
F. E. Nwilene
Africa Rice Center (AfricaRice), PMB 5320, Ibadan, Oyo State, Nigeria
A. Onasanya
Africa Rice Center (AfricaRice), 01 BP 2031, Cotonou, Benin Republic
A. Togola
Africa Rice Center (AfricaRice), 01 BP 2031, Cotonou, Benin Republic
O. Oyetunji
Africa Rice Center (AfricaRice), PMB 5320, Ibadan, Oyo State, Nigeria
M. Semon
Africa Rice Center (AfricaRice), PMB 5320, Ibadan, Oyo State, Nigeria
M. Tamo
International Institute of Tropical Agriculture (IITA), 01 BP 2031, Cotonou, Benin Republic
E. O. Bright
National Cereals Research Institute, PMB 8, Bida, Nigeria
S. Ofodile
International Institute of Tropical Agriculture, PMB 5320, Ibadan, Oyo State, Nigeria
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 5 | Page No.: 417-428







ABSTRACT
Rice and maize intercrop is a common feature of traditional upland rice cultivation in Nigeria. Stemborer larvae cause significant yield loss in rice. The study aimed at identifying stemborer resistant status of upland NERICA rice varieties and evaluating the effectiveness of maize as a trap crop to protect upland NERICA rice varieties against stemborers. The resistance status of 7 NERICA rice and 2 other rice varieties to stemborer was evaluated in 2006 and 2007 under natural infestation maize and cassava intercropping systems in the humid forest zone of Nigeria. In 2006 study, NERICA1, NERICA2 and NERICA5 together with the resistant check LAC23 were classified as stemborer resistant (SBR) and NERICA3, NERICA4, NERICA6 and NERICA7 together with susceptible check OS6 were classified as stemborer susceptible (SBS). The SBR varieties (NERICA1 and NERICA2) from the 2006 study intercropped with maize and cassava in 2007 revealed the effectiveness of maize as a trap crop and cassava as a refuge for generalist predators against stemborer damage on upland rice. Maize appeared an effective trap crop for rice stemborers because there was a marked and significant reduction in the stemborer attack on rice in the NERICA rice/maize intercrops (GrB cluster) as compared to the NERICA rice monocrops (GrA cluster) and NERICA rice/cassava intercrops (GrA cluster). Maliarpha separatella Ragonot was the predominant stemborer species on rice followed by Sesamia calamistis Hampson. It was concluded that NERICA1 and NERICA2 could be recommended to farmers in stemborer prone areas and that maize was a suitable trap crop for managing rice stemborers.
PDF Abstract XML References Citation
Received: February 03, 2011;
Accepted: May 02, 2011;
Published: May 30, 2011
How to cite this article
F. E. Nwilene, A. Onasanya, A. Togola, O. Oyetunji, M. Semon, M. Tamo, E. O. Bright and S. Ofodile, 2011. Effect of Intercropping Maize and Cassava with Upland NERICA Rice Varieties on Stemborer Attack in Southwest Nigeria. Journal of Entomology, 8: 417-428.
DOI: 10.3923/je.2011.417.428
URL: https://scialert.net/abstract/?doi=je.2011.417.428
DOI: 10.3923/je.2011.417.428
URL: https://scialert.net/abstract/?doi=je.2011.417.428
INTRODUCTION
Stemborers constitute the most widely distributed and injurious group of the insect pests of rice. They are key pests of rice in West Africa as they are in other rice-growing regions throughout the world (Heinrichs and Barrion, 2004). In fact, they have been implicated as the major constraint to rice production in West Africa (Heinrichs and Barrion, 2004). These stemborers comprise various species of lepidopterous moths and dipterous stalk-eyed, diopsis species (CABI, 2004). The visible symptoms of stemborer damage vary with the growth stage at which plant infestation begins (Baloch et al., 2004). Rice plants from seedling to maximum tillering stages that have been attacked at the base of the stem have deadhearts while plants that have started bearing panicles when they were attacked produce whitehead (IRRI, 2009b). Yield losses between 10-70% have been attributed to stemborers in cereal production in outbreak areas in sub-Saharan Africa (Pascal, 1998; Chabi-Olaye et al., 2005a, b; Ateyim et al., 2005; Setamou et al., 2000). grain damage by total crop failure has also been reported in a few instances (Mulungu et al., 2011; Khan et al., 2003; Pascal, 1998). In addition, lepidopterous stemborers have been reported to predispose cereal crops such as maize to pre-and post-harvest infestations by storage beetles, infections by pathogens such as Aspergillus flavus Link and Fusarium verticillioides (Saccardo) Nirenberg and subsequent contamination by mycotoxins (Oparaeke and Kuhiep, 2006; Cardwell et al., 1997; Setamou et al., 2000).
