
J.S. Sidhu
Department of Entomology, Punjab Agricultural University, Ludhiana, 141004, India
R.S. Mann
Department of Entomology and Nematology, Natural Area Drive, University of Florida, Gainesville, FL-32603, USA
N.S. Butter
Department of Entomology, Punjab Agricultural University, Ludhiana, 141004, India
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 1 | Page No.: 62-66







ABSTRACT
Cotton leaf curl virus (CLCuV) has a complex association with their whitefly vector. To further understand these relationships, longevity and fecundity of viruliferous and non-viruliferous Bemisia tabaci (Gennadius) was compared on three different ages of cotton plants (25, 45 and 55 days old). Cotton leaf curl virus infection reduced the fecundity and longevity of B. tabaci compared with non-viruliferous whiteflies. Both viruliferous and non-viruliferous whiteflies survived longer on older plants than on younger plants. Female whiteflies survived longer than the male whiteflies irrespective of their being viruliferous or non-viruliferous. Plant age did not affect the fecundity however egg viability declined with increasing age of plants.
PDF Abstract XML References Citation
How to cite this article
J.S. Sidhu, R.S. Mann and N.S. Butter, 2009. Deleterious Effects of Cotton leaf curl virus on Longevity and Fecundity of Whitefly, Bemisia tabaci (Gennadius). Journal of Entomology, 6: 62-66.
DOI: 10.3923/je.2009.62.66
URL: https://scialert.net/abstract/?doi=je.2009.62.66
DOI: 10.3923/je.2009.62.66
URL: https://scialert.net/abstract/?doi=je.2009.62.66
INTRODUCTION
Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) has become one of the most important pests of agricultural crops worldwide. In addition to feeding on more than 700 host plant species within 86 botanical families B. tabaci is a vector of numerous plant viruses including Cotton leaf curl virus (CLCuV) causing cotton leaf curl disease (CLCuD) (Ruiz et al., 2006; Akad et al., 2007; Qazi et al., 2007). This disease has become a major threat to cotton industry in Pakistan (Briddon and Markham, 2000) and later spread to India (Radhakrishnan et al., 2004; Briddon et al., 2006). The disease is believed to be caused by Cotton leaf curl virus (CLCuV), belonging to genus Begomovirus (Briddon and Markham, 2000; Mansoor et al., 2000; Varma and Malathi, 2003; Khan and Ahmad, 2005; Briddon et al., 2006; Iqbal et al., 2006; Qazi et al., 2007; Mann et al., 2008) of family Geminiviridae. The virus is transmitted by B. tabaci in a circulative, persistent manner and can be retained from a few days to the entire life period of the whitefly (Liu et al., 1998; Mann and Singh, 2004a).
Earlier studies suggest that viruses can have positive, neutral or negative effects on their vectors (Costa et al., 1993; Colvin et al., 2006; Jiu et al., 2007). These effects can be due to the direct effect of the viruses on their vectors (Pesic-van esbroeck et al., 1995; Rubinstein and Czosnek, 1997) or to the indirect effects of altered host plant morphology and nutritional quality (Belliure et al., 2005; Stout et al., 2006). Previous studies by Mann et al. (2008) demonstrated that B. tabaci reared on 20 and 40 days old Cotton leaf curl virus infected plants laid fewer eggs and had shorter longevity and total developmental period than reared on healthy plants of same age. However, in those studies, the whiteflies were reared on virus infected and healthy plants and hence could not distinguish if the noticed changes in whitefly biology were due to altered plant nutrition or to the direct effect of the virus on its vector. In the present studies, the fecundity and longevity of viruliferous and non-viruliferous B. tabaci were examined on CLCuV resistant cotton plants to investigate the direct effect of CLCuV and the plant age on fecundity and longevity of whiteflies.
MATERIALS AND METHODS
The present studies were carried out at Cotton Research and Development Laboratories, Punjab Agricultural University Ludhiana, India during 2005 and 2006. Whiteflies were obtained by collecting adults from untreated Desi cotton, Gossypium arboreum, (indigenous cotton) plants resistant to CLCuV (Nateshan et al., 1996; Briddon et al., 2001; Rahman et al., 2002). These adults were released on new cotton plants in 60 mesh, 1.2x1.2x1.2 m rearing cages for oviposition. The adults were removed and killed manually after 3 days to maintain eggs only. Newly emerged whitefly adults from these eggs were released on new healthy plants of CLCuV susceptible, G. hirsutum cultivar F846 for verification of virus infection. Adults from plants with no symptoms in 40 days were considered non-viruliferous. These adults were reared on CLCuV resistant plants of G. hirsutum hybrid LHH144 in separate rearing cages for further studies.
