
N. Geetha
Division of Crop Protection,Coimbatore-641007, Tamil Nadu, India
R. Balakrishnan
Section of Statistics, Sugarcane Breeding Institute, Coimbatore-641007, Tamil Nadu, India
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 2 | Page No.: 164-173







ABSTRACT
Laboratory-reared Trichogramma chilonis Ishii was assessed for its ability to move in open field through parasitism on sentinel cards of Corcyra cephalonica placed at distances of 1, 3, 5, 7, 10, 15, 20, 25 and 30 m, from the point of release of parasitoids at eight cardinal directions. Sentinel cards were recovered from the field after 1-7 days time at 24 h interval to observe parasitism and adult emergence. The dispersal rate was affected significantly by the time allowed as well as by the distances from the release point while the impact of direction was not significant. The overall dispersal rate was found to increase from 23.56% at 24 h to 37.07% on the completion of third day, irrespective of the distance or direction at which the cards were kept. The mean dispersal rate over the seven time periods tested was quite high at 1-10 m distance (73.78 to 32.05) which decreased to 7.67 at 30 m. Adult emergence did not show a definite influence by the interactive elements. High rates of adult emergence was observed (77.72-92.44%) in the cards that were left in the field for 1-7 days. Overlapping but significant variations in the emergence rates (76.06-91.9%) were observed in the cards kept at different distances from the release point. Results indicated that three days of time stabilized parasitoid dispersal and that at least 13 release points are required per acre of release area for ensuring uniform dispersal.
PDF Abstract XML References Citation
Received: April 05, 2010;
Accepted: May 24, 2010;
Published: July 22, 2010
How to cite this article
N. Geetha and R. Balakrishnan, 2011. Temporal and Spatial Dispersal of Laboratory-Reared Trichogramma chilonis Ishii in Open Field. Journal of Entomology, 8: 164-173.
DOI: 10.3923/je.2011.164.173
URL: https://scialert.net/abstract/?doi=je.2011.164.173
DOI: 10.3923/je.2011.164.173
URL: https://scialert.net/abstract/?doi=je.2011.164.173
INTRODUCTION
The egg parasitoid Trichogramma chilonis Ishii is used throughout India for management of many lepidopterous pests across the crops through inundative releases. Field fitness of such parasitoids mass reared for biological control is generally evaluated by assessing parasitization rates of pest eggs in the target crop (Kolliker-Ott et al., 2004) and efficient dispersal is a key factor in the host searching leading to high parasitization. Dispersal capacity and host location ability in the field are important quality attributes in biological control, but are difficult to estimate in the laboratory (Kölliker-Ott et al., 2004). In field, being small, Trichogrammatids are believed to disperse to short distances and the rates of such dispersal of the Trichogramma species is reduced in direct relation to the distance from the point of release (Geetha and Balakrishnan, 2010; Wright et al., 2001). The factors that influence the dispersal ability of a parasitoid may either be of parasitoid origin: physiological state of female and wing size (Reznik and Klyueva, 2006; Kolliker-Ott et al., 2004) or of host: egg density (Sarkar et al., 1999) or of host plant: crop age or growth (Geetha and Balakrishnan, 2010; Muhammad et al., 2004) and plant architecture (Gingras et al., 2003, 2008; McDougall and Mills, 1997; Wang et al., 1997), or climatic: wind (Yu et al., 1984), rain (Chapman, 2007) and temperature (Singh and Shenhmar, 2008; Sohi et al., 1996; Suverkropp et al., 2001) or strategical: density of parasitoid (Ayvaz et al., 2005; Pfannenstiel et al., 1996; Singh and Jalali, 1992; Taylor, 1978) or number of release points (Wang and Shipp, 2004) or the size of the experimental area (Canto-Silva et al., 2006). Mostly, these studies have been done to assess the ability of the Trichogramma sp. to move every 24 h but at least the T. chilonis adults live up to 11.6 days (Nadeem et al., 2009) ovipositing up to six days (Baitha et al., 2005) and survive in the field for many days though it is reported that the parasitoid lays most of its eggs on the first two days after emergence in the laboratory (Baitha and Verma, 2003) when the host is readily available.
