
R. Manohar Reddy
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
Suresh Rai
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
A.K. Srivastava
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
P.K. Kar
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
M.K. Sinha
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
B.C. Prasad
Central Tasar Research and Training Institute, Central Silk Board, Government of India, Piska Nagri PO, Ranchi-835 303, Jharkhand, India
Journal of Entomology
Year: 2010 | Volume: 7 | Issue: 3 | Page No.: 160-167







ABSTRACT
Assorted F1 hybrids and reciprocals of divergent tasar ecoraces of Antheraea mylitta viz., Daba, Jata and Raily were evaluated for relative heterosis. The DabaxJata [R] (random female and male) and its reciprocal hybrids have recorded high positive heterosis for fecundity (+17.45 and +1.70%), egg fertility (+11.58 and +7.20%), shell weight (+34.76 and +27.44%), silk ratio (+30.49 and +24.15%) and silk yields (+94.33 and +82.51%), respectively. Dabax Jata [PxS] (high pupal female and high shell male) hybrid, although recorded positive heterosis for all traits, but was next to random parental hybrids. The DabaxJata [PxP] (high pupal female and high pupal male) combination and its reciprocal have shown positive heterosis only for fecundity (+22.13 and +35.89%), while DabaxJata [SxS] (high shell female and high shell male) hybrid and its reciprocal are positive in shell weight (+45.12 and +33.54%) and silk ratios (+26.95 and 27.95%). All hybrids of DabaxRaily including reciprocals have shown negative heterosis for fecundity and silk yields. However, DabaxRaily [R], [SxS], [PxS] hybrids and their reciprocals have shown uneven positive heterosis for egg fertility (+1.65 to +20.74%), shell weight (+7.56 to +56.98%) and silk ratios (+1.02 to +54%). In general, all reciprocal F1 hybrids of Daba, Jata and Raily ecoraces have shown lesser heterosis. The dissimilar performance of assorted F1 hybrids and reciprocals of Daba, Jata and Raily ecoraces reveal their varied potential on relative heterosis. However, the study infers commercial prospective and optimal seed cocoon expediency in Daba and Jata ecoraces as F1 hybrids [R] and reciprocals. While, the trait specific positive heterosis in high pupal and high shell hybrids (assorted F1 hybrids) have application in segregating lines with desired traits and aggregating them in to needy one(s).
PDF Abstract XML References Citation
How to cite this article
R. Manohar Reddy, Suresh Rai, A.K. Srivastava, P.K. Kar, M.K. Sinha and B.C. Prasad, 2010. Heterosis Pattern and Commercial Prospective of Assorted F1 Hybrids of Indian Tropical Tasar Silkworm, Antheraea mylitta Drury. Journal of Entomology, 7: 160-167.
DOI: 10.3923/je.2010.160.167
URL: https://scialert.net/abstract/?doi=je.2010.160.167
DOI: 10.3923/je.2010.160.167
URL: https://scialert.net/abstract/?doi=je.2010.160.167
INTRODUCTION
The biological selection of parents on specific trait will lead to explicit individuals by their progeny in later generations, an advantage to have specific genotype of breeding application (Aruga, 1994; Singh et al., 2002; Moorthy et al., 2007). The strong heterosis and large variation among assorted hybrids supports further improvement of breed as per requirement (Li et al., 1998; Reddy et al., 2008). The crossbreeding is a simple method to increase health and efficiency of many animals causing heterosis by introducing and maintaining gene interactions (Siddiqui and Sengupta, 1993; Nagaraju et al., 1996; Puttaraju and Rajanna, 1997; Moghaddam et al., 2005). To develop productive hybrids and high yielding segregants in Antheraea mylitta, the pre requisites are selection of genetically diverged parental ecoraces, identification of commercial traits with heterotic potential (Sengupta et al., 1987; Siddiqui, 1997). The Daba ecorace reported as good combiner for silk yield and its F1 hybrids have shown better heterosis through dominance of gene action (Suryanarayana et al., 1987; Siddiqui et al., 1988), while silkworm crosses of congenic and syngenic lines provide better heterosis over conventional hybrids (Verma et al., 2005). The DabaxModal and Raily x Laria hybrid combinations recorded high heterosis in absolute silk yield with collective effect of fecundity, egg hatching, Effective Rate of Rearing (ERR) and shell weight (Siddiqui, 1997; Sinha et al., 2001; Singh et al., 2002). The variation in temperature, nutritional status, feeding duration and larval density influence body plasticity, thus the performance of breed and its voltinism (He and Wang, 2006; Hansda et al., 2008) and environment regulates the genotypic expression to produce different phenotypes (Zhao et al., 2007; Kumar et al., 2008). However, the information on parental selection with desired traits for hybridization and their trait related heterotic pattern in tropical tasar silkworm is very scanty. Hence, the hybridization conducted with parents of specific pupal and shell values using divergent (semi domesticated and wild) geographic ecoraces i.e., Daba (Jharkhand), Jata (Orissa) and Raily (Chhattisgarh) to cram the potential of heterosis on egg and silk related traits for commercial application.
