
Mohammad A. Al-Deghairi
Department of Crop Production and Protection, College of Agriculture and Veterinary Medicine, Qassim University, P.O. Box 6622, Buraydah 51452, Kingdom of Saudi Arabia
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 2 | Page No.: 72-81







ABSTRACT
Combined effects between the entomopathogenic fungi, Beauveria bassaina (Bals.) and whitefly parasitoid, Eretmocerus mundus Mercet on Bemisia tabaci (Genn.) were investigated under laboratory conditions. The competitive interactions among them were also evaluated either alone or in combination, in respect of the positive and negative effects. The deleterious effects on the parasitoid were extremely low, particularly when the parasitized nymphs exposed to the fungus later. In direct contact bioassay, fungus caused 5.1-15.3% mortality in post-releasing trial and from 8.9-22.1% in pre-releasing trial. Three to five days after treatment, B. tabaci nymphs were rejected as a host by E. mundus females due to the fungal infection. In infected nymphs, the majority of E. mundus females were not laid and no parasitism was detected. The control efficiency of the two natural enemies of B. tabaci, when used separately or in combination, varied according to the tested biological agent. E. mundus alone reduced pest populations by 19.4 and 51.1% in pre- and post-releasing trials, respectively. B. bassaina caused 38.1% in pre-releasing trial and 29.4% in post-releasing trial. Meanwhile, the interaction between fungus and the parasitoid in combination reduced the pestpopulation by 51.2 and 72.3% in pre- and post-releasing trials, respectively.
PDF Abstract XML References Citation
How to cite this article
Mohammad A. Al-Deghairi, 2009. Combining Effect of Beauveria bassaania (Bals.) and Ertemocerus
mundus Mercet (Hymenoptera: Aphelinidae) on Sweetpotato Whitefly,
Bemisia tabaci Gennadius (Aleyrodidae; Hemiptera). Journal of Entomology, 6: 72-81.
DOI: 10.3923/je.2009.72.81
URL: https://scialert.net/abstract/?doi=je.2009.72.81
DOI: 10.3923/je.2009.72.81
URL: https://scialert.net/abstract/?doi=je.2009.72.81
INTRODUCTION
The protected environment in controlling whiteflies appears to be an ideal for successful application of biological control agent. This can be gained by combining the effect of pest natural enemies to enhance their efficiency and hence, to provide environmentally safer methods than using chemicals (Ehler, 2000). In this respect, Cory and Hoover (2006) showed that both entomopathogenic fungi and insect parasitoids and/or predators of arthropod natural enemies can contribute in suppressing pest populations when used jointly or independently. To encourage the role of whitefly natural enemies in pest control, they should be tested through interaction with other natural enemies in the agro-ecosystem.
Role of this fungi as virulence agent in suppressing whitefly populations was studied by Al-Deghairi (2008). The suitability of B. bassaina, as part of whitefly control measures in IPM programs, is critically important. Therefore, the fungus, B. bassaina and the whitefly parasitoid, E. mundus, have the potential to complement or interfere with each others, based on the environmental conditions and other biological factors (Kim et al., 2005). This entomopathogenic fungi should be compatible with other naturally occurring biological control agents, such as, the hymenopteranparasitoids, Eretmocerus mundus, in order to maximize the efficiency of the biological pest control. However, most experiments have focused on the effect of a particular biological agent and availability of data on susceptibility of parasitoids to the entomopathogenic fungi, B. bassaina, in particular, are insufficient. In addition, there is no information available regarding the non-target effects of B. bassaina on this parasitoid.