Hence, in the present study, the ability of the parasitoid to move from the point of release as influenced by time allowed since release from 1-7 days is assessed in the open field and the use of such results on deciding the number of release points during inundative releases is discussed.
MATERIALS AND METHODS
The present study was undertaken in the fields of Sugarcane Breeding Institute, Coimbatore (77°E longitude and 11°N latitude in Southern India) during 2008. The factitious host Corcyra cephalonica Stainton eggs were used as sentinel cards to observe the parasitism by the dispersing parasitoids instead of its native host, sugarcane inter node borer Chilo sacchariphagus indicus (Kapur) eggs since mass-culturing technique for the latter is unavailable. However, in the inundative release strategy, UV-sterilized C. cephalonica eggs are used as a factitious host for mass rearing T. chilonis.
The T. chilonis parasitoids were derived from the culture maintained in the laboratory of insectary (28±2°C, 52±4% RH) at Sugarcane Breeding Institute (Coimbatore, Tamil Nadu). The Trichogramma cards were used at the stage when the adults had emerged, were less than 24 h old and mated. The parasitoids were used at the rate of Ca. Eight thousend for each of 3 replicates (the dose per unit area derived -30 m radius circle of 2825 m2 area was corresponding to field dosages of 2 cc/ac suggested (Geetha et al., 2009) for management of inter node borer in sugarcane).
The sentinel cards were thick chart paper sheets of 8x1 cm size with at least 100 UV-inactivated and unparasitised eggs of C. cephalonica. The fresh sentinel cards were positioned at 1, 3, 5, 7, 10, 15, 20, 25 and 30 m, from the point of release of parasitoids at eight cardinal directions along the straight radius lines forming a ring around the release point at every distance studied as per the protocol used by Geetha and Balakrishnan (2010). The Tricho cards (with fresh mated T. chilonis adults) used for the study were held in glass vials till release and the cards were placed in the sugarcane field in a small circle with the central point of the circle serving as release site of the parasitoid. Both the parasitoid cards and sentinel cards were tied onto the bamboo pegs placed at point of release and at specified distances, respectively.
At a single point seven such sentinel cards were placed simultaneously and at the interval of 24 each card was collected for determination of incidence of parasitism. Thus one of the seven cards at each monitoring point was collected after 24 h followed by one on day 2 and so on. The study was conducted for a period of seven days during 12.8.08-18.8.2008. When the sentinel cards were collected the distance and direction from the release points were noted. Records of weather parameters were obtained from the Meteorological Laboratory located at Sugarcane Breeding Institute, Coimbatore.
The recovered cards were maintained at the ambient temperature of 28-30°C for 1-7 days depending on the day of recovery, to observe the percentage of parasitized eggs. When there was complete destruction of a sample by predators, the data was construed as missing data (and replaced with mean of surrounding replication). The natural population of T. chilonis at the trial site was assessed by placing sentinel cards only, a week prior to experimentation as well as during the experiments at 100 m distance.
In an earlier study done in 2004-2005, the effect of plant age and distance on the dispersal of the parasitoid was found be distinct (Geetha and Balakrishnan, 2010). So in the present study, to eliminate the effect of crop cues and architecture, the study was undertaken in a crop-free field. Thus the results would reveal the effect of wind direction, distance of from the point of release and the time allowed for the parasitoids to move sans crop architecture and chemical cues.
The percentage data were transformed using arc-sine square root transformation. Treatment means of percent parasitism and emergence were analyzed through a nested split-split plot analysis of variance and means were separated by least significant difference (LSD) values at p = 0.05 level.
RESULTS
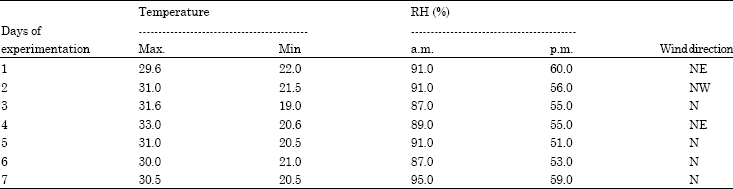
The weather parameters existed in the field during the experiments are given in Table 1. The temperature to some extent was higher than the optimal temperature required by T. chilonis for parasitism which is reported to be 28°C (Nadeem et al., 2009). The temperature ranged from 30-33°C during the day and 19-22°C during night. RH was 87-95% in the morning and 51 to 60% in the afternoon (Table 1). The natural parasitism rates in sentinel cards prior to and during the experiments were 0.0-0.05% and thus were not considered for analysis.