MATERIALS AND METHODS
The study including the generation of parental seed cocoons, preparation of assorted F1 hybrids and their field evaluation have been carried out on economic plantation, Terminalia tomentosa (Wand A) at field laboratory of Central Tasar Research and Training Institute (CTR and TI), Ranchi, India successively for two years during 2007 and 2008.
Generation of Parental Seed Cocoons
The parental stocks of ecoraces viz., Daba, Jata and Raily of tasar silkworm, Antheraea mylitta were raised during the seed crop rearing season (July-August) of two successive years following integrated package of tasar silkworm rearing in a Randomized Complete Block Design (RCBD) with three replications on economic plantation, Terminalia tomentosa at field laboratory of Central Tasar Research and Training Institute, Ranchi, to minimize the problems of erratic emergence and non synchronization in moth coupling (experienced with wild stocks collected from nature), while preparing the Disease free layings (Dfls) of varied F1 hybrid combinations and reciprocals. The stocks of three parents were maintained for further experimentation for two years over seed and commercial crop seasons.
Preparation of Assorted F1 Hybrids
The ecorace wise male and female cocoons are segregated based on pupal and shell weights and preserved in separate cocoon cages at the grainage house of the research institute. The male and female moths emerged out of said non-diapausing cocoon batches of three divergent ecoraces were used for preparation of Dfls following integrated package of tasar silkworm seed production. The ecorace wise selfing, general and selective F1 hybrid combinations were prepared along with their reciprocals based on high pupal [PxP], high shell [SxS] and high pupal and high shell [PxS] weights along with control i.e., random parents [R] at grainage house of CTR and TI, Ranchi during September for two successive years for their evaluation in commercial crop (October-December) season.
Evaluation of F1 Hybrids and Reciprocals
The four general and twelve selective F1 hybrid and reciprocal combinations along with three parents, nineteen in totals, were reared in a Randomized Complete Block Design (RCBD) with three replications for each treatment on economic plantation, Terminalia tomentosa at field laboratory of CSR and TI, Ranchi, India in commercial crop rearing season (October-December) for two successive years. The total number of eggs, fertilized eggs of one Dfl and silk yield based on average shell weight and number of cocoons harvested per Dfl in respect of parents as well as hybrids were considered as one replication and single shell weight, silk ratios were calculated with equal number (twenty) of random cocoon samples. The observations recorded on fecundity, egg fertility, shell weight, silk ratio and silk yield were subjected for statistical analysis.