Many strategies to reduce losses due to stemborers have been practiced. These include use of pesticides, varietal resistance/tolerance, biological control and cultural control measures through the biodiversity of plants. The use of synthetic insecticides can be environmentally disruptive and can result in the accumulation of residues in the harvested product (Dhuyo and Soomro, 2007; Pappas et al., 2011). Also, the risks to health and the environment associated with the use of an insecticide mostly outweigh the benefits. Besides, because the larvae which cause the damage are concealed inside the plant stem, these pests can be very difficult to control with insecticides. Systemic insecticides which are translocated within the plant system would have been the only reliable form of chemical control for stemborers after the borers have entered the rice stem when it is generally too late to save/salvage (Dhyo and Soomro, 2008).
Major emphasis has been given in recent years to the use of habitat management techniques such as crop diversification as a control strategy (Songa et al., 2007). Although this has for long been a prominent feature of small holder crop production in sub-Sahara Africa, its benefits are further promoted by the farmers’ concerns. This will not only enhance farm productivity per unit land but also for ensuring security against the potential risk of monoculture alongside creating a platform for stabilizing the diversified needs of the farming households whose production (by its subsistence nature) is greatly influenced by the vagaries of nature (Tahir et al., 2003; Tsubo et al., 2003). Diversified cropping systems play many roles in the agro-ecosystem, such as protection of plants of individual crop species from their host-specific predators and disease organisms, greater competition towards weeds, improved product quality and reduced negative impact of arable crops on the environment. A considerable number of studies have shown that pest populations are higher, more frequent and cause greater yield losses in monocrops than in more diverse cropping systems (Rami et al., 2002; Otway et al., 2005). Also, many studies in tropical as well as temperate zones reported decreased pest densities in diversified cropping systems (Rami et al., 2002; Thies and Tscharntke, 1999; Kruess and Tscharntke, 2000). The use of habitat management strategies with available natural resources such as wild hosts and non-host plants of stemborers against indigenous stemborer species, can increase the understanding of interactions between pests, their cultivated and wild hosts as well as their natural enemy fauna on both types of hosts plants (Khan et al., 2000). However, farmers are mostly not aware of the beneficial effect that mixed cropping can have on pest infestations.
Most of the traditional Oryza sativa varieties grown in West Africa are highly susceptible to stemborer attack. Africa rice has generated several hundred NERICA lines, opening new gene pools and increasing the biodiversity of rice to farmers. The present study aimed at identifying stemborer resistant status of upland NERICA rice varieties and evaluating the effectiveness of maize as a trap crop to protect upland NERICA rice varieties against stemborers. Maize and rice share some common stemborer species. There is also the need to assess if intercropping upland NERICA rice varieties with cassava can attract generalist predators such as spiders, cassava predatory phytoseiid mites, to reduce stem borer damage on rice plants.
MATERIALS AND METHODS
The study site: The field experiments were conducted under rainfed upland conditions in 2006 and 2007 at Ikenne, southwest Nigeria. The location is a ‘hot-spot’ for stemborers because maize is widely cultivated by farmers in the area. It lies between longitude 6° 32'N and 7° 97'N and latitude 2°67'E and 4°40'E. It has an average annual rainfall of 1480 mm and a monthly mean temperature range of 18-24°C during the rainy season and 30-35°C during the dry season. The rainfall pattern is bimodal and the vegetation type is humid forest. The soil type is a highly acidic ultisol with very low phosphorus content.