Dispersal as influenced by the time, distance, direction:
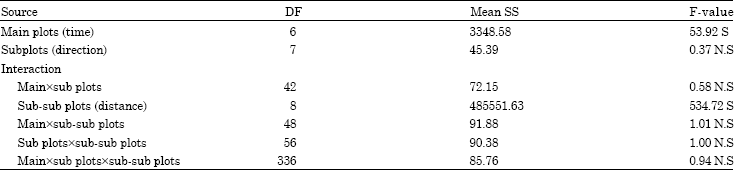
Parasitism: The dispersal rate was influenced significantly by the time allowed as well as by the distances from the release point while the impact of direction o n dispersal was insignificant (Table 2). The parasitoid was able to move up to farthest distance (30 m) tested in the present study (Table 3).
| Table 1: | Weather parameters during the conduct of experiment |
 | |
| Table 2: | ANOVA table for parasitism (using angular values) |
 | |
| Significance at p = 0.05 | |
| Table 3: | Percent parasitism: time x distance |
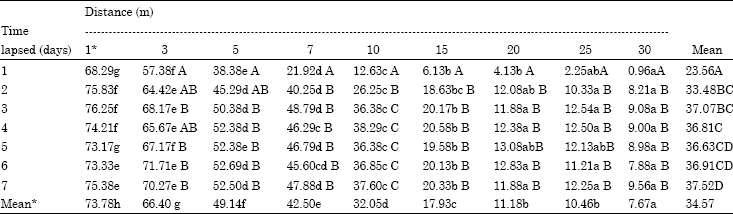 | |
| *No significant differences among mean values were found among the mean parasitism at different days at 1m distance. Mean values in a column (upper case) and in a row (lower case) followed by the same letter are not significantly different (LSD, p = 0.05). Analysis conducted on arcsine- transformed data; Interaction effect of Days (Time) x Distance was insignificant (p=0.05) | |
The overall dispersal rate was found to increase from 23.56% at 24 h to 37.07% on the completion of third day, irrespective of the distance or direction at which the cards were kept. After 72 h the dispersal rate was stabilized (Table 3). With in each time period tested, the impact of distance was categorically observed. For example, at 24 h after release the dispersal steadily and proportionally decreased from 68.29% at 1 m distance to 12.63% at 10 m distance. However when compared across the days, the impact of lapse of time was found to have a positive significant effect till 72 h after release at most of the distances tested, with the average percent parasitism across the distances increasing from 23.5 (on day 1) to 37.07 (on day 3). The latter was on par with the parasitism levels by parasitoid allowed for still longer periods (4-7 days) in the field.
Distance had a visible direct impact on the dispersal. The mean dispersal rate over all the time periods tested, was quite high at 1-10 m distance as indicated by the percent parasitism ranging from 73.78 to 32.05 which decreased to 7.67 at 30 m (Table 3). The effect of time allowed on the dispersal was not observed in the shortest distance tested (1 m) though for all other distances tested, with passage of time significantly more dispersal was found. For example, at distances of 20, 25, 30 m, the percent parasitism observed on day 3 was 3-10 fold more than what was observed within 24 h time (Table 3).
Dispersal at any distance from the point of release was not influenced by the direction of wind (Table 4). On the other hand, within each wind direction, the movement of the parasitoid was found to be affected by the distance from the release point (Table 5). The mean parasitism over all the time periods tested was not found to vary significantly from 33.27 (North) to 35.63 (North East) though, the wind flow during the experimental period was mostly North or North East. Within each direction of wind, the impact of time allowed (days) on dispersal ability was found with overlapping significancse (Table 4).
Interaction effects: Interaction effects of time allowed (days) x directions, time allowed x distances and direction x distance were found to be insignificant showing no effect on the dispersal rate of the T. chilonis (Table 2). Similarly there was no significant effect of three way interaction of time x direction x distance on dispersal ability (Table 2).