Whiteflies were prepared as viruliferous by giving them an acquisition access period of 24 h on virus source as required. For maintaining virus source cotton plants, exhibiting typical symptoms of CLCuD described by Singh et al. (1994) and Kang et al. (2003) were obtained from the field and transplanted in earthen pots containing a mixture of loamy soil and farm yard manure in a 1:1 ratio. New plants were also grown up and infected with CLCuV through whitefly mediated transmission. Whiteflies were given an acquisition access period of 24 h on virus source and were confined on new healthy plants for 8 h (inoculation access period). The new plants were grown up and maintained in 1.2x1.2x1.2 m cages and used as virus source after the appearance of typical CLCuD symptoms.
Effect of CLCuV on longevity and fecundity of whiteflies was studied on 25, 40 and 55 days old plants (DOP) of cotton. The experimental cotton plants, G. hirsutum, CV LHH144, were planted in earthen pots (30x23 cm) with 3 to 4 seeds per pot. The plants were raised and maintained in separate cages following the standard procedures for growing cotton crop. Twenty test plants were used for each treatment comprising three replications for each treatment. Ten newly emerged viruliferous and non-viruliferous whiteflies were separately confined on underside of cotton leaves with clip cages to record their longevity. Insect mortality was recorded daily till all the insects died. Insects were transferred to new leaves every week to avoid emergence of new adults on the same leaves. Experiments were conducted for males as well as females separately.
To study fecundity two pairs of newly emerged male and female viruliferous and non-viruliferous whiteflies were separately confined on underside of leaves of 25, 40 and 55 DOP with clip cages for egg laying. The females were serially transferred daily to new test plants after oviposition till all the insects died. The numbers of eggs laid and hatched under each clip cage were counted with the help of a binocular microscope. A two-factor ANOVA was conducted using whitefly infection and plant age as factors. When the factors or their interactions were significant at p<0.05, the means were separated with Turkey`s HSD using SAS, Version 9.1 (SAS, 2003).
RESULTS AND DISCUSSION
The non-viruliferous male and female whiteflies survived longer than the viruliferous male and female whiteflies (Table 1). Female whiteflies survived longer than the male whiteflies irrespective of their being viruliferous or non-viruliferous. The average viruliferous and non-viruliferous male longevity ranged from 5.35 to 12.88 days and 6.37 to 15.09 days, respectively. While the average viruliferous and non-viruliferous female longevity ranged from 8.83 to 15.51 days and 10.76 to 17.31 days, respectively. Both viruliferous and non-viruliferous whiteflies (male and female) survived longer on 55 DOP than 25 and 40 DOP. The viruliferous whiteflies laid fewer eggs than non-viruliferous whiteflies irrespective of the plant age (Table 2). However, eggs laid by viruliferous whiteflies had better viability than the eggs from non-viruliferous whiteflies. Overall whiteflies had better egg viability on younger plants than older plants, while plant age did not affect the fecundity. Non-viruliferous whiteflies laid more eggs on 55 DOP, while, viruliferous whiteflies laid similar number of eggs on 25, 40 and 55 DOP. However, both viruliferous and non-viruliferous whiteflies had better egg viability on 25 DOP than on 40 and 55 DOP.
| Table 1: | Mean longevity of viruliferous and non-viruliferous Bemisia tabaci adults on cotton plants (G. hirsutum) |
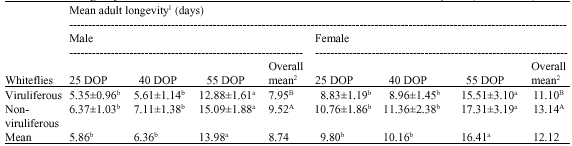 | |
| Values are shown in Mean±SE, 1Mean of 3 replications comprising 20 plants/replication, 2Means within a columns and rows followed by a different letter(s) differ significantly (p<0.05) | |
| Table 2: | Mean fecundity and egg viability of viruliferous and non-viruliferous Bemisia tabaci adults on cotton plants (G. hirsutum) |
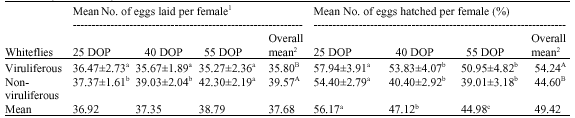 | |
| Values are shown in Mean±SE, 1Mean of 3 replications comprising 20 plants/replication, 2Means within a columns and rows followed by a different letter(s) differ significantly (p<0.05) | |
Earlier studies by Mann et al. (2008) showed that whiteflies laid fewer eggs on virus infected plants than healthy plants of same age and we speculated the results due to the altered plant nutrition or to the direct negative effect of the virus on its vector. Present studies showed that CLCuV have direct negative effects on B. tabaci to reduce its longevity and fecundity. Present results are similar with that of Rubinstein and Czosnek (1997).