Rice varieties and experimental design: In 2006, the reaction of nine rice varieties (Table 1) to stemborer attack was evaluated under natural infestation at Ikenne. The seven NERICA rice varieties were selected because they were the first set of upland NERICA rice varieties released in Nigeria. The experimental design was Randomized Complete Block Design (RCBD) with three replications. There were 9 plots per replicate and each plot measured 2 by 3 m. In 2007 however, only two NERICA rice varieties (NERICAs 1 and 2) with the least level of stemborer infestation from the 2006 study and two control rice varieties were intercropped with maize and cassava (Table 2). There were a total of 14 plots per replicate and each plot measured 5 m by 5 m. The stemborer resistant maize variety, TZP-SR used was obtained from the International Institute of Tropical Agriculture (IITA) and the cassava was a local variety. In both years, rice seeds in the monocrops were directly sown at a spacing of 20 by 20 cm and seedlings later thinned to one per hole. The rice plots were separated by 40 cm alley from the maize. The plant spacing in the maize monocrops was 20 by 60 cm and seedlings later thinned to one per hole. The cassava stem cuttings were buried 10 cm into the ground at a spacing of 50 cm and later trimmed to one plant per hole. A basal fertilizer application of 30 kg ha-1 each of N, P2O5 and K2O was applied before planting followed by top dressings with 20 kg ha-1 of N at the maximum tillering and early panicle initiation stages. Weeding was done at 21 and 42 Days after Sowing (DAS) and subsequently when necessary.
| Table 1: | Identity of rice varieties evaluated for stemborer attack under natural infestation at Ikenne Nigeria in 2006 |
 | |
| Table 2: | Identity of rice varieties with maize and cassava intercropped combinations evaluated for stemborer attack under natural infestation at Ikenne Nigeria in 2007 |
 | |
| RCK: Resistant check, SCK: Susceptible check, TZP-SR: Maize variety name, SBR: Stemborer resistant | |
Assessment of stemborer damage: At 30 and 60 DAS, 20 hills were randomly selected per plot for assessing the intensity of stemborer damage (deadhearts). Scoring for whitehead was carried out at 90 DAS. The scoring and percentage plants with deadhearts and whiteheads were calculated as described in the standard evaluation system for rice stemborers (IRRI, 2009a).
Incidence and abundance of stemborer species: This was carried out following procedure adopted by Duale and Nwanze (1999). Twenty hills were randomly selected per plot and dissected in the laboratory for the species of the stemborer larvae and pupae present. The proportion of each species to the total number of hills was calculated. No insecticide was used in or near the experimental area throughout the study period.
Data analysis: Analysis of variance, mean comparison and unweighted pair group method arithmetic mean (UPGMA) principal component cluster analysis were carried out on % deadheart, % whitehead and % stemborer larvae and pupae (% M. separatella and % S. calamistis) data using SAS statistical software (SAS Institute, 2002-2003).
RESULTS
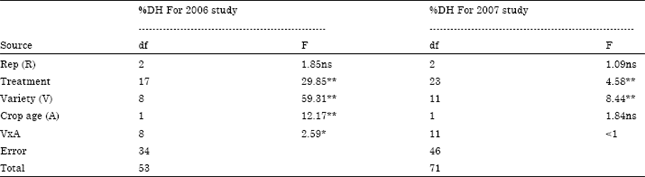
The effect of intercropping maize with upland NERICA rice varieties on stemborer attack in southwest Nigeria has been evaluated. According to ANOVA results, the assessment of stemborer damage (% whitehead and % deadheart) among 7 NERICA rice varieties initially studied in 2006 was highly significant (1% level), meaning that all the 7 NERICA rice varieties were attacked but with different level of resistant status (Table 3). On the basis of ANOVA results, the assessment of stemborer damage (% whitehead, % deadheart, % M. separatella and % S. calamistis) under maize and cassava intercropping condition with NERICA1 and NERICA2 rice varieties studied in 2007 was highly significant (1 and 5% levels), revealing that NERICA1 and NERICA2 rice varieties as well as their maize and cassava intercropping combinations were attacked but with different level of resistant status (Table 3). Besides, in the 2006 study ANOVA results also revealed significant interaction (1% level) between crop age (30 and 60 DAS) and stemborer attack, indicating that at different age the 7 NERICA rice varieties responded differently to stemborer attack (Table 4).