Since, new biotypes of Bemisia tabaci have very high reproductive rates and are very difficult to control with a single biological control agent, various species of natural enemies may be introduced simultaneously in greenhouses. These biological control agents may act synergistically, additively or antagonistically. Moreover, insect natural enemies have developed to be employed in multi-trophic relations, it is important to assess their interactions within natural enemies complexes if they are used in combination in IPM programs (Roy and Pell, 2000). Thus, in order to use insect natural enemies more effectively in IPM program, they should act in harmony with minimal antagonistic interaction between groups and other interventions (Lacey et al., 1997). Multiple species of natural enemies can interact either synergistically/additively or antagonistically (Ferguson and Stilling, 1996; Roy and Pell, 2000). Obviously, Roy and Pell (2000) concluded that the synergistic interactions among natural enemies may lead to higher mortality than the combined individual mortalities of the pest populations. Meanwhile, the additive mortality occurs if the biological agents do not interplay and consequently, the total mortality levels by various agents are generally equal. This study deals with laboratory trials to investigate the susceptibility of interaction between E. mundus and B. bassaina, the widely used natural enemies for whitefly control. Thus the goal of this study focuses on the interactions between the entomopathogenic fungi, B. bassaina and the internal whitefly parasitoid, E. mundus. The capability of B. bassaina and/or E. mundus alone and in combination in controlling B. tabaci populations is also determined.
MATERIALS AND METHODS
Maintenance of Experimental Organisms
The sweetpotato whitefly was reared on the kidney bean, Phaseolus vulgaris L., under laboratory conditions 25±2.2°C and 70±5% R.H., College of Agriculture and Veterinary Medicine, Qassim University, KSA. The pest culture was originally initiated from individuals collected from squash plants, free of insecticides, in Qassim region. About hundred plastic pots cultivated with P. vulgaris were maintained until formation of two fully expanded leaves. Ten pairs of B. tabaci adults were introduced to the kidney bean leaves, confined under screen cages for 48 h in order to lay their eggs and then removed. The eggs were counted on each leaf and left till hatching and transformed to the 2nd instar nymphs.
Theparasitoid was collected from different plant hosts free of insecticides and its colony was initiated on P. vulgaris plants infested by whitefly nymphs and kept under laboratory conditions at 25±2.2°C; 70±5% R.H. under normal photoperiod. To obtain adult parasitoids, the parasitoid pupae were isolated for adult emergence using small tubes. The adults were fed on honey solution and were placed on newly plants infested with whitefly nymphs. The fungus was isolated from naturally infected whiteflies according to Abdel-Baky (2000) and Al-Deghairi (2008) and kept in slant Agar media at 5°C. The fungal spores were harvested from two weeks old cultures on autoclaved PDA media at 28±2°C by rinsing with sterilized distilled water.
Combined Effects of B. bassaina and E. mundus on B. tabaci
In order to assess the combining effect between the entomopathogenic fungus, B. bassaina and E. mundus, two trials were designed, included: whitefly nymphs treated with fungal suspension pre-releasing the parasitoid and whitefly nymphs treated with fungal suspension post-releasing the parasitoid. In each trial, thirty kidney bean pots infested by constant numbers of whitefly nymphs (40 nymphs leaf-1) were used. Three fungal concentrations were applied (Al-Deghairi, 2008) to assess the positive and negative impact on the parasitoid development and parasitism percentage. The trials were preformed as follows:
Pre-Releasing Trial
Sixty kidney bean pots infested by whitefly nymphs were used in this trial, divided to two groups and treated with three fungal concentrations of 2x106, 4x106 and 6x106 spores mL-1. Twenty pots were used to each fungal concentration. A ratio of 1 parasitoid: 5 whitefly nymphs was released at two intervals. One group of the parasitoid were released immediately after treated by the fungal concentration, the other were released three days after the treatment. The parasitoid oviposition process behavior, symptoms of parasitism and parasitoid mortality by the fungus were studied carefully. Emergence of parasitoid adults was also recorded.
Post-Releasing Trial
Sixty plastic pots planted with kidney beans were also used in this trial. A constant number of the parasitoid (1 parasitoid: 5 WF nymphs) was introduced to kidney beans plants infested with whitefly 2nd instar under screen cages. The parasitoids were left for 24 h to oviposit their eggs and then removed. The pots were also divided to two groups; one group was treated, three days later, by fungal suspension at concentrations of 2x106, 4x106 and 6x106 spores mL-1. The other group, was treated by the fungal suspensions after five days from removing the parasitoid adults. Two days later after introducing the fungus, kidney bean leaves infested by WF nymphs were investigated by hand lens and examination was continued until parasitoid adult emergence. The mortalities of WF nymphs and emerging parasitoid adults by the fungus were observed under each fungal concentration. The obtained data were compared with the check treatment (treated by distilled water only).
