
N. Steenken
Institute of Plant Diseases and Plant Protection, Leibniz Universit�t Hannover, Herrenh�user Str. 2, 30419 Hanover, Germany
N. Halaweh
Institute of Plant Diseases and Plant Protection, Leibniz Universit�t Hannover, Herrenh�user Str. 2, 30419 Hanover, Germany
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 2 | Page No.: 198-203







ABSTRACT
Host plant preference of Ceratothripoides claratris and Capsicum chlorosis virus (CaCV) were conducted in Bangkok, Thailand. More information about the host plants of CaCV and its thrips vector is needed, which could help working out cultural control measures that might hinder spread of the virus and pest population growth. The potential relevance of the results for crop rotation approaches that would reduce tospovirus epidemics is discussed and common pea is suggested to be integrated in crop rotation. Fourteen crop plants were potted and placed in a greenhouse, which was covered with non-thrips proof net to allow natural infestation. Thereafter, the number of adult thrips and larvae was counted in order to determine host preference. The susceptibility of plants to CaCV was tested by ELISA. Ceratothripoides claratris was the predominant thrips species that infested all offered plant species except for angled luffa and common pea. As scarce other thrips species were recorded, the high numbers of larvae were assumed to be Ceratothripoides claratris larval stages. Ceratothripoides claratris had a clear and significant preference to hosts in the families Solanaceae and Cucurbitaceae. Tospovirus could be detected in tomato, pepper and tobacco, which all belong to the family of Solanaceae.
PDF Abstract XML References Citation
Received: February 18, 2010;
Accepted: May 09, 2010;
Published: July 22, 2010
How to cite this article
N. Steenken and N. Halaweh, 2011. Host Plant Preference Study for Ceratothripoides claratris (Shumsher) (Thysanoptera: Thripidae) and CaCV (Genus Tospovirus; Family Bunyaviridae) in Bangkok, Thailand. Journal of Entomology, 8: 198-203.
DOI: 10.3923/je.2011.198.203
URL: https://scialert.net/abstract/?doi=je.2011.198.203
DOI: 10.3923/je.2011.198.203
URL: https://scialert.net/abstract/?doi=je.2011.198.203
INTRODUCTION
Herbivorous thrips are of economic interest, as some species damage their host plants by piercing and sucking in parenchyma cell contents, which leads to substantial yield losses (Premachandra et al., 2005a). Vegetable, legume and ornamental plants are preferred host groups (Pappu et al., 2009). In Bangkok, Thailand, Ceratothripoides claratris Shumsher (Thysanoptera: Thripidae) is the predominant pest species on tomatoes Lycopersicon sp. (Solanaceae) both in fields and in greenhouses (Murai et al., 2000; Rodmui, 2002; Premachandra et al., 2004). Premachandra et al. (2004) showed that C. claratris is well adapted to high temperatures (i.e., 30-35°C) within greenhouses in the tropics.
Besides the direct damage caused by feeding, a few thrips species transmit tospoviruses (Family Bunyaviridae) that cause serious epidemics. With estimated global yield losses of up to US $ 1 billion in a wide range of crops, tospoviruses are considered as the most aggressive emerging plant viruses (Naidu, 2007). The thrips/tospovirus relationship is very specific as only a few of the many known thrips species are able to acquire and transmit the virus (Adkins et al., 2005). Ten thrips species are accepted and confirmed vectors for tospovirus under experimental and natural conditions (Jones, 2005; Pappu et al., 2009). So far, there are 19 distinct tospovirus species (Pappu et al., 2009). Premachandra et al. (2005a) demonstrated that C. claratris is a vector of Capsicum chlorosis virus (isolate AIT) (CaCV-AIT) on tomato. For the survival of the pathogen it is essential that the vector lays its eggs on the CaCV-infected plants. The immobile first instar larvae acquire the virus and start to transmit as adults. Second instar larvae can transmit but are not that infectious. Thus, only some of the young thrips larvae are able to transmit the tospovirus and mainly as adults. The ability of thrips to acquire the tospovirus is dependent on the thrips developmental stages (Moritz et al., 2004) as well as on the thrips genetics (Halaweh and Poehling, 2009).
The thrips C. claratris invaded the greenhouses shortly after transplanting the crop and after lengthy periods of empty greenhouses. That is to say, other crops in the vicinity of the greenhouses could act as an alternative host and hence a reservoir for the thrips/tospovirus complex. Premachandra and Borgemeister (2006) determined the infestation of selected plant species in Thailand and defined tomatoes as the most preferable species for C. claratris. Other infested food crops were eggplant, water melon, cucumber and pumpkin. Thus far, however, there is no investigation about preferred host plant species for both C. claratris and CaCV.