| Table 3: | Analysis of variance for % whitehead, % deadheart, % Maliarpha separatella and % Sesamia calamistis as measure for rice stemborer attack from 2006 and 2007 studies |
 | |
| **Significant at 1% level; *Significant at 5% level; ns: Not significant. WH: Whitehead; DH: Deadheart; M: Maliarpha separatella; S: Sesamia calamistis | |
| Table 4: | Analysis of variance revealing the effect of crop age on rice stemborer attack from 2006 and 2007 studies |
 | |
| **Significant at 1% level; *Significant at 5% level; ns: Not significant. WHWehead; DH: Deadheart | |
| Table 5: | Mean comparison for percentage stemborer damage (deadheart) of nine rice varieties relative to day after sowing without intercropping at Ikenne Nigeria in 2006 wet season. |
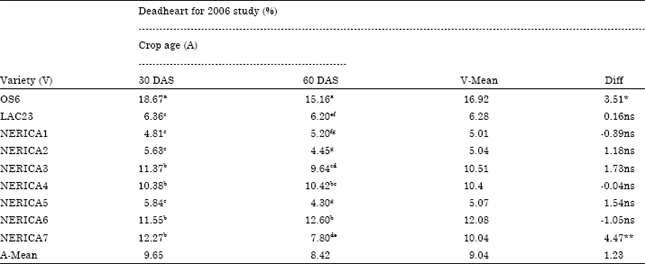 | |
| Means followed by the same letter are not significantly different at p<0.05 Fisher’s least significant difference, **Significant at 1% level, *Significant at 5% level, ns: Not significant, DAS: Day after sowing. | |
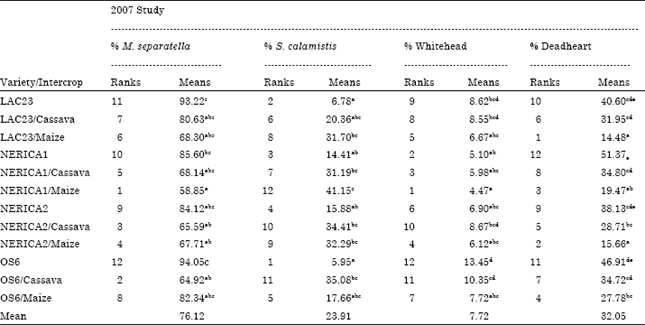
Besides, in the 2007 study ANOVA results revealed no significant interaction between crop age (30 and 60 DAS) and stemborer attack, indicating that at different age under maize and cassava intercropping conditions NERICA1 and NERICA2 rice varieties responded in the same way to stemborer attack in contrary to 2006 study without intercropping (Table 4). The percentage stemborer damage on the 7 NERICA rice varieties at 30 and 60 DAS was between 4.81 to 12.27% and 4.3 to 12.6%, respectively (Table 5). The percentage stemborer damage on the NERICA1 and NERICA2 rice varieties and their maize and cassava intercrops at 30 and 60 DAS was between 7.34 to 21.22% and 8.32 to 30.15% respectively (Table 6). The mean stemborer damage in terms of % whitehead and % deadheart among the 7 NERICA rice varieties was between 5.35 to 14.5 and % 5.01 to 12.06%), respectively (Table 7). The mean stemborer damage in terms of % M. separatella,% S. calamistis, % whitehead and % deadheart, among NERICA1 and NERICA 2 rice varieties and their maize and cassava intercrops was between 58.85 to 85.6%, 14.41 to 41.15%, 4.47 to 8.67% and 15.66 to 51.37%, respectively (Table 8).
The classification of the levels of resistance of the 7 NERICA rice varieties to stemborer damage according to cluster dendrogram was as follows: NERICA1, NERICA2 and NERICA5 together with the resistant check (RCK) LAC23 were classified as stemborer resistant (SBR) and NERICA3, NERICA4, NERICA6 and NERICA7 together with susceptible check (SCK) OS6 were classified as stemborer susceptible (SBS) (Fig. 1). Consequently on the basis of SBR classification, NERICA1 and NERICA2 emerged as the best in terms of stemborer resistance among the 7 NERICA rice varieties studied in 2006 and were used in maize and cassava intercropping study in 2007 to further confirm their resistance status under intercropping condition.