RESULTS
Comparative Performance (in situ) of Parental Ecoraces
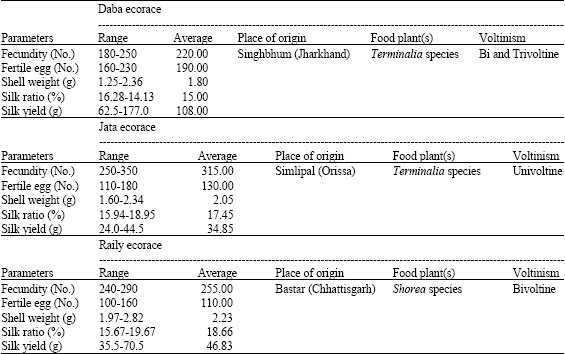
The performance levels of parental ecoraces viz., Daba, Jata and Raily under in situ habitats (Table 1) show variations in respect of their place of origin, food plants and voltinism in addition to commercial traits like fecundity, egg fertility, shell weight, silk ratio and silk yield. This indicates substantial phenotypic variability and genetic diversity besides their nativity in chosen parental material for hybridization. The semi domesticated Daba parent show superiority in egg fertility (190 No.) and silk yield (108 g) in spite of lesser fecundity (220 No.), shell weight (1.80 g) and silk ratios (15.00%). While, the other two wild ecoraces Jata and Raily could not compete with Daba in respect of silk yield (main commercial parameter), though they posses better fecundity (315 and 255 No.), shell weight (2.05 and 2.23 g) and silk ratios (17.45 and 18.66%) because of their low egg fertility (130 and 110 No.) and silk yield (34.85 and 46.83 g), respectively.
| Table 1: | Performance levels of parental ecoraces (Daba, Jata and Raily) under in situ habitation |
 | |
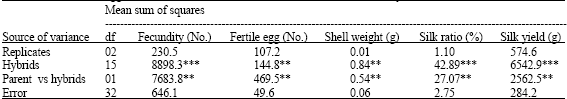
| Table 2: | ANOVA for egg and cocoon commercial traits of Daba ecorace and its F1 hybrids |
 | |
**Significant at 1% level, ***Significant at 0.1% level | |
| Table 3: | Relative heterosis (%) in F1 hybrids and reciprocals of Daba and Jata ecoraces |
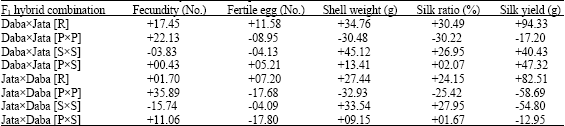 | |
[R] = Random female and male, [PxP] = High pupal weight female and high pupal weight male, [SxS] = High shell weight female and high shell weight male, [PxS] = High pupal weight female and high shell weight male | |
The ANOVA among the common parent of hybrids, i.e., Daba with F1 hybrids and reciprocals (Table 2), indicates significant variability for all characters viz., fecundity, egg fertility, shell weight, silk ratio and silk yield. This indicates remarkable relative heterosis potential in varied F1 hybrids along with reciprocals over semi-domesticated and commercially exploited Daba parental ecorace. While, the mean sum of squares in respect of parents versus hybrids, which is the measure of heterosis potential, found significant in egg and silk commercial traits, emphasizing the imminent of parental specificity and varied crossings (assorted F1 hybrids and reciprocals) on heterotic expression for commercial traits.
Performance F1 Hybrids of Daba and Jata Ecoraces
The pattern of heterosis among the parents Daba and Jata along with their reciprocals (Table 3) indicate superior performance in DabaxJata [R] and its reciprocal followed by high pupal and high shell combination of DabaxJata [PxS] and high shell combination of DabaxJata [SxS]. The relative heterosis recorded positive in five traits studied viz., fecundity (+17.45 and +1.70%), egg fertility (+11.58 and +7.20%), shell weight (+34.76 and +27.44%), silk ratio (+30.49 and +24.15%) and silk yields (+94.33 and +82.51%), respectively in DabaxJata [R] and its reciprocal hybrid combinations. The high pupal and high shell combination, JataxDaba [PxS] has shown lesser positive heterosis in fecundity (+0.43%), egg fertility (+5.21%), shell weight (+13.41%), silk ratio (+2.07%) and silk yield (+47.32%). The DabaxJata [PxP] combination and its reciprocal have recorded positive heterosis in fecundity (+22.13% and +35.89%). The DabaxJata [SxS] combination recorded positive heterosis in shell weight (+45.12%), silk ratio (+26.95%) and silk yield (+40.43%) while its reciprocal, JataxDaba [SxS] is positive in shell weight (+33.54%) and silk ratio (+27.95%). The reciprocal F1 hybrid JataxDaba [PxS] has shown positive heterosis in fecundity (+11.06%), shell weight (+9.15%) and silk ratio (+1.67%) and negative in egg hatching and silk yield (-17.80 and -12.95%).
