
F.E. Nwilene
Africa Rice Center (WARDA), P.M.B. 5320, Ibadan Nigeria, West Africa
A.K. Traore
Africa Rice Center (WARDA), 01 B.P. 2551, Bouaké 01, Côte d’Ivoire, West Africa
A.N. Asidi
Africa Rice Center (WARDA), 01 B.P. 2551, Bouaké 01, Côte d’Ivoire, West Africa
Y. Sere
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin, West Africa
A. Onasanya
Africa Rice Center (WARDA), 01 B.P. 2031, Cotonou, Benin, West Africa
M.E. Abo
National Cereals Research Institute (NCRI), Badeggi, P.M.B. 8, Bida, Niger State, Nigeria, West Africa
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 4 | Page No.: 198-206







ABSTRACT
The study aimed to investigate the vectorial capacity of twelve insect species to transmit Rice Yellow Mottle Virus (RYMV) from diseased seedlings of a susceptible rice variety (Bouaké 189) and a perennial wild rice (Oryza longistaminata) to seven alternative host plants. Results indicated that Trichispa sericea, Chaetocnema pulla, Chnootriba similis, Conocephalus longipennis, Oxya hyla, Paratettix sp., Zonocerus variegatus, Euscyrtus sp., Cofana spectra, Cofana unimaculata, Locris rubra and Locris maculata were capable of transmitting RYMV from infected Bouaké 189 and Oryza longistaminata to alternative weed hosts Leersia hexandra, Imperata cylindrica, Digitaria horizontalis, Echinochloa colona, Echinocloa crus-pavonis, Eleusine indica and Brachiaria lata. Only Chaetocnema pulla, Trichispa sericea, Chnootriba similis, Oxya hyla, Zonocerus variegatus, Euscyrtus sp., Parattetix sp., Cofana spectra, Cofana unimaculata and Locris rubra played an important role in transmitting the disease from rice to O. longistaminata, Leersia hexandra and Imperata cylindrica. The present study confirmed the vectorial capacity of these vectors out of which eight were reported for the first time in West Africa.
PDF Abstract XML References Citation
How to cite this article
F.E. Nwilene, A.K. Traore, A.N. Asidi, Y. Sere, A. Onasanya and M.E. Abo, 2009. New Records of Insect Vectors of Rice Yellow Mottle Virus (RYMV) in Cote d`Ivoire, West Africa. Journal of Entomology, 6: 198-206.
DOI: 10.3923/je.2009.198.206
URL: https://scialert.net/abstract/?doi=je.2009.198.206
DOI: 10.3923/je.2009.198.206
URL: https://scialert.net/abstract/?doi=je.2009.198.206
INTRODUCTION
Rice Yellow Mottle Virus (RYMV) is one of the most devastating diseases of irrigated and lowland rice, occurring in the sahel, tropical humid and sub-humid ecosystems of West Africa. However, it does occur in mangrove and inland swamps in Guinea and in upland rice in Sierra Leone and Côte d’Ivoire (Awoderu, 1991). The disease is characterized by mottling and yellow symptoms of varying intensities depending on the genotype. The virus is environmentally stable, highly infectious and endemic to Africa and surrounding islands. The host range of RYMV is narrow and restricted to gramineous species mainly in the rice tribes Oryzeae and Eragrostidae. The pathogen belongs to the sobemovirus group and transmitted mechanically through rice plants injuries and by insect vectors (Abo et al., 1998). Virus transmission by insects with biting and chewing mouthparts is commonly explained by an effective dose of the virus on contaminated mouthparts (Abo et al., 1998). The regular occurrence of insects in rice fields in West Africa has prompted close examination of these species as possible vectors (Abo et al., 1998). Although, there has been considerable effort to identify resistant rice varieties and to develop an integrated management of RYMV for the region’s farmers, the poor understanding of virus variability, epidemiology and role of vectors has hindered progress in the control of the disease. Previous studies in East Africa and West Africa to understand the epidemiology of the disease concentrated on insects feeding on diseased rice plants where the insects collect the virus and then pass them on to the next healthy plants that they feed on (Bakker, 1970, 1971; Abo et al., 2000a). Since rice is not cultivated in the fields continuously for 12 months in a year, there is need to know where, the virus spends the off-season. In this case, the alternative host plants may have a role to play. Therefore, the present study was designed to assess the vectorial capacity of insects found within rice fields to transmit the disease from rice to alternative hosts. This is important because in the absence of rice in the field, the vectors fly and take refuge on alternative hosts around the bunds or in nearby fields (Abo et al., 2000a). A better understanding of RYMV epidemiology requires information on the insects that serve as vectors and alternative hosts that act as reservoirs. This could then provide a basis for the development of appropriate methods of control that would improve sustainable intensification of lowland rice-based systems as well as the resource-use efficiency in irrigated rice-based systems.
