
T.O. Femi-Ola
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
E.Y. Aderibigbe
Department of Microbiology, University of Ado-Ekiti, P.M.B. 5363, Ado-Ekiti, Nigeria
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 1 | Page No.: 67-71







ABSTRACT
The bacterial population in the hindgut of the higher wood-eating termite, Amitermes evuncifer Silvestri was estimated at both dry and wet seasons. The total bacterial counts in the soldier and worker termites were 1.46±0.26x106 and 2.51±0.31x106 cfu mL-1 during the wet season; while it was 5.30±1.1x104 and 5.8±0.9x104 cfu mL-1 during the dry season, respectively. There was no significant difference in the total bacterial count in the hindgut of the worker and soldier termites in both seasons. The total bacterial count in the dry season was significantly lower (p≤0.05) than the population during the wet season. The bacterial species were identified to be Bacillus subtilis, B. cereus, Micrococcus luteus, Streptococcus sp. and Serratia marcescens.
PDF Abstract XML References Citation
How to cite this article
T.O. Femi-Ola and E.Y. Aderibigbe, 2009. Effects of Seasonal Changes on the Microflora In the Hindgut of Wood-Eating Termites. Journal of Entomology, 6: 67-71.
DOI: 10.3923/je.2009.67.71
URL: https://scialert.net/abstract/?doi=je.2009.67.71
DOI: 10.3923/je.2009.67.71
URL: https://scialert.net/abstract/?doi=je.2009.67.71
INTRODUCTION
Termites are social insects having three distinct castes, the reproductive (queen and kings), the soldiers and workers (Malaka, 1996; Roger et al., 2006). The workers are involved in the building and maintenance of the nests. They do all the foraging and care for the eggs and young as well as caring for the queen. The soldiers are defenders of the nests (Malaka, 1996). Gut microbes are very important for termite vitality and much of the termite nutrition is derived from products of microbial metabolism. Production of acetate, propionates and other organic acids during microbial fermentation of carbohydrates in the hindgut serves as important oxidizable energy source for termites (Todaka et al., 2007).
Until recently, hydrolysis and utilization of cellulose in amount sufficient to produce usable energy to an organism were thought to be carried out exclusively by microorganisms. It appears that some animal species including termites, nematodes and crayfish produce their own cellulases, which differ substantially from those of their indigenous micro flora (Bera-Maillet et al., 2000; Watanabe and Tokuda, 2001; Khademi et al., 2002). Wood-eating termites have caused substantial economic loses as a result of their destruction of wood edifices. Their nefarious activities are more pronounced during the wet/ rainy season. New frontiers are conquered and more nests are established during the wet season. There are two seasons in Nigeria; the rainy/wet and the dry/harmattan, which are characterized by different ambient temperature, amount of rainfall, relative humidity and intensity of sunshine. These environmental factors might affect both the population and types of microorganisms in the hindgut of these insects.
This study reports the effects of different seasons (dry and wet) on the population of the wood-eating termite, A. evuncifer Silvestri. The microbial load in the hindgut of workers and soldier termites were also compared.
MATERIALS AND METHODS
Geographical Parameters of the Study Area
The amount of total rainfall was monitored using a rainguage (Casela, London), while the temperature was determined with a dry bulb thermometer (Model 88982C Casela, London). The relative humidity of the study area was monitored with a whirling hygrometer (Model T8563/1, Casela, London).
Collection of Termites
Wood-eating termites were collected during the dry (December to March) and rainy (April to November) season from infested planks and bamboo in the University of Ado-Ekiti campus and environs. Termite-infested wood containing both the worker and soldier termites were moistened with distilled water and kept in plastic boxes prior to processing. Termite samples were identified as Amitermes evuncifer of the order Isoptera family Termidae at the Department of Zoology, University of Ado-Ekiti, Nigeria.
Dissection of Termite Samples
Termites were surface-sterilized in 10% (v/v) sodium hypochlorite and distilled water. Thereafter, they were rinsed in two changes of sterile distilled water. Each of them was aseptically dissected to remove the hindgut, which was then placed in 10 mL sterile ringers solution containing the following (g L-1): NaCl, 10.0; KCl, 0.42; CaCl2.2H2O, 0.48 and NaHCO3, 0.20. The gut sample was then crushed with a sterile glass rod to release the intestinal contents. Serial dilution of gut sample was carried out in sterile dilution blanks of distilled water and plated on Nutrient Agar (NA) plates and Potato Dextrose Agar (PDA) amended with penicillin (Ako-Nai et al., 1992). The replicate plates were incubated at 37°C for 2 days (NA) to 5 days (PDA), respectively. Colonies on plates were counted and multiplied with the dilution factor to determine the total microbial count. The mean of the total count was calculated and expressed as colony forming units per milliliter (cfu mL-1). Cultural characteristic of bacterial isolates on agar plates were recorded. Pure cultures of the isolates were obtained on agar plates obtained by sub-culturing serially onto sterile Nutrient agar plates.