Emergence: The percent emergence of the parasitoid, T. chilonis was very high in the open field ranging from 77.72 to 92.44 with the highest on day 2 and the lowest on day 4.Though the climatic factors were almost the same on day 2 as in other days, the temperature on day 3 was the highest among the days of test.
| Table 4: | Percent parasitism: time x direction |
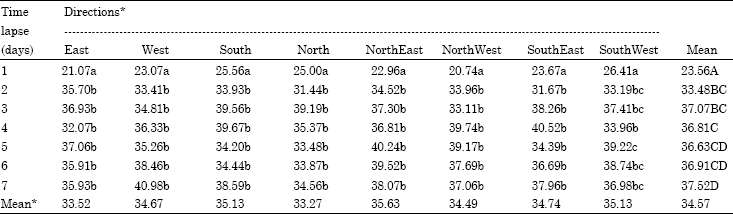 | |
| *Over all means of parasitism across the directions are not significantly different; overall means of parasitism across days (last column) followed by same letters are not significantly different; Means in a column followed by same letter (lower case) are not significantly different (LSD, p = 0.05); analysis conducted on arc sine transformed data | |
| Table 5: | Percent parasitism: direction x distance |
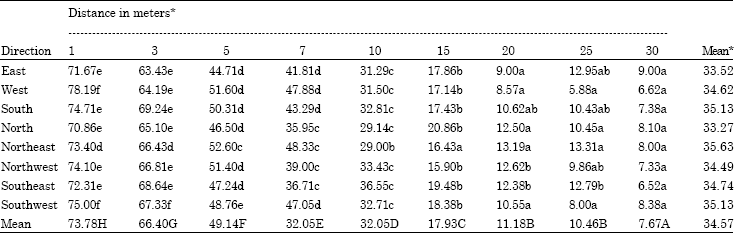 | |
| *Over all means of parasitism in different directions were not significantly different; Means of percent parasitism across the directions in a single distance, were not significantly different; Means in a row followed by same letters (lower case) and overall means at different distances as shown in the last row (upper case) are not significantly different (LSD, p = 0.05); analysis conducted on arc sine transformed values | |
| Table 6: | ANOVA table for emergence (using angular values) |
 | |
In all the directions at which the sentinel cards were kept, irrespective of the day and distance, the percent emergence was high (84.19 to 87.5%) with no significant differences (Table 6). With respect to distances, the emergence was more than 80% up to 20 m distances with overlapping levels of significance (Table 7).
| Table 7: | Percent emergence as influenced by individual effects of time, direction and distance |
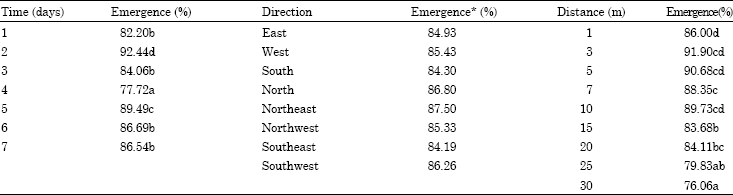 | |
| *Means are not significantly different; Means in a column followed by similar alphabets are not significantly different (LSD, p = 0.05); Interaction effects of Timex Direction, DirectionxDistances, DaysxDirectionxDistances were insignificant (p = 0.05) while Time (days)xdistance effect was significant | |
The lower rate of emergence in the distances beyond 20 m could probably due to the lower (smaller values) of % parasitism at farthest distances where in the impact of even few failures to emerge were seen magnified in calculations. Significance of time lapse since release, i.e., days for which the sentinel cards were left in the field on the emergence was also overlapping (Table 7) and not proportionate.
Interaction between either any of the two factors or among all the three elements tested did not have a significant effect on the emergence (Table 6).
DISCUSSION
The high rates of parasitization in the cards placed up to 3 m distance show concentrated pattern of dispersal closer to the point of release, which however declined progressively with increase in distance. At 10 m distance from release point the percent parasitism was only 12.63 in 24 h time, though by seventh day it increased to 37.6. This is in contrast with the earlier study wherein the dispersal in 45 days old sugarcane crop by T. chilonis was 28.96% in 24 h at 10 m distance from the release point (Geetha and Balakrishnan, 2010) indicating the role of plant cues and possibly the cooler microclimate in the field, in better dispersal. However, when more time was allowed to lapse after release, increased percent parasitism was observed at most of the distances tested which was more pronounced in the farthest distances from release point. This may be because after settling in a point on the day one, the parasitoid might have started dispersing again further on the consecutive day. Stabilization in parasitization after 72 h indicates either that the parasitoids had perished in the field by then, or they have off- loaded all their eggs within this time frame. Faster decline in parasitization due to time lapse has been observed earlier where in intensity of parasitization by T. chilonis was reduced at 48 h post release of parasitoids compared to 24 h time (Muhammad et al., 2004).