MATERIALS AND METHODS
Study Site
The study was carried out between year 2000 and 2001. The trial sites were located at Sakassou and Gagnoa in Côte d’Ivoire. Sakassou is at latitude and longitude of 5°06 W and 7°52 N. The areas experienced a bimodal rainfall pattern, mechanized upland cultivation and double cropped lowland rice with dam-irrigation. Gagnoa is located in the forest zone with a latitude and longitude of 6°00 W and 6°06 N. Bimodal rainfall, single and double cropped lowland rice with stream irrigation.
The Alternative Host
Eight gramineous weeds including a wild rice commonly found in rice fields in West Africa were selected for the trials (Table 1). Each of these hosts was selected because (1) they represent annual and perennial grasses found along the toposequence, (2) they are diverse enough to capture the spread of the disease to rice at the beginning of the cropping season and (3) the grasses occur naturally from the free-draining uplands, through hydromorphic areas, to the seasonally flooded lowlands which may maintain the build-up of insect populations.
Insect Species
Adult insects were collected from rice fields using a new trapping net cage method and maintained on Bouaké 189 seedlings in rearing net cages measuring (1.0x0.8x0.5 m) according to Abo et al. (2000b) with some modification. Insects were allowed to undergo a generation in rearing cages before being transferred to a second net cage (0.8 x0.6x0.4 m) containing diseased seedlings of RYMV susceptible rice variety, Bouaké 189.
Transmission Tests
The viral transmission test was according to Sere et al. (2008) with some modification. Two groups of insects (leaf biting and leaf sucking) found in and around rice fields were tested for their ability to transmit RYMV from rice to alternative hosts and from a wild host to other alternative host plants. The insect were allowed to feed on diseased Bouake 189 for 48 h, after a group of five adult insect were removed having acquired the virus and caged to feed on healthy seedlings of each host plant.
| Table 1: | Names, grass type, plant height and ecology of seven alternative host plants used in the screening trials |
 | |
| Johnson (1997) | |
A control with virus-free Bouaké 189 and Oryza longistaminata was also set up. The cage experiment was replicated three times. Three potted plants were used per cage and the seedlings were kept under irrigation. The caged adult insects were allowed to feed on healthy seedlings for 10 days. At the end of 10 days, the insects were removed from the cage and killed by exposure under freeze temperature for 30 min. The ability of the five adult insects to transmit RYMV to each host plant was checked serologically using enzyme-linked immunosorbent assay (ELISA) test (Clark and Adams, 1977). The leaf samples from the three replications were analyzed separately in ELISA to detect the virus. In order to relate the results of artificial inoculation tests to the natural sources of RYMV in wild insects and alternative hosts, a monthly survey was carried out in August, 2000 through July, 2001 at Sakassou and Gagnoa. The insects were collected using the trapping net cage method and the plant samples were also taken within the area of the net cage. Five samples were taken per field at every visit. All insects caught and plant samples were put in small plastic containers and plastic bags, respectively, for further laboratory ELISA test (Clark and Adams, 1977).