Performance F1 Hybrids of Daba and Raily Ecoraces
The pattern of heterosis among the parents Daba and Raily along with their reciprocals (Table 4) indicate positive performance only in egg hatching, shell weight and silk ratios while they recorded negative in fecundity and silk yields.
| Table 4: | Relative heterosis (%) in F1 hybrids and reciprocals of Daba and Raily ecoraces |
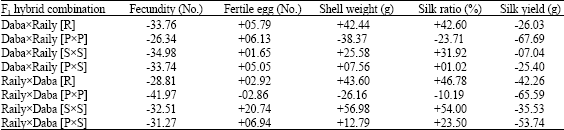 | |
[R] = Random female and male, [PxP] = High pupal weight female and high pupal weight male, [SxS] = High shell weight female and high shell weight male, [PxS] = High pupal weight female and high shell weight male | |
The DabaxRaily [R], DabaxRaily [SxS], DabaxRaily [PxS] and their reciprocals are positive in heterosis for egg hatching (+1.65 to +20.74%), shell weight (+7.56 to 56.98%) and silk ratios (+1.02 to 54.00%). The high pupal hybrid, DabaxRaily [PxP] shown marginal positive heterosis only in egg hatching (+6.13%), while it’s reciprocal RailyxDaba [PxP] was negative in all traits. The best performance among the DabaxRaily hybrids was exhibited by reciprocal hybrid combination, RailyxDaba [SxS] with positive heterosis in egg hatching (+20.74%), shell weight (+56.98%) and silk ratio (+54.00%) followed by reciprocal of RailyxDaba [R] and DabaxRaily [R] F1 hybrids.
DISCUSSION
The use of phenotypic variability in parents can build up a hybrid, exploiting heterosis to augment productivity and quality (Singh et al., 2002) and crossing of high x low, high x medium and medium x low yielding parents show positive heterosis (Siddiqui and Sengupta, 1993). In most of conventional heterosis breeding programmes, the geographical and phenotypic diversity is considered for available genetic distance among secluded populations. The other way of assessing the genetic diversity is with expression of heterosis in a particular hybrid as it demonstrates the existing degree of genetic diversity in parents. The phenotypic diversity though occurs due to genetic structure, the genotype environment (GxE) interactions at times manifest phenotypic variations. The reason for selecting Daba, Jata and Raily ecoraces was their varied yielding pattern in commercial trait(s) and their origin from semi domesticated and wild habitats. The parental ecoraces from divergent eco-geographic areas (Jharkhand, Orissa and Chhattisgarh states of India) with higher phenotypic diversity can magnify the inherent expression of heterosis at F1 level is the other reason for choosing these three parental ecoraces for the study. While, the selection of individual parents based on high pupal and high shell weights (for making varied F1 hybrids) was due to close association of bigger pupae with egg fecundity and higher cocoon shell with shell weight and silk yield, as these three traits are commercially important.