Thrips are difficult to control due to their short life cycle and a narrow window of time for control possibilities (eggs in leaf tissue, pupation in the soil and resistance against insecticides) (Lewis et al., 1995; Naidu et al., 2006). There are no curative methods for virus infected plants. Preventive control methods therefore are predominantly designed to reduce population growth and spread of the virus. The objective of this investigation is to learn more about the host plants of the vector and the virus and thereby find preventive cultural methods, e.g., crop rotation and intercropping.
This study was part of a larger research project aiming to develop sustainable vegetable production under protected cultivation in the humid tropics.
MATERIALS AND METHODS
Host plants: The experiment was performed at the Asian Institute of Technology (AIT) in greater area of Bangkok (Pathumthani), Thailand, during January February 2006, as part of a collaborative project (Protected cultivation-an approach to sustainable vegetable production in the humid tropics) with the Faculty of Horticulture, Leibniz University of Hannover, Germany. Host plant seeds were sown at different times depending on their germination time in peat compost. To prevent the immigration of pests the plants were kept in a sealed greenhouse. The greenhouse was equipped with an evaporative fan and pad cooling system and was maintained at 28-30°C with a Relative Humidity (RH) of 90-100%. When seedlings were about 20 cm high, thereafter, they were planted coevally in 7 L containers and transferred into an experimental greenhouse where they were placed on one meter high tables. During the experiment a local soil/compost substrate (supplier: Dinwondeekankasat, Ayutthaya, Thailand, soil texture: 30% sand, 39% silt and 31% clay, organic matter: 28%, pH 5.3) was used. The experimental greenhouse was 2x4x0 m and was covered with a non thrips-tight 40-mesh screen (Econet M, Ludvig Svensson Inc., Kinna, Sweden) which allowed natural infestation of thrips from outside. Plants were daily irrigated in the morning with tap water at temperatures of 28-30°C and RH of 70-80%.
Counting of thrips: Specimens of C. claratris discovered in the experiments were identified by R. zur Strassen and voucher specimens were deposited at the Senckenberg Museum, Frankfurt, Germany (Premachandra et al., 2005b).
To determine levels of thrips infestation and hence host preference, number of adults and larvae were counted 5 weeks after transplanting in February 2006 (dry season). Whole plants were carefully collected in plastic bags and sealed before transferring them to the laboratory, where the leaves and other plant parts were placed into PVC containers (15x9 cm) containing 70% ethanol, this easily dislodged the insects. After five minutes, the plant materials were discarded, while the liquid was poured through a filter with thrips proof nylon gauze (64-microns open mesh size). The gauze was dried in the air and then examined under a stereomicroscope Olympus SZ30 (Olympus, Japan). Adults of C. claratris, Thrips palmi Karny (Thysanoptera: Thripidae) and other thrips species were counted. As there is no clear taxonomic key for the classification of larvae their absolute number was recorded. The thrips were determined by morphological characteristics following the key in Jangvitaya (1993).
Virus detection by enzyme-linked immunosorbent assay: The susceptibility of the plants to tospovirus (genus Tospovirus; family Bunyaviridae) was tested by DAS-ELISA (Double Antibody Sandwich-Enzyme-linked immunosorbent assay). Earlier experiments showed that it takes about 10-14 days until the virus can be detected in the whole plant (tomato) using DAS-ELISA (Premachandra et al., 2005b). Plants showing symptoms were tested with DAS-ELISA after five weeks, plants without symptoms were tested after nine weeks.
The compound direct ELISA for Watermelon silver mottle virus (WSMoV) and Groundnut bud necrosis virus (GBNV) (AGDIA® Inc., Elkhart, IN, USA. Cat. No. SRA 61500) was used, following the manufacturer’s instructions, for detecting CaCV in the plants (Premachandra et al., 2005b). CaCV-Infected leaves of tomato, tobacco and pepper were used as a positive control reference. And, as negative controls, tomato, pepper and melon leaves were used in addition to the sample puffer. The absorbance was carried out at OD405 nm (A405) using an EL 312 ELISA-reader (BIO-Tek Instruments, Inc, Vermont, USA). The results were evaluated as Rek (1987). To ensure the virus (CaCV), samples of the plant leaflets, infected with CaCV, were tested with PCR using specific primers (Knierim et al., 2006) at the Institute of Plant Diseases and Plant Protection, Hannover University.
Data analysis: The experimental design included five replicates per plant species and within the greenhouse the pots were arranged in a randomized complete block design. Thrips counts were analyzed using Kruskal-Wallis test via the SAS option proc npar1way. If the test result was significant it was followed by Bonferroni (Dunn) test (p = 0.05).
RESULTS
Data from the counting of the thrips and the ELISA-test with antibodies to serogroup IV are given in Table 1.