Similar trend of increased dispersal with time was observed in a two year study on Trichogramma ostriniae Pang and Chen from release point, the greatest number of wasps recaptured occurred from 3-4 days after emergence (Chapman, 2007). However, the majority of the newly-emerged adults of Trichogramma pretiosum Riley remained around the release point during days 1 and 2 post-release, resulting in the highest parasitoid-induced mortality of 38 and 45% on the release plant (Wang and Shipp, 2004).
Distance had influenced the rate of dispersal which is in line with the findings in earlier work by several authors in many Trichogramma sp. (Bigler et al., 1988; Geetha and Balakrishnan, 2010; Mills, 2003; Wang et al., 1996, 1997; Wright et al., 2001). Though for other species of Trichogramma high dispersal ability has been reported, T. chilonis dispersal is reported to be limited (Anonymous, 1995; Muhammad et al., 2004; Singh et al., 1994). Brar et al. (2000) observed that T. chilonis could disperse up to 9 m in search of its host and maximum parasitization was observed up to 2 m. Conversely, the study by Sohi et al. (1996) did not show any significant differences in the parasitization rates by T.chilonis between 2-10 m distances. Similar low levels of parasitization in sentinel cards by T. chilonis in proportion to the distance tested has been established by Singh and Shenhmar (2008) where the maximum parasitization was recorded at 1 m and the least at 10 m from the release point. Thomas and Rainer (2009) found parasitism by Trichogramma brassicae to be the highest in the close vicinity of the release point, i.e., 86%, whereas 6.5 m away the mean parasitization rate decreased to 3%though the parasitoids persisted in the field for 9 days. In open field conditions, Trichogramma sp., seem to have low ability to disperse on their own which may probably be because they lose their ability for directed flight and a similar phenomenon was observed in case of T. pretiosum (Smith, 1996; Wang and Shipp, 2004).
These declines in parasitism are likely to reflect wasp density, as in these experiments wasps are released at a single point in the field and there is a decrease in parasitoid density with increasing distance from the release point (Kolliker-Ott et al., 2004). Further, Trichogramma females concentrate their search for hosts close to the release point (Gingras et al., 2003). Due to the small size of Trichogramma the exact physiological or environmental reasons behind the characteristic dispersal ability are difficult to segregate.
On the contrary, Parsons and Ullyett (1936) concluded that dispersal of Trichogramma sp. was rapid, extending from the centre to the outer part of 5-acre fields within 24 h. Similar quick movement of T. ostriniae adults to 45 m from the release point was observed with high rates of parasitization (20-50%) at this distance, but the levels decreased with increasing distance from the release point (Chapman et al., 2009).
There are several reasons for the findings on the level of parasitism observed in the present study. Dispersal leading to parasitism was dependent on release rate with the proportion of parasitized egg masses and eggs per mass increasing with the number of Trichogramma platneri Nagarkatti released (Pfannenstiel et al., 1996). However in the present study, absence of crop cues could have been an hindrance in directed host location and thus increase in the parasitoid density (8000 parasitoids) at the release point as compared to the release rate (4000 parasitoids) and the increase in the host density (seven times more) from those used in the earlier study (Geetha and Balakrishnan, 2010) has not really altered the resultant parasitism. Combined with host plant attraction, it could have been still higher.
It was thought the parasitoid would disperse downwind due to its small size. But in the present study no association of the dispersal was found with wind direction was observed. This kind of independence from wind is reported in the study of Chapman (2007) where in the effect of drift was negligible in the dispersal of the T. ostriniae. However, Saavedra et al. (1997) had observed that the low wind speed on the release days allowed uniform T. pretiosum dispersal in experimental area while Singh et al. (1994) found higher parasitism by T. chilonis in the direction of wind. In another study with the latter, dispersal of the parasitoid was found to be influenced by both distance and wind direction (Anonymous, 1995).