The higher positive relative heterosis in egg and cocoon commercial traits of DabaxJata [R] and its reciprocal indicates that Daba and Jata are best combiners and these findings are in conformity with earlier reports on Daba ecorace as best combiner (Suryanarayana et al., 1987; Siddiqui et al., 1988). This has commercial applicability in tasarculture as both parents can be utilized reciprocally for maximum egg output with minimum quantity of seed cocoons, which helps in-time supply of commercial seed with least cost of production. The positive heterotic gain among randomly coupled parents, Daba and Jata in F1 hybrids and reciprocals might be due to epistasis effect (Falconer, 1985; Aruga, 1994), while the positive heterosis in egg fecundity and shell weight might be due to dominant and over dominant gene(s) expression for a character (Verma et al., 2005; Reddy et al., 2008). Though, the F1 hybrid combination, DabaxJata [PxS] could show heterosis in egg and cocoon commercial traits, it could not achieve the production status of random parental hybrids in silk yield due to lesser levels of heterosis in fecundity, egg fertility, shell weight and silk ratio traits.
The expression of highly varied and in consistent heterosis by DabaxRaily and its reciprocal F1 hybrids might be due to hybrid weakness with non-allelic interaction which also can decrease heterosis. The negative heterosis for fecundity and silk yield of these hybrids coincides with inferior performance of one of its parental ecorace i.e., Raily under ex situ conditions with its non amenability and non acclimatization to changed environment and food plant; as it survives better under natural (in situ) habitat and prefers to feed on Shorea robusta. The major share of country’s tasar raw silk production from natural grown Raily cocoons, although indicate its production potential, but they are only under its natural habitat. Though, Daba parent emerged as best general combiner for most characters, the DabaxRaily hybrid is best reported for shell weight and silk ratios (Suryanarayana et al., 1987) and in present study, the RailyxDaba [SxS] and [PxS] combinations (reciprocals) have also shown same trend. In spite of positive heterosis for egg fertility, shell weight and silk ratio, the DabaxRaily and its reciprocal hybrid combinations could not enhance the economically important total silk yield due to higher negative heterosis in fecundity. This low fecundity has nullified the marginal positive heterosis of egg fertility and made these F1 hybrid combinations economically non-viable.
The low fecundity and egg fertility with better cocoon commercial traits and poor cocoon commercial characters with better fecundity and egg fertility prevailing in several ecoraces is one of the major constraints of commercial tasarculture. The potential of selection response in chosen character of parental herd was found improved in offspring generation; however, it basically depends on genetic variation existing among parents, selection intensity and accuracy induced while choosing the parents (Siddiqui and Sengupta, 1993; Reddy et al., 2008). The better relative heterosis recorded for fecundity in F1 hybrid combinations of JataxDaba [PxP], high pupal parents and its reciprocal indicate the selection response in chosen trait. Constructive modification of a breed for enhanced production needs the parental races of desired commercial traits (Falconer, 1985) and higher breeding magnitude of silk moth from heavier pupae laying more eggs (Singh et al., 1994; Reddy et al., 2009) might be the reasons for enhanced fecundity. This approach of improving/evolving a line of higher fecundity has applicability against persistent problem of low fecundity in many ecoraces of tropical tasar silkworm, which were otherwise good in remaining cocoon commercial traits. Also, the positive relative heterosis shown by selective hybrid combinations and reciprocals of DabaxJata [SxS] and DabaxRaily [SxS] in respect of shell weight and shell ratio indicates the positive heterotic response of parents with high shell weights. The better positive heterosis for shell weight and silk ratio also might be due to positive heterotic effect and better combinability of parents for yield contributing traits (Suryanarayana et al., 1987; Naqvi et al., 2004) significant correlation of a genotypic with phenotypic expression in respect of shell weight (Siddiqui and Sengupta, 1993), besides the dominance and over dominance in gene(s) expression for a chosen commercial trait (Verma et al., 2005).
Evolving a breed of high egg and silk yield traits depends on parental selection, effective utilization and adoption of apt breeding procedure (Moorthy et al., 2007). The objective of improving either fecundity or shell weight or both is to attain overall gain in silk yield of ecorace for its commercial sustenance. But, such selection of one or few economic character(s) results negatively with correlated changes on other commercially important character(s) or performance of breed (Moghaddam et al., 2005) and this might be the reason for decrease either in fecundity or in shell weight among selective (high pupal and high shell combinations) F1 hybrids. However, these combinations can be improved again to some extent by subsequent crossings with another strain of higher fecundity or better shell weight (Aruga, 1994). The productivity enhancement in all commercial or economically important traits is not achievable as productivity and survivability of a breed are negatively correlated (Moorthy et al., 2007) hence, a line moderately balancing both and better over existing on hand races should be designed. However, the segregated lines (assorted F1 hybrids) with special characters have certain application for a specific season, breeding purpose, to infuse and balance a line with needy commercial trait(s).