According to principal component cluster dendrogram, the classification of the effects and levels of resistance of NERICA1 and NERICA 2 rice varieties and their maize and cassava intercrops to stemborer damage revealed three major groups (GrA, GrB and GrC) (Fig. 2).
| Table 6: | Mean comparison for percentage stemborer damage (deadheart) of four rice varieties relative to day after sowing with intercropping at Ikenne Nigeria in 2007 wet season |
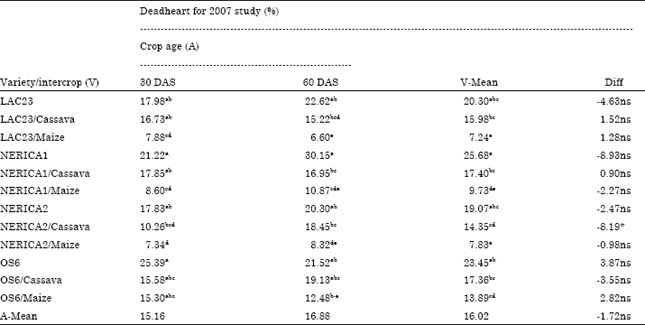 | |
| Means followed by the same letter are not significantly different at p<0.05 Fisher’s least significant difference, *Significant at 5% level, ns: Not significant; DAS: Day after sowing | |
| Table 7: | Mean comparison for percentage stemborer damage (whitehead and deadheart) of nine rice varieties without intercropping at Ikenne Nigeria in 2006 wet season |
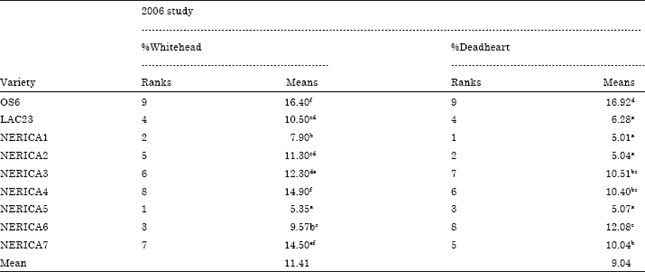 | |
| Means followed by the same letter are not significantly different at p<0.05 Fisher’s Least Significant Difference; DAS = day after sowing | |
| Table 8: | Mean comparison for percentage stemborer damage of four rice varieties with intercropping at Ikenne Nigeria in 2007 wet season |
 | |
| Means followed by the same letter are not significantly different at p<0.05 Fisher’s least significant difference | |
GrA was made up of stemborer resistant (SBR) varieties and their cassava intercrop only (NERICA1, NERICA1/Cassava, NERICA2, NERICA2/Cassava, LAC23 and LAC23/Cassava), GrB formed maize intercrop only of SBR varieties (NERICA1/Maize, NERICA2/Maize and LAC23/Maiza) while GrC constituted the stemborer susceptible (SBS) variety and its maize and cassava intercrops (OS6, OS6/Maize and OS6/Cassava) (Fig. 2). The basis of GrA cluster in terms of stemborer resistance revealed that both NERICA1 and NERICA2 monocrops and their cassava intercrops showed similar response to stemborer damage, whereas GrB cluster revealed both NERICA1 and NERICA2 monocrops and their maize intercrops have different response to stemborer damage.
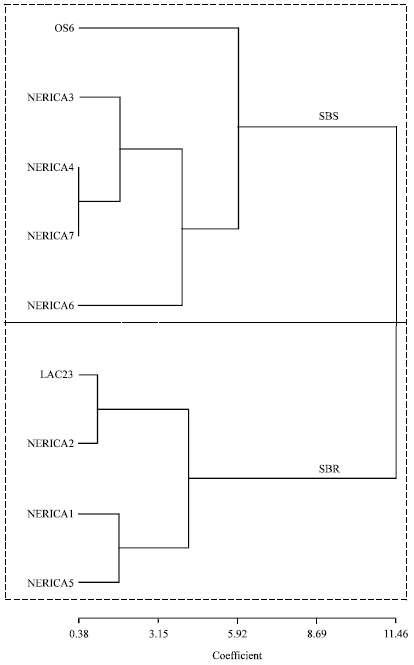 | |
| Fig. 1: | Unweighted pair group method arithmetic mean (UPGMA) principal component cluster showing stemborer resistance status of 7 NERICA rice and 2 other rice varieties in 2006 study |
Therefore, the resistance status of NERICA1 and NERICA2 was further enhanced under maize intercropping condition as critically revealed by GrB cluster (Fig. 2).
DISCUSSION
The current trend in global agriculture which is to search for a highly productive, sustainable and environmentally friendly cropping system has resulted in the renewed interest in cropping systems research (Crews and Peoples, 2004). Two hypotheses, the trap crop and natural enemy hypotheses, postulated by Shelton and Badenes-Perez (2006) were the objectives for the cropping patterns used in the present study.