Statistical Analysis
The mortalities number and values in all tests were subjected to ANOVA analysis. All statistical analysis were preformed using CoStat Software program (1990). The percentages of mortality due to fungal activities were calculated according to Abbott (1925). The interaction between the two bio-control agents and contribution of each biological agent if used alone or in combination to regulate the pest population were calculated by MINITAB program.
RESULTS
Beauveria bassaina on Bemisia tabaci Nymphs
Beauveria bassaina caused different mortality rates among nymphs of B. tabaci. These mortality percentages were significantly different (p<0.05) based on the fungal concentrations (Fig. 1). Infection levels were generally greater when higher spore concentrations of the fungus was used. All tested conidial concentrations were pathogenic and highly virulent among B. tabaci nymphs (Fig. 1). The average mortality percentages among B. tabaci nymphs were 18.75, 24.6 and 46.5 with 2x106, 4x106 and 6x106, respectively. This means that mortality among B. tabaci nymphs increased with the increase of the fungal concentrations and thus, sufficient control could be achieved when higher conidial concentrations were applied. The statistical analysis also revealed that nymphal mortality varied significantly among fungal concentrations and insect life stages (Fig. 1).
Eretmocerus mundus on Bemisia tabaci
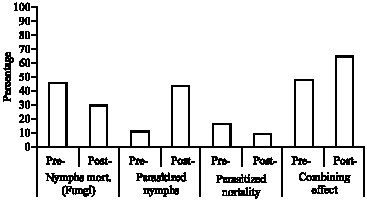
Releasing E. mundus against varied pest densities affected the control levels of B. tabaci (Fig. 5). Desirable control levels were achieved when higher number of parasitoids are released specially when low number of the pest is present. About 43.57% mortalities among B. tabaci population were achieved when releasing the parasitoids before the fungal treatments, while mortality reached 12.08% among whitefly nymphs that were treated by the fungus before introducing the parasitoids (Fig. 5).
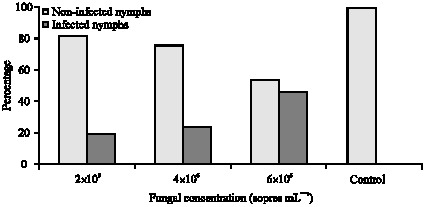 | |
| Fig. 1: | Efficiency of Beauveria bassaina against Bemisia tabaci nymphs |
| Table 1: | Summarization of the interacting between Beauveria bassaina and Eretmocerus mundus in controlling Bemisia tabaci |
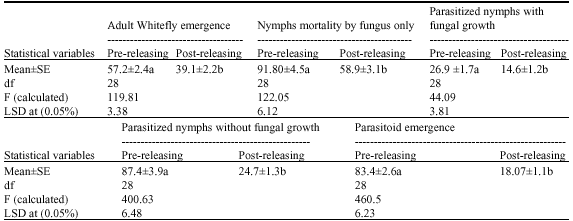 | |
| aThe numbers followed by the same letter(s) within a widest column are not significantly different at 5% level | |
Combined Effect of B. bassaina and E. mundus on B. tabaci Nymphs
Table 1 show the potential combining effect of two natural enemies of sweetpotato whitefly, B. tabaci, assessing five factors in two trials. The first factor was the emerged number of whitefly adults, which was high in pre-releasing trial compared with post-releasing trial. At least 57.2±2.4 WF adults, of 200 individuals, were emerged in pre-releasing trial compared to 39.1±2.2 adults in post-releasing trial (Table 1). The 2nd factor was the number of infected WF nymphs by B. bassaina alone, which gave 91.80±4.5 and 58.9±1.7 nymphs of 200 individuals, for pre- and post-releasing trials, respectively. The 3rd and 4th factors were related to the effect of E. mundus, which gave 87.4±3.9 and 24.7±1.3 of parasitized WF nymphs in pre- and post- releasing trials, respectively. Consequently, the number of emerged parasitoid was 83.4±2.6 and 18.07±1.1 in pre-releasing and post-releasing trial, respectively (Table 1). The last factor was dealing with the negative interaction between the two biological agent. This factor assessed weather the fungus affected the parasitization or the population of the parasitoid. In pre-releasing trial, the fungus destroyed 26.9±1.7 parasitized WF nymphs and destroyed 14.6±1.2 nymphs in post-releasing trial (Table 1).