From the total of 653 counted adults Thrips, only 11 T. palmi and 5 other unidentified thrips species were counted, which clearly demonstrates the preponderance of C. claratris adults over other available thrips species in the experimental area. Almost no T. palmi or other thrips species occurred on the offered plants in the greenhouse. In addition, C. claratris adults and larvae had a similar distribution; therefore, it is accepted that the larvae counts represent C. claratris larvae.Adults of C. claratris had significantly preferred certain hosts among others for feeding (H = 48.69, 13 d.f., p = 0.0001) as well as breeding, as determined from the larvae counts (H = 51.11, 13 d.f., p = 0.0001). Adults of C. claratris were collected from all tested crop plants except for angled luffa and were very low on pumpkin, common pea, lettuce and pepper Chili.
| Table 1: | Adult and larvae counts of the thrips Ceratothripoides claratris and Thrips palmi as well as CaCV incidence |
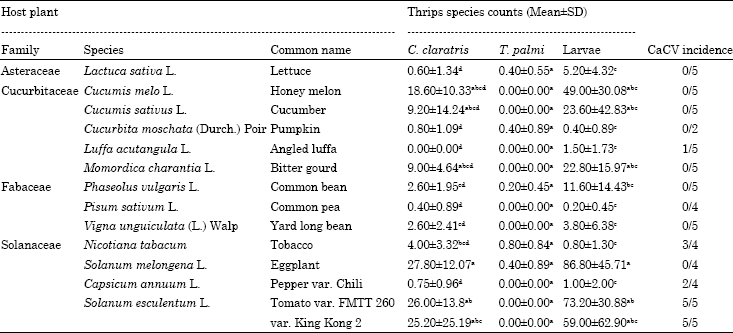 | |
| Thrips counts were analysed using Kruskal-Wallis test, followed by Bonferroni (Dunn) test (p = 0.05). Means followed by different letters in columns indicate significant difference | |
The highest infestation levels of C. claratris were on the eggplants and the two tomato varieties. For the larvae, the preferred hosts were similar to those of the adults. Moderate, though not significant, infestation of larvae and adults was found on honey melon, cucumber and bitter gourd.
The presence of CaCV isolate was confirmed by the PCR tests except in the angled luffa specimen (data not shown). CaCV was detected by DAS-ELISA and PCR in three repetitions of tobacco (3/4), two of pepper Chili (2/4) and all ten tomato samples (Table 1). The infested plant species all belong to the family of Solanaceae. Exceptionally one repetition of angled luffa (Cucurbitaceae) had a positive ELISA-result. Hence, susceptibility for tospovirus could not be excluded for this plant species. Noteworthy, some crops, though, were a preferred host of C. claratris, it was not a good host of the tospovirus.
DISCUSSION
The results clearly show that C. claratris is the predominant thrips species in the greenhouses at the AIT as described by Premachandra et al. in 2004. No more than one T. palmi on only a few vegetable species could be collected. Previous studies showed that T. palmi is not as well adapted to the high temperatures in the tropics and in the greenhouses as C. claratris (Murai, 2001; Premachandra et al., 2004).
Premachandra and Borgemeister (2006) already described tomatoes and eggplants as suitable plants for feeding and reproduction of C. claratris. From our results, moreover, honey melon, cucumber, common bean and bitter gourd were preferred host plants for C. claratris. Lettuce could not be excluded as a suitable host with more than five larvae in average. CaCV systemic infected lettuce has also been described (McMichael et al., 2002).
Despite the high thrips infestation, the host preference of C. claratris is actually selective. Almost no thrips specimens were found on pumpkin, angled luffa and common pea. In contrast, Premachandra and Borgemeister (2006) described pumpkin as a good host plant for C. claratris.
Common pea had the lowest infestation level with only two adults and one larva on all the five replicates. This investigation, to our knowledge, is the only host plant preference study for C. claratris on common pea. Therefore, the common pea could be excluded as a host plant for C. claratris and thus may be considered for crop rotation control approaches.
Although, pepper had a very low infestation rate, CaCV could be detected in half of the plants. Pepper has already been described as a host of CaCV (McMichael et al, 2002; Premachandra et al., 2004; Persley et al., 2005). Despite the low infestation by C. claratris, the plants were actually infected with CaCV, which shows that vectors can lead to virus epidemics regardless of the size of their population (Kucharek et al., 2000).
All of the tested tomato plants were infected with CaCV. This confirms tomato as the optimal host for both CaCV and its vector C. claratris and corroborate results of (Premachandra et al., 2005b). Although the infestation of tobacco by C. claratris was low, the tospovirus infection level was high. Nicotiana sp. was described as an important host mainly to the tospoviruses (Premachandra et al. 2004).
