a treehole mosquito. J. Anim. Ecol., 67: 805-818.
CrossRef - El-Badry, A.A. and K.H. Al-Ali, 2010. Prevalence and seasonal distribution of dengue mosquito, Aedes aegypti (Diptera: Culicidae) in Al-Madinah Al-Munawwarah, Saudi Arabia. J. Entomol., 7: 80-88.
CrossRefDirect Link - Githeko, A.K., M.W. Service, C.M. Mbogo, F.K. Atieli and F.O. Juma, 1993. Plasmodium falciparum sporozoite and entomological inoculation rates at the Ahero rice irrigation scheme and the Miwani sugar-belt in Western Kenya. Ann. Trop. Med. Parasitol, 87: 379-391.
PubMed - Githeko, A.K., M.W. Service, C.M. Mbogo, F.K. Atieli and F.O. Juma, 1994. Origin of blood meals in indoor and outdoor resting malaria vectors in western Kenya. Acta Trop., 58: 307-316.
CrossRef - Hurd, H., 2003. Manipulation of medically important insect vectors by their parasites. Annu. Rev. Vector Ecol., 48: 141-161.
CrossRef - Klowden, M.J. and R.C. Russell, 2004. Mating affects egg maturation in Anopheles gambiae Giles (Diptera: Culicidae). J. Vector Ecol., 29: 135-139.
PubMedDirect Link - Koenraadt, C.J.M., A.K. Githeko and W. Takken, 2004. The effects of rainfall and evapotranspiration on the temporal dynamics of Anopheles gambiae s.s. and Anopheles arabiensis in a Kenyan village. Acta Trop., 90: 141-153.
CrossRef - Lutomiah, J.L., C. Mwandawiro, J. Magambo and R.C. Sang, 2007. Infection and vertical transmission of Kamiti river virus in laboratory bred Aedes aegypti mosquitoes. J. Insect Sci., 7: 1-7.
PubMed - Moutailler, S., M. Bouloy and A.B. Failloux, 2007. Efficient oral infection of Culex pipiens quinquefasciatus by Rift Valley fever virus using a cotton stick support. Am. J. Trop. Med. Hyg., 76: 827-829.
Direct Link - Njan Nloga, A.M., P. Saotoing, J.C. Tchouankeu and J. Messi, 2007. Effect of essential oils of six local plants used insecticide on adults of Anopheles gambiae, Giles 1902. J. Entomol., 4: 444-450.
CrossRefDirect Link - Olayemi, I.K. and A.T. Ande, 2009. Life table analysis of Anopheles gambiae (diptera: culicidae) in relation to malaria transmission. J. Vector-borne Dis., 46: 295-298.
PubMedDirect Link - Prasad, R.S., 1987. Nutrition and reproduction in haematophagous arthropods. Proc. Indian Acad. Sci. (Anim. Sci.), 96: 253-273.
CrossRef - Roitberg, B.D. and I. Gordon, 2005. Does the Anopheles blood meal-fecundity curve, curve? J. Vector Ecol., 30: 83-86.
PubMed - Takken, W., M.J. Klowden and G.M. Chambers, 1998. Effect of body size on host seeking and blood meal utilization in Anopheles gambiae sensu stricto (Diptera: Culicidae): The disadvantage of being small. J. Med. Entomol., 35: 639-645.
PubMed - Taylor, P.J. and H. Hurd, 2001. The influence of host haematocrit on the blood feeding success of Anopheles stephensi: Implications for enhanced malaria transmission. Parasitology, 122: 491-496.
PubMed - Taylor, K.A., J.K. Koros, J. Nduati, R.S. Copeland, F.H. Collins and A.D. Brandling-Bennett, 1990. Plasmodium falciparum infection rates in Anopheles gambiae, An. arabiensis and An. funestus in western Kenya. Am. J. Trop. Med. Hyg., 43: 124-129.
PubMed