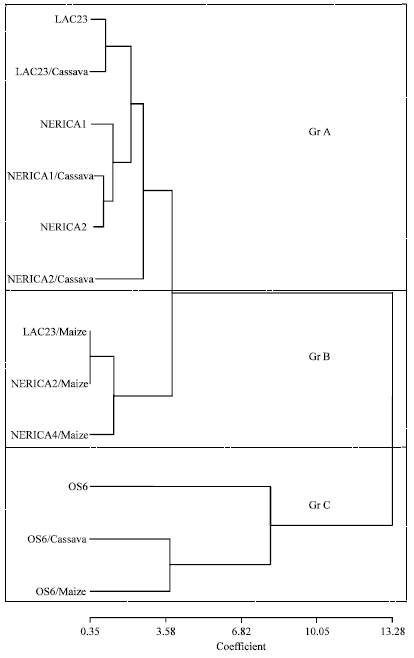 | |
| Fig. 2: | Unweighted pair group method arithmetic mean (UPGMA) principal component cluster showing the effect of rice/maize intercropping on NERICA stemborer attack in 2007 study |
Trap cropping as a control strategy exploits the insects’ preference for certain plant species or growth stages of a plant by attracting the pest species away from a nearby main crop (Shelton and Badenes-Perez, 2006). Recent studies in West Africa showed that mixed cropping involving maize and non-host plants such as legumes or cassava, reduced borer infestations by up to 80% (Schulthess et al., 2004; Chabi-Olaye et al., 2005b). Maize was included in the present study to assess its effectiveness as a trap crop for the rice stemborers. The results obtained in the present study indicated that maize diverted some of the stemborers from attacking upland NERICA 1 and NERICA 2 rice varieties under maize intercropping condition as critically revealed by GrB cluster. In the present study, results further points to maize as an effective trap crop for rice stemborers because there was a marked and significant reduction in the stemborer attack on the rice in the NERICA rice/maize intercrops (GrB cluster) as compared to the NERICA rice monocrops (GrA cluster) and NERICA rice/cassava intercrops (GrA cluster). The reason for the preference could not be farfetched. The sturdier and fuller stems of maize as compared to rice which has a thin and hollow stem could be the reason for the preference for maize as this would provide more readily available food to the stemborer larvae (Heinrichs and Barrion, 2004).
The inclusion of cassava in the study was to observe if it would attract predators such as spiders, cassava predatory phytoseiid mites, etc. and also to observe if the afore-mentioned might have some foraging activity on the stemborers. Results obtained however from the rice/cassava intercrops did not prove this as NERICA rice monocrops and NERICA rice/cassava intercrops formed the same GrA cluster. The reason for this result could be that the cassava variety used for the study does not naturally harbour predators such as spiders and phytoseiid mites and as well does not provide food to the stemborer larvae.
Of the stemborer species that attack cereals in West Africa, four are of major importance on rice which includes the dipterous stalk-eyed fly (Diopsis longicornis Macquart), lepidopterous white stemborer (M. separatella), striped stemborer (Chilo zacconius Bleszynski) and the pink stemborer (S. calamistis) (Rami et al., 2002; Nwilene et al., 2008). Two of the afore-mentioned stemborer species, M. separatella and S. calamistis were observed in the present study to infest upland rice at Ikenne with the population of M. separatella being higher than that of S. calamistis. Previous studies at M’be, Côte d’Ivoire have shown that M. separatella was the most abundant stemborer species in the drought-prone upland and lowland rice ecosystems based on tiller dissections (Heinrichs and Barrion, 2004). In another study carried out at the vegetative, booting and flowering (ripening) stages of rice growth on farmers’ fields in Côte d’Ivoire, M. separatella was also found to be more abundant than S. calamistis at the three stages of the rice growth cycle (Heinrichs and Barrion, 2004). The percent stemborer composition of C. zacconius, M. separatella and S. calamistis in upland and irrigated rice ecosystems in Ibadan, Nigeria revealed M. separatella as predominating in both ecosystems (Nwilene et al., 2008). The higher population of M. separatella notwithstanding, its feeding damage is unique as it seldom causes deadhearts and whiteheads because of its habit of dwelling at the lower internodes (Heinrichs and Barrion, 2004). This possibly suggests that most of the stemborer damage reported on rice in this study could be attributed to S. calamistis since its feeding damage readily results in deadhearts and whiteheads. The population of S. calamistis was generally higher in the NERICA rice/maize intercrops than NERICA rice monocrops and the NERICA rice/cassava intercrop suggesting a preference of S. calamistis for maize. Linnet and Adele (2001) reported populations of S. calamistis to be highest in the upland rice fields near maize, as maize is the preferred host plant. In a study by Songa et al. (2007) on the role of intercropping different cereal species in controlling maize stemborers in Kenya, a high infestation of S. calamistis was reported not only on monocropped millet but also in maize/millet intercrop. It was observed that S. calamistis was predominant on the interspecific NERICA rice varieties while M. separatella was predominant on the O. sativa varieties (LAC 23 and OS 6). This could also suggest a preference of S. calamistis for the interspecific NERICA rice varieties.