Characterization and Identification of Bacterial Isolates
Pure cultures of the isolates were identified on the basis of their cultural, morphological and physiological characteristics by reference to Holt et al. (1994) and Barrow and Feltham (1993). The tests performed include Gram-stain, spore-stain, motility test, catalase test, coagulase, urease production, nitrate reduction, methyl red, Voges-Proskauer, oxidative/fermentative test, gelatin hydrolysis, oxygen requirement, growth at different salt concentration and utilization of carbon-sources.
Physiological Studies of Microorganisms in the Hindgut of Amitermes evuncifer
Bacterial isolates were screened for their cellulolytic and amylolytic activities.
Determination of Cellulolytic Activities
The medium was Skinner cellulose medium B (Skinner, 1960), supplemented with either 1% (w/v) Whatman powdered cellulose or 1% carboxymethyl cellulose (CMC). The Skinner medium without cellulose served as control. Each of the isolates were streaked on plates containing carboxymethyl cellulose and incubated at 35°C for 48 h, while those containing cellulose powder were incubated at 27°C for 5 days. Culture plates were flooded with 1% Congo red, followed by 2M NaCl; and observed for zone of clearing by cellulose or carboxymethyl cellulose-degrading bacteria (Paul et al., 1993).
Determination of Amylolytic Activity
Alpha amylase production was tested by inoculating the bacterial culture on starch agar (Nutrient agar plus 1% soluble starch). Plates were incubated at 35°C for 3 days, after which they were flooded with iodine solution (0.3% I2 (w/v) in 3% KI (w/v)). Amylase activity was indicated by a clear/brown zone surrounding the colony.
Statistical Analysis
Analysis of variance of data obtained was carried out using the Statistical Package for Social Sciences (SPSS version 11.0).
RESULTS
The amount of total rainfall in the study area was about 1500 mm in September and October. During wet season (April to November), the relative humidity ranged between 85 and 90%, while it ranged between 50 and 55% during the dry season (December to March). The temperature during the wet season ranged from 23 and 26°C, while it ranged between 27 and 32°C during the dry season. The annual mean temperature was approximately 27°C.
The bacterial count in the hindgut of A. evuncifer is shown in Table 1. The total bacterial counts in soldier and worker termites were 1.46±0.26x106 and 2.51±0.31x106 cfu mL-1, respectively during the wet season. During the dry season, the bacterial counts were 5.30±1.1x104 and 5.80±0.9x104 cfu mL-1 in the soldier and worker termites respectively. The total microbial count during the wet season was significantly higher than the dry season (p≤0.05). However, there were no significant differences in the total bacterial counts in the gut of the worker and soldier termites in both seasons.
The characterization of bacterial isolates revealed that five different species of bacteria were isolated in the hindgut of A. evuncifer during the wet season. The bacteria were identified as Micrococcus luteus, Bacillus subtilis, B. cereus, Streptococcus sp. and Serratia marcescens. However, only Micrococcus luteus, Bacillus subtilis and Streptococcus sp. were isolated during the dry season (Table 1). The rate of occurrence of the bacterial isolates is shown in Table 2. Micrococcus had the highest occurrence of 100%, followed by B. subtilis which had 87.5% in both worker and soldier termites, respectively during the wet season. However the occurrence rate of B. subtilis during the dry season was 35 and 45% in the hindgut of soldier and worker termites, respectively. All the isolates in this study were amylolytic, while 73.7% of the Bacillus species was cellulolytic. None of the isolate showed cellulolytic activity on Whatman cellulose agar medium (Table 3).
| Table 1: | Total load (cfu mL-1) of bacteria in the hindgut of Amitermes evuncifer during the wet and dry seasons |
 | |
| Cfu = Colony forming unit | |
| Table 2: | Rate of occurrence of bacterial isolates |
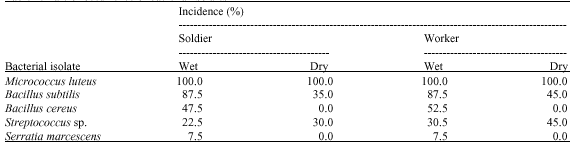 | |
| Table 3: | Enzymatic (amylase and cellulase) activities of bacteria isolated from the hindgut of A. evuncifer |
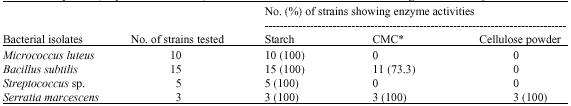 | |
| *CMC = Carboxymethyl cellulose | |
DISCUSSION
This study has shown that the hindgut of worker termites were more populated with bacteria than the hindgut of the soldier caste, both during the wet and the dry seasons. This observation is in agreement with the findings of Amund et al. (1986), who suggested that the high bacterial counts in workers could be attributed to the division of labour in the termite colonies. The workers are more active because they are responsible for the building of forage galleries nest and also take care of the young ones (Malaka, 1996). The high total bacterial population during the wet season may be due to the more pronounced activity of the termites during the wet season in which they build nest. The various environmental factors viz temperature, relative humidity and amount of rainfall characterizing the different season could be favorable to termite proliferation and the multiplication of the hindgut micro flora during the wet season. Four genera of bacteria were encountered in the hindgut of the wood-eating termite, A. evuncifer and they were identified to be species belonging to the genera Serratia, Micrococcus, Bacillus and Streptococcus. Some other researchers have reported the presence of these genera of bacteria in the gut of termites (Amund et al., 1986; Ohkuma and Kudo, 1996; Schäffer et al., 1996). Micrococcus luteus had the highest incidence rate in both workers and soldier castes.