CONCLUSION
In general, all reciprocal F1 hybrids of Daba, Jata and Raily ecoraces have shown lesser heterosis. The dissimilar performance of assorted F1 hybrids and reciprocals of Daba, Jata and Raily ecoraces reveal their varied potential on relative heterosis. However, the study infers commercial prospective and optimal seed cocoon expediency in Daba and Jata ecoraces as F1 hybrids [R] and reciprocals. While, the trait specific positive heterosis in high pupal and high shell hybrids (assorted F1 hybrids) have application in segregating lines with desired traits and aggregating them in to needy one(s).
REFERENCES
- Hansda, G., R. Manohar-Reddy, M.K. Sinha, N.G. Ojha and N.B.V. Prakash, 2008. Ex situ stabilization and utility prospects of Jata ecorace of tropical tasar silkworm Antheraea mylitta Drury. Int. J. Indus. Entomol., 17: 169-172.
Direct Link - He, X.Z. and Q. Wang, 2006. Asymmetric size effect of sexes on reproductive fittness in an aphid parasitoid Aphidius ervi (Hymenoptera: Aphidiidae). Biol. Control, 36: 293-298.
Direct Link - Li, B., G.T. Howe and R. Wu, 1998. Developmental factors responsible for heterosis in aspen hybrids (Populus tremuloides, P. tremula). Tree. Physiol., 18: 29-36.
Direct Link - Moorthy, S.M., S.K. Das, N.B. Kar and S.R. Urs, 2007. Breeding of bivoltine breeds of Bombyx mori suitable for variable climatic conditions of tropics. Int. J. Ind. Entomol., 14: 99-105.
Direct Link - Reddy, R.M., N. Suryanarayana and N.B.V. Prakash, 2008. Heterosis potential in selective parental F1 hybrids of divergent geographic ecoraces of tropical tasar silkworm, Antheraea mylitta D (Lepidoptera: Saturniidae). Acad. J. Entomol., 1: 32-35.
Direct Link - Reddy, R.M., N. Suryanarayana, N.G. Ojha, G. Hansda, S. Rai and N.B.V. Prakash, 2009. Basic seed stock maintenance and multiplication in Indian tropical tasar silkworm Antheraea mylitta Drury: A strategic approach. Int. J. Ind. Entomol., 18: 69-75.
Direct Link - Siddiqui, A.A., 1997. Studies on heterosis and heterobeltiosis in the tasar silkworm, Antheraea mylitta D. Sericologia, 37: 59-69.
Direct Link - Singh, R., H.K. Chaturvedi and R.K. Datta, 1994. Fecundity of mulberry silkworm, Bombyx mori in relation to female cocoon weight and repeated matings. Indian J. Sericult., 33: 70-71.
Direct Link - Suryanarayana, N., K. Sengupta and B.N. Bramhachari, 1987. Heterosis and combining ability in Indian tasar silk worm, Antheraea mylitta D. Sericologia, 27: 701-709.
Direct Link - Verma, A.K., G.K. Chattopadhyay, M. Sengupta, S.K. Das and A.K. Sarkar, 2005. Heterobeltiotic genetic interaction between congenic and syngenic breeds of silkworm, Bombyx mori L. Int. J. Ind. Entomol., 11: 119-124.
Direct Link - Zhao, Y., K. Chen and S. He, 2007. Key principles for breeding spring-and-autumn silkworm varieties: From our experience of breeding 873x874. Caspian J. Environ. Sci., 5: 57-61.
Direct Link