The other plant species were not susceptible to tospovirus. Crops that are resistant to CaCV could be a dead end for the tospovirus (Pappu et al., 2009). In other words, as CaCV has a smaller host range and seems to prefer plants in the family Solanaceae, such important information may be useful when crop rotations are considered in protected cultivation in order to reduce tospovirus epidemics.
CONCLUSION
Although, C. claratris has colonised the majority of the offered host crops, it selectively and significantly prefers only a few of them. In contrast to its vector, CaCV has less host range, which was mostly within the Solanaceae family. In protected cultivation crop rotation could reduce thrips population and the spread of the tospovirus. Therefore a rotation with tomato and common pea could be a preventive control method against thrips population growth and the consequent tospovirus spread. Moreover, Crops that are resistant to CaCV, such as cucumber, pumpkin, common bean, common pea or eggplant, could reduce the spread of the tospovirus when included in crop rotations with tomato culture.
REFERENCES
- Halaweh, N. and H.M. Poehling, 2009. Inheritance of vector competence by the thrips Ceratothripoides claratris (Shumsher) (Thysanoptera: Thripidae). J. Applied Entomol., 133: 386-393.
CrossRefDirect Link - Jones, D.R., 2005. Plant viruses transmitted by thrips. Eur. J. Plant Pathol., 113: 119-157.
CrossRefDirect Link - Knierim, D., R. Blawid and E. Maiss, 2006. The complete nucleotide sequence of a capsicum chlorosis virus isolate from Lycopersicum esculentum in Thailand. Arch. Virol., 151: 1761-1782.
CrossRefDirect Link - McMichael, L.A., D.M. Persley and J.E. Thomas, 2002. A new tospovirus serogroup IV species infecting capsicum and tomato in Queensland, Australia. Aust. Plant Pathol. Soc., 31: 231-239.
CrossRefDirect Link - Moritz, G., S. Kumm and L. Mound, 2004. Tospovirus transmission depends on thrips ontogeny. Virus Res., 100: 143-149.
CrossRef - Murai, T., S. Kawai, W. Chongratanameteekul and F. Nakasuji, 2000. Damage to tomato by Ceratothripoides claratris (Shumsher) (Thysanoptera: Thripidae) in central Thailand and a note on its parasitoid, Goethena shakespearei Girault (Hymenoptera: Eulophidae). Applied Entomol. Zool., 35: 505-507.
Direct Link - Murai, T., 2001. The pest and vector from the East: Thrips palmi. Thrips and Tospoviruses: Proceedings of the 7th International Symposium on Thysanoptera, (IST`01), Research Institute for Bioresources, Okayama University, Kurashiki, pp: 19-32.
Direct Link - Naidu, A., D. Riley, K. Ravi, C. Chaisuekul and S. Adkins, 2006. Integrated management of thrips-borne tospoviruses in vegetable cropping systems in South and Southeast Asia. February 1, Department of Plant Pathology, Washington State University. http://www.ars.usda.gov/research/publications/publications.htm?SEQ_NO_115=192078.
- Naidu, A., 2007. An integrated pest management approach for mitigating the impact of thrips-borne tospoviruses in vegetable cropping systems. Proceedings of the Central Asian Region Integrated Pest Management Stakeholders Forum. Irrigated Research and Extension Center, Washington State University, pp: 72-78.
- Pappu, H.R., R.A.C. Jones and A.K. Jain, 2009. Global status of tospovirus epidemics in diverse cropping systems: Successes achieved and challenges ahead. Virus Res., 141: 219-236.
Direct Link - Persley, D.M., J.E. Thomas and M. Sharman, 2005. Tospoviruses-An Australian perspective. Aust. Plant Pathol., 35: 161-180.
CrossRefDirect Link - Premachandra, W.T., C. Borgemeister, A. Chabi-Olaye and H.M. Poehling, 2004. Influence of temperature on the development, reproduction and longevity of Ceratothripoides claratris (Thysanoptera: Thripidae) on tomatoes. Bull. Entomol. Res., 94: 377-384.
PubMed - Premachandra, W.T., C. Borgemeister and H.M. Poehling, 2005. Effects of neem and spinosad on Ceratothripoides claratris (Thysanoptera: Thripidae), an important vegetable pest in Thailand, under laboratory and greenhouse conditions. J. Econ. Entomol., 98: 438-448.
CrossRef - Premachandra, W.T., C. Borgemeister, E. Maiss, D. Knierim and H.M. Poehling, 2005. Ceratothripoides claratris, a New Vector of a Capsicum chlorosis virus isolate infecting tomato in Thailand. Phytopathology, 95: 659-663.
PubMed - Premachandra, W.T.S.D. and C. Borgemeister, 2006. Infestation of Ceratothripoides claratris (Shumsher) (Thysanoptera: Thripidae) on selected food crops in Thailand. Ruhuna J. Sci., 1: 41-46.
Direct Link