The researchers demonstrated that the life expectancy of TYLCV infected whiteflies was reduced by more than 20% than that of non-viruliferous whiteflies and viruliferous whiteflies showed significantly lower fecundity with no effects on hatchability. Similar effects of viruses on B. tabaci were also reported by Jiu et al. (2007) on Tomato yellow leaf curl China virus. However, in the same study the researchers reported just the opposite results in Tobacco curly shoot virus infected whiteflies. Similarly, Pesic-Van Esbroeck et al. (1995) reported ultra-structural abnormalities in the reproductive, digestive and excretory systems of Squash leaf curl virus infected whiteflies. While, McKenzie (2002) demonstrated the opposite with higher fecundity and equivalent fertility in Tomato mottle virus infected whiteflies.
No effect of CLCuV on whitefly fertility can be because of its inability to transmit transovarially while, the negative effects may be related to cytological and metabolic changes in the vectors or to the progressive invasion of many tissues by the virus, in particular invasion of the reproductive system and cells, tissues and other organs (Pesic-Van Esbroeck et al., 1995; Rubinstein and Czosnek, 1997). However, further studies would be required to confirm such affects of CLCuV on B. tabaci as Cotton leaf curl virus has a complex etiology and vector interactions from above mentioned viruses (Khan and Ahmed, 2005; Qazi et al., 2007).
Present results supported the views of Jiu et al. (2007) that viruses and their vectors compete for the same host plants as a shared food source and do not exhibit the beneficial mutualism vector-virus relationships leading into an evolutionary dead end for the virus as well as vectors. Both male and female whiteflies had longer adult duration on 55 days old plants than on 25 and 40 days old plants. These results supported the earlier findings of Menn et al. (2008) of improved longevity on older plants than on younger plants. However, Mann and Singh (2004b) reported that whiteflies transmit CLCuV more efficiently on younger plants than old plants.
Improved or negative performance of vectors on the virus infected plants is often correlated with nutritional quality of virus plants. However, present findings suggest that the performance of insect vectors on virus infected plants is also influenced by the direct effects of the viruses on their vector along with the altered nutritional status of the host plants. Therefore, direct effects of the viruses on their vector should also be taken into account while studying plant-virus-vector interactions and host plant resistance for these circulative persistent viruses. Further, the antagonistic relationship between plant viruses and their vectors should further be exploited for management of whiteflies and viruses.
REFERENCES
- Akad, F., A. Eybihtz, D. Edelbaum, R., Gorovits, O. Darissa, N. Iraki and H. Czosnek, 2007. Making a friend from a foe: Expressing a GroEL gene from the whitefly Bemisia tabaci in the phloem of tomato plants confers resistance to tomato yellow leaf curl virus. Arch. Virol., 152: 1323-1339.
CrossRefPubMedDirect Link - Belliure, B., A. Janssen, P.C. Maris, D. Peters and M.W. Sabelis, 2005. Herbivore arthropods benefit from vectoring plant viruses. Ecol. Lett., 8: 70-79.
CrossRefDirect Link - Briddon, R.W., S. Mansoor, I.D. Bedford, M.S. Pinner and K. Saunders et al., 2001. Identification of DNA components required for induction of cotton leaf curl disease. Virology, 285: 234-243.
CrossRefPubMedDirect Link - Briddon, R., S.E. Bull and I.D. Bedford, 2006. Occurrence of Sweet potato leaf curl virus in Sicily. Plant Pathol., 55: 286-286.
CrossRefDirect Link - Briddon, R.W. and P.G. Markham, 2000. Cotton leaf curl virus disease. Virus Res., 71: 151-159.