Negative Impact of the Interaction Among the Two Biological Control Agents
Indirect Contact
Beauveria bassaina could destroy the parasitoid in its embryonic phase as it penetrates the pest cuticle either mechanically or biologically, therefore, the fungus could affect the emerged numbers of the parasitoid adults. In this respect, Fig. 2 shows the percentages of emerged and non-emerged parasitoids adults under three fungal concentrations at two time intervals of treatments. In pre-releasing trial, the percentages of non-emerged parasitoid adults were 1.3, 4.9 and 8.6%, with 13.57% for check treatment, with concentrations set at 2x106, 4x106 and 6x106 spores mL-1, respectively. Meanwhile, in post-releasing trial, the low spore concentrations 2x106 (spores mL-1) caused 17.5% mortalities of the parasitoid in the embryonic phase, followed by moderate concentrations 4x106 (spores mL-1) which caused 28.86%. On the other hand, higher fungal concentrations 6x106 (spores mL-1) resulted to 34.89% mortalities in parasitoid at the embryonic developmental phase. Parasitoids mortality in the embryonic developmental phase was differed significantly based on the time of fungal treatments and fungal concentrations (Fig. 2).
Direct Contact
Parasitoids may succeed in avoiding fungal infection during the embryonic phase, however, direct contact with the fungus which covers the body of whitefly nymphs during its emergence from the pupal case or during its search for non-infected hosts may contribute to its mortality. Mortality percentages in E. mundus populations in pre-releasing trial were 8.9, 14.8 and 22.1% for the three fungal concentrations used (2x106, 4x106 and 6x106 spores mL-1), respectively (Fig. 3). In the post-releasing trial, mortality percentages were 5.1, 9.2 and 15.3% for the three fungal concentrations used (2x106, 4x106 and 6x106 spores mL-1).
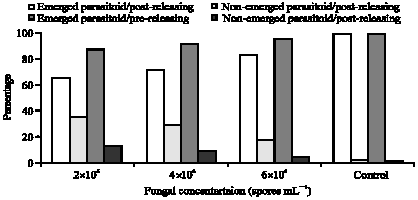 | |
| Fig. 2: | Negative impact of Beauveria bassaina on the whitefly parasitiod, Eretmocerus mundus under three fungal spore concentrations |
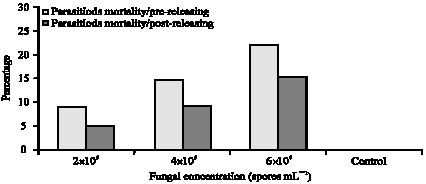 | |
| Fig. 3: | Mortality percentages among Eretmocerus mundus population treated by Beauveria bassaina |
 | |
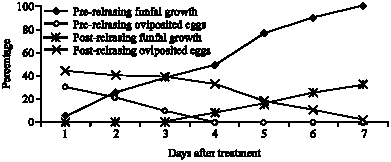
| Fig. 4: | Percentages of the fungal growth and the oviposetid eggs/female parasitiod in pre- and post releasing treatments |
 | |
| Fig. 5: | An Average impact of the interactions among Beauverava bassina and Eretmocerus mundus in controlling Bemisia tabaci |
In conclusion, the negative effects of B. bassaina on E. mundus population were very limited compared to chemical insecticides, if used in the appropriate time. Indirect moralities among the parasitoids in pre-releasing trial were very low and ranged from 4.9 to 13.57% when parasitized WF nymphs were treated by the fungus (Table 1). Meanwhile, when the parasitoid adults were treated by the fungal suspension (direct contact), they rejected the infected WF nymphs as a host. The majority of female parasitoids laid no eggs and in few cases showed a non-preference to infected nymphs (Fig. 4).
Positive Interaction Effect Between the Two Biological Control Agents
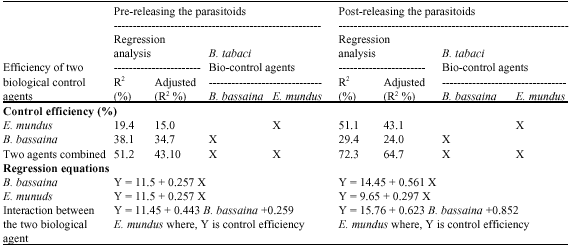
The entomopathogenic fungi, B. bassaina and the internal whitefly parasitoid, E. mundus, alone and together, were able to suppress B. tabaci population. In this respect, the regression analysis showed that sharing rates of each one of the two natural enemy when used alone or together in controlling the pest under laboratory conditions (Table 2, Fig. 5).