CONCLUSION
The NERICA rice varieties are becoming increasingly popular especially amongst farmers for their yield advantage over their O. glaberrima and O. sativa parents through their superior weed competitiveness, drought tolerance, higher yielding potentials and pest and disease resistance. Of the upland NERICAs screened in the present studies, NERICAs 1 and 2 stood out as stemborer resistant varieties. They have therefore, proved to be potentially good varieties in stemborer prone areas and as resistant checks in stemborer studies. Also, the importance of intercropping and diversified cropping systems has been discussed and can not be ignored. Apart from its advantages in insect pest control, diversified cropping systems is also helpful for farmers that have limited access to land and inputs such as fertilizer. In the control of rice stemborers, diversified cropping systems involving maize can be an effective tool against stemborer attack.
ACKNOWLEDGMENTS
The authors gratefully thank Mr. Adebayo Kehinde for technical assistance.
REFERENCES
- Dhyo, A.R. and N.M. Soomro, 2008. Pathogenicity of Beauveria bassiana (Deuteromycota: Hyphomycetes) against the yellow rice stem borer, Scirpophaga incertulas (Lepidoptera: Pyralidae) under laboratory conditions. Pak. Entomol., 30: 37-42.
Direct Link - Ateyim, S.T.S., D. Obeng-Ofori, M.A. Botchey and E.O. Owusu, 2005. Some aspects of the biology and behaviour of Sesamia nonagrioides botanephaga Tams and Bowden (Lepidoptera: Noctuidae), a major stem borer pest of maize in Southern Ghana. West Afr. J. Applied Ecol., 8: 116-126.
Direct Link - Cardwell, K.F., F. Schulthess, R. Ndemah and Z. Ngoko, 1997. A systems approach to assess crop health and maize yield losses due to pests and diseases in Cameroon. Agric. Ecosyst. Environ., 65: 33-47.
CrossRefDirect Link - Chabi-Olaye, A., C. Nolte, F. Schulthess and C. Borgemeister, 2005. Abundance, dispersion and parasitism of the stem borer Busseola fusca (Lepidoptera: Noctuidae) in maize in the humid forest zone of Southern Cameroon. Bull. Entomol. Res., 95: 169-177.
CrossRefPubMedDirect Link - Chabi-Olaye, A., C. Nolte, F. Schulthess and C. Borgemeister, 2005. Relationships of intercropped maize, stem borer damage to maize yield and land-use efficiency in the humid forest of Cameroon. Bull. Entomol. Res., 95: 417-427.
CrossRefDirect Link - Crews, T.E. and M.B. Peoples, 2004. Legume versus fertilizer sources of nitrogen: Ecological tradeoffs and human needs. Agric. Ecosyst. Environ., 102: 279-297.
CrossRefDirect Link - Dhuyo, A.R. and N. M. Soomro, 2007. Efficacy of plant extracts against yellow rice stem borer, Scirpophaga incertulas (Walk.) in rice at Dokri, Sindh, Pakistan. Pak. Entomol., 29: 23-26.
Direct Link - Duale, A.H. and K.F. Nwanze, 1999. Incidence and distribution in sorghum of the spotted stem borer Chilo partellus and associated natural enemies in farmers` fields in Andhra Pradesh and Maharashtra states. Int. J. Pest Manage., 45: 3-7.