CrossRefPubMedDirect Link - Colvin, J., C.A. Omongo, M.R. Govindappa, P.C. Stevenson and M.N. Maruthi, 2006. Host plant viral infection effects on arthropod-vector population growth, development and behavior: Management and epidemiological implications. Adv. Virus Res., 67: 419-452.
CrossRefPubMedDirect Link - Costa, H., J.K. Brown, S. Sivasupramaniam and J. Bird, 1993. Regional distribution, insecticide resistance and reciprocal crosses between the A and B biotypes of Bemisia tabaci. Int. J. Trop. Insect Sci., 14: 255-266.
CrossRef - Iqbal, N., M.Y. Ashraf, F. Javed, M. Ashraf and S. Hameed, 2006. Cotton leaf curl virus: Ionic status of leaves and symptom development. J. Integrat. Plant Biol., 48: 558-562.
CrossRefDirect Link - Jiu, M., X.P. Zhou, L. Tong, X.Yang, F.H. Wan and S.S. Liu, 2007. Vector-virus mutualism accelerates population increase of an invasive whitefly. PLoS One, 2: e182-e182.
CrossRefPubMedDirect Link - Khan, J.A. and J. Ahmad, 2005. Diagnosis, monitoring and transmission characteristics of Cotton leaf curl virus. Curr. Sci., 88: 1803-1809.
Direct Link - Liu, D.H., D.J. Robinson and B.D. Harrison, 1998. Nuclear location of the 16K non-structural protein of tobacco rattle virus. J. Gen. Virol., 72: 1811-1817.
CrossRefPubMedDirect Link - Mann, R.S., J.S. Sidhu, N.S. Butter, A.S. Sohi and P.S. Sekhon, 2008. Performance of Bemisia tabaci (Hemiptera: Aleyrodidae) on healthy and Cotton leaf curl virus infected cotton. Florida Entomol., 91: 249-255.
CrossRefDirect Link - Mansoor, S., R. Briddon, S.E. Bull, I.D. Bedford and A. Bashir et al., 2003. Cotton leaf curl disease is associated with multiple monopartite begomoviruses supported by single DNA β. Arch. Virol., 148: 1969-1986.
CrossRefPubMedDirect Link - McKenzie, C.L., 2002. Effect of Tomato mottle virus (ToMoV) on Bemisia tabaci biotype B (Homoptera: Aleyrodidae) oviposition and adult survivorship on healthy tomato. Florida Entomol., 85: 367-368.
CrossRefDirect Link - Nateshan, H., M., V. Muniyappa, M.M. Swanson and B.D. Harrison, 1996. Host range, vector relations and serological relationships of Cotton leaf curl virus from southern India. Ann. Applied Biol., 128: 233-244.
CrossRef - Qazi, J., I. Amin, S. Mansoor, M.J. Iqbal and R.W. Briddon, 2007. Contribution of the satellite encoded gene betaC1 to cotton leaf curl disease symptoms. Virus Res., 128: 135-139.
CrossRefPubMedDirect Link - Radhakrishnan, G., V.G. Malathi and A. Varma, 2004. Detection of DNA A and DNA β associated with cotton leaf curl and some other plant diseases caused by whitefly transmitted geminiviruses. Ind. Phytopathol., 57: 53-60.
Direct Link - Rahman, M., D. Hussain and Y. Zafar, 2002. Estimation of genetic diversity among elite cotton cultivars, genotypes by DNA fingerprinting technology. Crop Sci., 42: 2137-2144.
Direct Link - Rubinstein, G. and H. Czosnek, 1997. Long term association of Tomato yellow leaf curl virus with its whitefly vector Bemisia tabaci: Effect on the insect transmission capacity, longevity and fecundity. J. Gen. Virol., 78: 2683-2689.
PubMedDirect Link - Ruiz, L., D. Janssen, G. Martin, L.Velasco, E. Segundo and I. M. Cuadrado, 2006. Analysis of the temporal and spatial disease progress of Bemisia tabaci-transmitted Cucurbit yellow stunting disorder virus and Cucumber vein yellowing virus in cucumber. Plant Pathol., 55: 264-275.
CrossRef - Stout, M.J., J.S.Thaler and B.P.H.J. Thomma, 2006. Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Ann. Rev. Entomol., 51: 663-689.
CrossRefPubMedDirect Link - Varma, A. and V.G. Malathi, 2003. Emerging geminivirus problems: A serious threat to crop production. Ann. Applied Biol., 142: 145-164.
CrossRefDirect Link