E. mundus alone decreased the pest populations by 19.4% in pre-releasing trial and 51.1% in the post-releasing trial (Table 2). Meanwhile, B. bassaina was able to reduce the pest population by 38.1 and 29.4% in both pre- and -post releasing trials, respectively. On the contrary, the combined effect of B. bassaina and E. mundus raised the control efficiency to 51.2% in pre-releasing treatment and 72.3% in the post-releasing trial. This means that both biological control agents of whitefly interact positively and enhanced the pest control efficiency, if applied in correct way particularly after releasing the parasitoid after four to five days (Fig. 4, 5).
| Table 2: | Regression analysis of the interactions among Beauveria bassaina and Eretmocerus mundus in controlling Bemisia tabaci either applied separately or in combinations |
 | |
Accepted and Rejected Whitefly Nymphs Treated by Beauveria bassaina as a Host by Eretmocerus mundus Females
Figure 4 shows percentages of fungal growth and number of oviposited eggs/female parasitoids when B. tabaci nymphs treated by the fungus before or after releasing the parasitoids. In pre-releasing trials, fungal growth was very low one day after treatment, giving only 5%. Meanwhile, the growth of the fungal was accelerated in the 2nd day giving a 25%, then increased sharply which reach its completed development within the 6th day of treatment. Thus, the fungal mycelium covered the cadavers of whitefly nymphs destroying them and making them un-healthy. Consequently, female parasitoids rejected the nymphs and never oviposit their eggs except in the first two days before the fungal mycelium covered the nymphal body (Fig. 4).
No fungal growth was observed within 1st three days after releasing the parasitoids. However, the development of the fungal growth was very low in comparison with the pre- releasing treatment. Although, whitefly nymphs were accepted as a host by parasitoids` female giving very high oviposition percentages ranged from 44 to 32% after four days from treatment, the female parasitoids rejected the hosts and number of oviposited eggs decreased sharply towards the end of the trial when the fungus was at its maximum growth (Fig. 4).
DISCUSSION
Entomopathogenic fungi may interact with other insect natural enemies either as individual species or as species complexes (Roy and Pell, 2000). Consequently, a competition may occur as interactions between closely related species or among unrelated organisms (Hochberg and Lawton, 1990). Increasing demands for entomopathogenic fungi and parasitoids in biological control programs, made it important to evaluate their interactions to maximize their efficiency in IPM programs (Pell and Roy, 2000; Lacey and Mesquita, 2002). Fungal pathogens have impact on the beneficial insects or non-target organisms either through direct and/or indirect infection of these organisms (Ehler, 2000). Direct infection of any beneficial insect or non-target organisms usually has undesirable effects in any biological control program. Moreover, Rosenheim et al. (1995) explained the indirect effects where a pathogen may interfere with the natural enemy complex by reducing the pest population or rendering the host pest unsuitable for other natural enemies. This completely in agreement with the current studies (Fig. 2, 3). B. bassaina caused direct and indirect mortalities among E. mundus populations and mortalities were differed based on the fungal concentrations.
When Beauveria bassaina and Eretmocerus mundus are applied separately or in together, it is important that they may be mutually compatible or interfere under field conditions. Due to its numerous hosts, B. bassiana infects various insect species (Al-Deghairi, 2008), thus, the fungus could potentially pose a threat to other beneficial insects and non-target organisms. Combined use of B. bassaina and E. mundus may be more efficient than being used separately resulting in complete mortality of the pest.
Interaction among B. bassaina and E. mundus was positively in favor of the insect pest control. Comparatively with the earlier studies, most interactions between entomopathogenic fungi and the insect parasitoid were asymmetrically in favor of the pathogen (Hochberg and Lawton, 1990). Nevertheless, the relative timing of parasitism and fungal infection is often crucial to the final competitive outcome.