There were non-significant differences in parasitoid emergence pattern with respect to directions in any distance or on any day while unexpectedly the time allowed as well as distances significantly affected though the emergence rates were overlapping. Among the days, the emergence on day four was the lowest, corresponding to the highest temperature (33°C) recorded in the open field during experimentation. In a cropped- field though, the microclimate would have been optimal at this temperature. Comparable results on the emergence of parasitoids have been reported by Geetha and Balakrishnan (2010). Wind speed which was not recorded or the number of hours spent by the parasitized sentinel cards in the experimental tropical climate could have had an impact on the adult emergence rate by causing loss of moisture in the open field. Environmental conditions, primarily temperature has been shown to affect the movement activity of Trichogramma sp. (Fournier and Bovin, 2000; Suverkropp et al., 2001). Temperature has a profound influence on the survival and dispersal of T. chilonis in the field and seasonal variation in parasitism rates due to change in temperature has been reported by many workers (Brar et al., 2000; Muhammad et al., 2004; Nadeem et al., 2009; Singh and Shenhmar, 2008; Sohi et al., 1996).
Importance of this study in the light of inundative releases Insect dispersal has been and will always be a study under the realm of ecologists. However, this knowledge of insect dispersal may be of greater importance in the insect management tactics (Stinner et al., 1983) and for a biocontrol advocate, it becomes a prerogative as well as a prerequisite while formulating an inundative release program. As narrated in the text there are several factors affecting the movement of this parasitoid in the field. Understanding the dispersal capacity and dynamics is important in terms of developing effective release techniques and for predicting the efficacy of releases (Wright et al., 2001). While inundative releases rely on rapid dispersal and uniform coverage of the target area (Chapman, 2007), limited dispersal ability may be an impediment for parasitization efficacy but fine-tuning of release rates and number of release points might adequately compensate that. Mills (2003) had suggested that either more frequent releases, or an extended period of emergence of adult parasitoids from each release, could increase the efficacy of Trichogramma inundation to compensate the lower dispersal. Based on the findings from the current study, we understand that the released parasitoid move only up to 10 m effectively and restrict their parasitization up to 3 days in the field though the presence of host plant cues and complexity of architecture could have either added or diminished these results further. Because host location by Trichogramma sp., is influenced by structural complexity but with time they seem to be nullified. So with the present results, it can safely be assumed that the number of the release points is to be at least 13 per ac for uniform high distribution resulting in good dispersal and thus host parasitization. Release of parasitoid cards containing differential emergence of adults would be another efficient alternative for ensuring presence of parasitoid in a temporal continuum for efficient pest management.
ACKNOWLEDGMENTS
Authors thank Dr. N. Vijayan Nair, the Director, Sugarcane Breeding Institute for the facilities and encouragement. Dr. P. Rakkiyappan, the Head (Crop Production) and Shri. R. Palanisamy, SBI are gratefully acknowledged for the provision of weather data.
REFERENCES
- Bigler, F., M. Bieri, A. Fritschy and K. Seidel, 1988. Variation in locomotion between laboratory strains of Trichogramma maidis and its impact on parasitism of eggs of Ostrinia nubilalis in the field. Entomol. Exp. Appl., 49: 283-290.
CrossRef - Chapman, A.V., T.P. Kuhar, P.B. Schultz and C.C. Brewster, 2009. Dispersal of Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) in potato fields. Environ. Entomol., 38: 677-685.
Direct Link - Canto-Silva, C.R., R. Kolberg, H.P. Romanowski and L. Redaelli, 2006. Dispersal of the egg parasitoid Gryon gallardoi (Brethes) (Hymenoptera: Scelionidae) in tobacco crops. Braz. J. Biol., 66: 9-17.
Direct Link - Fournier, F. and G. Boivin, 2000. Comparative dispersal of Trichogramma evanescens and Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) in relation to environmental conditions. Environ. Entomol., 29: 55-63.
CrossRef - Geetha, N. and R. Balakrishnan, 2010. Dispersal pattern of Trichogramma chilonis Ishii in sugarcane field. J. Biol. Control, 24: 1-7.
Direct Link - Gingras, D., P. Dutilleul and G. Boivin, 2003. Effect of plant structure on host finding capacity of lepidopterous pests of crucifers by two Trichogramma parasitoids. Biol. Control, 27: 25-31.