CrossRefDirect Link - Khan, Z.R, J.A. Pickett, J. Van Den Berg, L.J. Wadhams and C.M. Woodcock, 2000. Exploiting chemical ecology and species diversity: Stem borer and striga control for maize and sorghum in Africa. Pest Manage. Sci., 56: 957-962.
CrossRefDirect Link - Kruess, A. and T. Tscharntke, 2000. Species richness and parasitism in a fragmented landscape: experiments and field studies with insects on Vicia sepium. Oecologia, 122: 129-137.
CrossRefDirect Link - Linnet, S.G. and J.N. Adele, 2001. The chemical ecology of host location by parasitoids of African stemborers. Insect Sci. Appl., 21: 361-368.
Direct Link - Tahir, M., M.A. Malik, A. Tanveer and R. Ahmad, 2003. Agro-economic advantages of different canola-based intercropping systems. J. Agron., 2: 40-43.
CrossRefDirect Link - Khan, M., A.R. Saljoqi, A. Latif and K. Abdullah, 2003. Evaluation of some rice varieties against rice stem borer (Tryporyza incertulas). Asian J. Plant Sci., 2: 498-500.
CrossRefDirect Link - Baloch, M.S., I.U. Awan, G. Hassan, H.U. Khan and K. Abdullah, 2004. Rice stem borers moth (Tryporyza spp. Lepidoptera: Pyralidae) resting behaviour in relation to time of nursery seeding and seedlings per hill. Asian J. Plant Sci., 3: 45-49.
CrossRefDirect Link - Mulungu, L.S., M.R. Jilala, M.W. Mwatawala and J.K. Mwalilino, 2011. Assessment of damage due to larger grain borer (Prostephanus truncatus Horn) on stored paddy rice (Oryza sativa L. Poaceae). J. Entomol., 8: 295-300.
CrossRefDirect Link - Oparaeke, A.M. and G.C. Kuhiep, 2006. Toxicity of powders from indigenous plants against Sitophilus zeamais motsch on stored maize grains. J. Entomol., 3: 216-221.
CrossRefDirect Link - Otway, S.J., A. Hector and J.H. Lawton, 2005. Resource dilution effects on specialist insect herbivores in a grassland biodiversity experiment. J. Anim. Ecol., 74: 234-240.
CrossRefDirect Link - Pappas, M.L., G.D. Broufas and D.S. Koveos, 2011. Chrysopid predators and their role in biological control. J. Entomol., 8: 301-326.
CrossRefDirect Link - Pascal, M., 1998. Crop losses caused by maize stem borers (Lepidoptera: Noctuidae, Pyralidae) in Cote d`Ivoire, Africa: Statistical model based on damage assessment during the production cycle. J. Econ. Entomol., 91: 512-516.
Direct Link - Rami, K., W.A. Overholt, Z.R. Khan and A. Polaszek, 2002. Biology and management of economically important lepidopteran cereal stem borers in Africa. Annu. Rev. Entomol., 47: 701-731.
CrossRefDirect Link - Schulthess, F., A. Chabi-Olaye and S. Gounou, 2004. Multi-tropic level interactions in a cassava-maize mixed cropping system in the humid tropics of West Africa. Bull. Entomol. Res., 94: 261-272.
PubMedDirect Link - Setamou, M., F. Schulthess, H.M. Poehling and C. Borgemeister, 2000. Monitoring and modeling of field infestation and damage by the maize ear borer Mussidia nigrivenella (Lepidoptera: Pyralidae) in Benin, West Africa. J. Econ. Entomol., 93: 650-657.
Direct Link - Shelton, A.M. and -F.R. Badenes-Perez, 2006. Concepts and applications of trap cropping in pest management. Ann. Rev. Entomol., 51: 285-308.
CrossRefDirect Link - Songa, J.M., N. Jiang, F. Schulthess and C. Omwega, 2007. The role of intercropping different cereal species in controlling lepidopteran stem borers on maize in Kenya. J. Applied Entomol., 131: 40-49.
CrossRefDirect Link - Thies, C. and T. Tscharntke, 1999. Landscape structure and biological control in agroecosystems. Science, 285: 893-895.
CrossRefDirect Link - Tsubo, M., E. Mukhala, H.O. Ogindo and S. Walker, 2003. Productivity of maize-bean intercropping in a semiarid region of South Africa. Water SA, 29: 381-388.
Direct Link