In the current study, a higher percentage of emerged parasitoid adults were recorded in post-releasing treatment meaning that the parasitized host individuals were less susceptible to infection by the fungus than unparasitized ones. This may be due to the changes in the host caused by the parasitoid or its progeny after parasitism (Vinson and Iwantsch, 1980). This is generally defined as host regulation. Changes in parasitized hosts may include morphological, biochemical and physiological or physical activities within the nymphal stage, as well as, host cuticle melanization (Fransen and Van Lenteren, 1993, 1994). Additionally, the researchers correlated the changes in host susceptibility with the emergence of the parasitoid larvae from the eggs inside the host as happen in Encarsia formosa and Aschersonia alyerodis.
In general, Fransen and Van Lenteren (1993) shown the factors that may influence the effective colonization of parasitized greenhouse whitefly by E. formosa after treatment by A. alyerodis as follows: increasing parasitized hosts survival in post-releasing trials can induce a decrease in host susceptibility for infection, fungal penetration of the parasitized larvae may be more difficult than unparasitized hosts because of indirect changes in the host cuticle competition between unrelated organisms may be present and after successful penetration of the parasitized host, the fungus may be hampered due to the defense mechanisms. This latter interpretation was in agreement with Vinson (1976) and Vinson and Iwantsch (1980). The current study also shows that E. mundus females were able to discriminate between infected nymphs by B. bassiana, that have a mycelium growth and non-infected nymphs. This may explain the rejection of the parasitoid adults to use the infected WF nymphs as a host.
Successful development of the parasitoid and increasing its efficiency when combined with the fungal pathogen depends on the timing of fungal spore applications to the hosts of the parasitoids. This may be useful to determine whether the insect hosts were first infected by the pathogen or were first parasitized by the parasitoid as in case of Verticillium lecanii infecting Aphidius nigripes, a parasitoid of potato aphid, Macrosiphum euphorbiae (Askary and Brodeur, 1999). Fransen and Van Lenteren (1994) also reported that the infection rates of E. Formosa, a greenhouse whitefly parasitoid, by A. aleyrodis varied with the timing of spore application after parasitism. They found that when fungal spores were applied on parasitized whitefly nymphs one to three days after oviposition, the parasitized nymphs were significantly reduced as a result of cumulative infection by fungus and then by parasitism. However, when fungal spores were applied 4-10 days after introducing parasitoids, parasitism wasn`t reduced compared with the control. Finally, parasitization at an early phase of fungal infection is, accordingly, detrimental to the parasitoid progeny survival. In this study, E. mundus adults were able to discriminate between infected and non-infected WF nymphs at a later phase, gave the parasitoid the opportunity to successfully parasitize healthy WF nymphs and thereby to cause host mortality complementary to the fungal treatment. This complies with the results of Rosa et al. (2000) who mentioned that the high virulence of Metarhizium anisopliae and Beauveria bassaina to the bethylid parasitoid Prorops nasuta, did not affect significantly the predatory or parasitic capacity of P. nasuta.
Intra-guild predation is a dramatic expression on intervention among natural enemies, which could lead to antagonism and reduced host mortality and its dominance in the biological control agents (Polis and Holt, 1992; Rosenheim et al., 1995). Generally, the earlier studies on the interactions among entomopathogens and insect natural enemies consider the pathogen as the intra-guild predator which may directly able to infect the other natural enemies in the guild (Ehler, 2000).
In conclusion, E. mundus negatively affected the biological control potentiality of this fungus (Table 1). The successful augmentation of these natural enemies may be impeded by the antagonism, however, careful management ensuring temporal separation of the interacting natural enemies could result in effective biological control (King and Bell, 1978). In addition, the use of an interaction among biological control agents is required through understanding of the dynamic relationship between pathogens, parasitoids and host insects. Moreover, the successful manipulation of natural enemies in IPM program is dependent upon such an understanding. Therefore, the present study indicates that the fungus B. bassaina may be compatible with the action of the parasitoid or the predator under field conditions, giving that pathogen applications and parasitoid or/and predator releasing are timed not to coincide.
ACKNOWLEDGMENT
The researchers is very grateful to an anonymous reviewers for most valuable comments improving the manuscript considerably.