The involvement of the bacterial flora of the termite gut in the degradative activities, particularly of cellulose, which forms the bulk of the termite diet, was investigated in this study. Bacillus sp. were found to constitute the major celluloytic bacteria in the hindgut of A. evuncifer The occurrence of M. luteus and other non-cellulolytic organism may indicate the preponderance of degradative bye-products of cellulolytic activities in the hindgut. Such degraded compounds could readily be utilized to enhance the proliferation of non-cellulolytic microbes in the hindgut. Lee et al. (2002) reported that in nature, cellulose utilization is carried out by multiple cellulolytic species coexisting with each other and with many non-cellulolytic species. While cellulolytic species compete for cellulose, both cellulolytic and non-cellulolytic species compete for the products of cellulose hydrolysis (Fondevilla and Dehority, 1996). All the strains of Bacillus species were cellulolytic on carboxymethyl cellulose agar medium; none of them hydrolyzed Whatman cellulose powder. This may be due to the difference in the structures of the two cellulose substrates. It has been reported that the hydrolysis of amorphous cellulose is much faster than the crystalline forms of cellulose (Beguin, 1990). However, Serratia marcescens hydrolysed both the crystalline and non-crystalline forms of cellulose. Cellulose degradation requires the presence of a complex of enzymes namely exo-β-1, 4 glucanase, endo-β-1, 4 glucanase (cellobiohydrolase) and β-D-glucoside glucohydrolase (β glucosidase) (Beguin and Aubert, 1994).The detection of amylolytic activity in all the bacterial isolates might be due to the fact that amylase is a constitutive enzyme in many microorganisms (Pandey et al., 2000).
Even though it has been well documented that the microflora in the hindgut of A. evuncifer, could degrade cellulose, further research is needed to determine the presence of endogenous cellulase gene in the hindgut of Amitermes evuncifer and also to investigate its relationship with the cellulase gene of the bacterial isolates in the hindgut of A. evuncifer.
REFERENCES
- Ako-Nai, K.A., O. Adejuyigbe, T.O. Adewumi and O.O. Lawal, 1992. Sources of intra-operative bacteria colonization of clean surgical wound and subsequent post-operative wound infection in a Nigerian hospital. East Afr. Med. J., 69: 500-507.
PubMed - Beguin, P., 1990. Molecular biology of cellulose degradation. Ann. Rev. Microbiol., 44: 219-248.
CrossRef - Beguin, P. and J.P. Aubert, 1994. The biological degradation of cellulose. FEMS Microbiol. Rev., 13: 25-58.
CrossRef - Fondevilla, M. and B.A. Dehority, 1996. Interaction between Fibrobacter succinogenes, Prevotella ruminicola and Ruminococcus flavefaciences in the digestion of cellulose from forages. J. Anim. Sci., 74: 678-684.
Direct Link - Holt, J.G., N.R. Kreig, P.H.A. Sneath, J.T. Staley and S.T. Williams, 1994. Bergey's Manual of Determinative Bacteriology. 9th Edn., Lippincott Williams and Wilkins, Baltimore, USA., ISBN-13: 9780683006032, Pages: 787.
Direct Link - Khademi, S., L.A. Guarino, H. Watanabe, G. Tokuda and E.F. Meyer, 2002. Structure of an endoglucanase from termite Nasutitermes takasagoensis. Acta Cryst., D58: 653-659.
Direct Link - Lynd, L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius, 2002. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev., 66: 506-577.
CrossRefPubMedDirect Link - Ohkuma, M. and T. Kudo, 1996. Phylogenetic diversity of the intestinal bacteria community in the termite Reticulitermes speratus. Applied Environ. Microbiol., 62: 461-468.
PubMed - Pandey, A., P. Nigam, C.R. Soccol, V.T. Soccol, D. Singh and R. Mohan, 2000. Advances in microbial amylases. Biotechnol. Applied Biochem., 31: 135-152.
PubMedDirect Link - Paul, J., S. Saxena and A. Varma, 1993. Ultrastructural studies of the termite Odontermes obesus gut microflora and its cellulolytic properties. World J. Microbiol. Biotechnol., 9: 108-112.
CrossRef - Todaka, N., S. Moriya, K. Saita, T. Hondo and I. Kiuchi et al., 2007. Environmental cDNA analysis of-the genes involved in lignocellulose digestion in the symbiotic protist community of Reticulitermes speratus. FEMS Microbiol. Ecol., 59: 592-599.
Direct Link - Schafer, A., R. Konrad, T. Kuhnigk, P. Kampfer, H. Hertel and H. Konig, 1996. Hemicellulose-degradating bacteria and yeasts from the termite gut. J. Applied Microbiol., 80: 471-478.
CrossRefDirect Link