CrossRef - Gingras, D., P. Dutilleul and G. Boivin, 2008. Effect of plant structure on searching strategy and searching efficiency of Trichogramma turkestanica. J. Insect Sci., 8: 1-9.
CrossRefDirect Link - Kolliker-Ott, U.M., F. Bigler and A.A. Hoffmann, 2004. Field dispersal and host location of Trichogramma brassicae is influenced by wing size but not wing shape. Biol. Control, 31: 1-10.
CrossRef - Mills, N.J., 2003. Augmentation in orchards: Improving the efficacy of Trichogramma Inundation. Proceeding of the 1st International Symposium on Biological Control of Arthropods, Jan. 14-18, 2002, University of California, Berkeley, California, U.S.A., pp: 130-135.
Direct Link - Muhammad, H., N. Sajid and M.T. Siddiquee, 2004. Dispersing and parasitizing ability in Trichogramma chilonis Ishii in early and late sown NIAB-86 cotton variety. Pak. J. Sci. Indus. Res., 47: 309-311.
Direct Link - Parsons, F.S. and G.C. Ullyett, 1936. Investigations on Trichogramma lutea Gir., as a parasite of the cotton boll worm Heliothis obsoleta Fabr. Bull. Entomol. Res., 27: 219-235.
CrossRef - Pfannenstiel, R.S., J.F. Brunner and M.D. Doerr, 1996. Trichogramma platneri and leaf rollers. Proceedings of the 70th Annual Western Orchard Pest and Disease Management Conference, Jan. 10-12, Portland, pp: 1-3.
Direct Link - Reznik, S.Y. and N.Z. Klyueva, 2006. Egg retention and dispersal activity in the parasitoid wasp, Trichogramma principium. J. Insect Sci., 6: 1-6.
PubMedDirect Link - Saavedra, J.L.D., J.B. Torres and M.G. Ruiz, 1997. Dispersal and parasitism of Heliothis virescens eggs by Trichogramma pretiosum (Riley) in cotton. Int. J. Pest Manage., 43: 169-171.
CrossRef - Singh, S. and M. Shenhmar, 2008. Host searching ability of genetically improved high temperature tolerant strain of Trichogramma chilonis ishii in sugarcane. Ann. Plant Prot. Sci., 16: 107-110.
Direct Link - Smith, S.M., 1996. Biological control with Trichogramma: Advances, successes and potential of their use. Ann. Rev. Entomol., 41: 375-406.
CrossRef - Stinner, R.E., C.S. Barfield, J.L. Stimac and L. Dohse, 1983. Dispersal and movement of insect pests. Ann. Rev. Entomol., 28: 319-335.
CrossRef - Suverkropp, B.P., F. Bigler and J.C. van Lenteren, 2001. Temperature influences walking speed and walking activity of Trichogramma brassicae (Hymenoptera: Trichogrammatidae). J. Applied Entomol., 125: 303-307.
CrossRef - Taylor, R.A.J., 1978. The relationship between density and distance of dispersing insects. Ecol. Entomol., 3: 63-70.
CrossRef - Thomas, K. and M. Rainer, 2009. Performance of two Trichogramma brassicae strains under greenhouse and field conditions for biocontrol of the silver Y moth in spinach cultures. J. Pest Sci., 82: 73-79.
CrossRef - Wang, B., D.N. Ferro and D.W. Hosmer, 1997. Importance of plant size, distribution of egg masses and weather conditions on egg parasitism of the European corn borer, Ostrinia nubilalis, by Trichogramma ostriniae in sweet corn. Entomol. Exp. Appl., 83: 337-345.
CrossRef - Wright, M.G., M.P. Hoffmann, S.A. Chenus and J. Gardner, 2001. Dispersal behavior of Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) in sweet corn fields: Implications for augmentative releases against Ostrinia nubilalis (Lepidoptera: Crambidae). Biol. Control, 22: 29-37.
CrossRef - Yu, D.S.K., J.E. Laing and E.A.C. Hagley, 1984. Dispersal of Trichogramma spp. (Hymenoptera: Trichogrammatidae) in an apple orchard after inundative releases. Environ. Entomol., 13: 371-374.
Direct Link