REFERENCES
- Abbott, W.S., 1925. A method of computing the effectiveness of an insecticide. J. Econ. Entomol., 18: 265-267.
CrossRefDirect Link - Abdel-Baky, N.F., 2000. Cladosporium spp. an entomopathogenic fungus for controlling whiteflies and aphids in Egypt. Pak. J. Biol. Sci., 3: 1662-1667.
CrossRefDirect Link - Al-Deghairi, M.A., 2008. Bioassay evaluation of the entomopathogenic fungi, Beauveria bassaina Vuellemin against eggs and nymphs of Bemisia tabaci Gennadius (Homoptera: Aleyrodidae). Pak. J. Biol. Sci., 11: 1551-1560.
CrossRefPubMedDirect Link - Askary, H. and J. Brodeur, 1999. Susceptibility of larval stages of the aphid parasitoid Aphidius nigripes to the entomopathogenic fungus Verticillium lecanii. J. Invertebr. Pathol., 73: 129-132.
CrossRefDirect Link - Ferguson, K.I. and P. Stilling, 1996. Non-additive effects of multiple natural enemies on aphid populations. Oecologia, 108: 375-379.
CrossRefDirect Link - Fransen, J.J. and J.C. van Lenteren, 1993. Host selection and survival of the parasitoid Encarsia formosa on greenhouse whitefly, Trialeurodes vaporariorum, in the presence of hosts infected with the fungus Aschersonia aleyrodis. Entomol. Exp. Applic., 69: 239-249.
CrossRefDirect Link - Van Lenteren, J.C. and J.J. Fransen, 1994. Survival of the parasitoid Encarsia formosa after treatment of parasitized greenhouse whitefly larvae with fungal spores of Aschersonia aleyrodis. Entomol. Exp. Applic., 71: 235-243.
CrossRefDirect Link - Hochberg, M.E. and J.H. Lawton, 1990. Competition between kingdoms. Trends Evol. Ecol., 5: 367-371.
CrossRef - Kim, J.J., K.C. Kim and D.W. Roberts, 2005. Impact of the entomopathogenic fungus Verticillium lecanii on development of an aphid parasitoid, Aphidius colemani. J. Invertebr. Pathol., 88: 254-256.
Direct Link - King, E.G. and J.V. Bell, 1978. Interactions between a bracoind, Microplitis crocepis and a fungus, Nomuraea rileyi, in laboratory-reared bollworms larvae. J. Invertber. Pathol., 31: 337-340.
Direct Link - Lacey, L.A., A.L.M. Mesquita, G. Mercadier, R. Debire, D.J. Kazmer and F. Leclant, 1997. Acute and sublethal activity of the entomopathogenic fungus Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes) on adult Aphelinus asychis (Hymenoptera: Aphelinidae). Environ. Entomol., 26: 1452-1460.
CrossRefDirect Link - Polis, G.A. and R.D. Holt, 1992. Intra-guild predation: the dynamics of complex trophic interactions. Trends Ecol. Eval., 7: 151-154.
CrossRef - Poppy, G.M., 1997. Tritrophic interactions: Improving ecological understanding and biological control? Endeavor, 21: 61-65.
CrossRefDirect Link - Cory, J.S. and K. Hoover, 2006. Plant-mediated effects in insect pathogen interactions. Trends Ecol. Evol., 21: 278-286.
CrossRef - Rosa de la, W., H.R. Segura, J.F. Barrera and T. Williams, 2000. Laboratory evaluation of the impact of entomopathogenic fungi on Prorops nasuta (Hymenoptera: Bethylidae), a parasitoid of the coffee berry borer. Environ. Entomol., 29: 126-131.
Direct Link - Rosenheim, J.A., H.K. Kaya, L.E. Ehler, J.J. Marois and B.A. Jaffee, 1995. Intra-guild predation among biological control agents: Theory and evidence. Biol. Control, 5: 303-335.
CrossRef - Roy, H.E. and J.K. Pell, 2000. Interactions between entomopathogenic fungi and other natural enemies: Implications for biological control. Biocontrol Sci. Technol., 10: 737-752.
CrossRef - Vinson, S.B. and G.F. Iwantsch, 1980. Host suitability for insect parasitoids. Annu. Rev. Entomol., 25: 397-419.
CrossRef