RESULTS AND DISCUSSION
In the artificially inoculated screen house cages, 12 insect species (Trichispa sericea, Chaetocnema pulla, Chnootriba similis, Conocephalus longipennis, Oxya hyla, Paratettix sp., Zonocerus variegatus, Cheilomenes lunata, Euscyrtus sp., Cofana spectra, Cofana unimaculata and Locris rubra) were found capable of transmitting RYMV from diseased Bouaké 189 seedlings to 8 alternative host plants (Oryza longistaminata, Leersia hexandra, Imperata cylindrica, Digitaria horizontalis, Echinochloa colona, Echinocloa crus-pavonis, Eleusine indica, and Brachiaria lata (Table 2). Insect species capable of transmitting RYMV from wild rice (diseased Oryza longistaminata) to other alternative host plants were also identified (Table 3). From wild hosts under natural conditions in Gagnoa, RYMV infection was detected in the following insect vectors: Chaetocnema pulla, Cheilomenes lunata, Conocephalus longipennis, Oxya hyla, Cofana spectra and Cofana unimaculata (Table 3-7). Out of these insect vectors, four are known vectors (Trichispa sericea, Chaetocnema pulla, Chnootriba similis and Conocephalus longipennis) (Bakker, 1971; Abo et al., 2000b). The remaining eight are new and reported for the first time in West Africa.
| Table 2: | Insect species capable of transmitting RYMV from diseased Bouaké 189 to alternative hosts after 48 h acquisition feeding period |
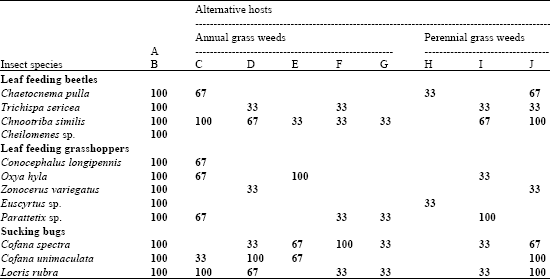 | |
| The symbols A: Control, B: Oryza longistaminata, C: Digitaria horizontalis, D: Eleusine indica, E: Brachiaria lata, F: Echinochloa colona, G: Echinochloa crus-pavonis, I: Leersia hexandra and J: Imperata cylindrica. Bold values are percent infected plants in ELISA test | |
| Table 3: | Insect species capable of transmitting RYMV from wild rice (diseased Oryza llongistaminata) to alternative hosts after 48 h acquisition feeding period |
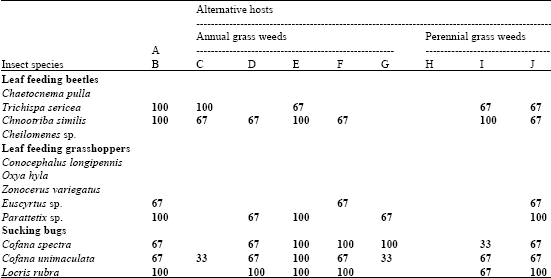 | |
| A: Control, B: Oryza longistaminata, C: Digitaria horizontalis, D: Eleusine indica, E: Brachiaria lata, F: Echinochloa colona, G: Echinochloa crus-pavonis, I: Leersia hexandra and J: Imperata cylindrica. Bold values are percent infected plants in ELISA test | |
| Table 4: | Natural sources of RYMV found in wild vectors and grass weeds at Gagnoa and Sakassou, Côte d’Ivoire |
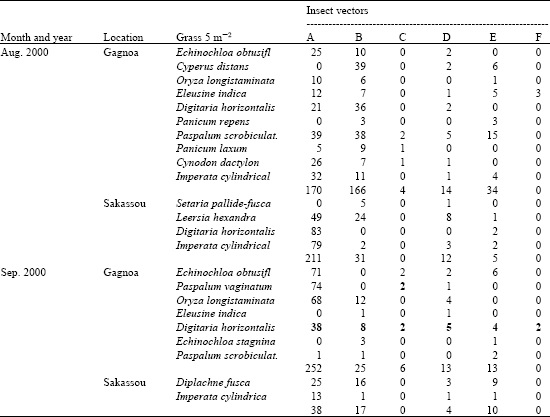 | |
| A: Chaetocnema pulla, B: Conocephalus longipennis, C: Oxya hyla, D: Cofana spectra, E: Cofana unimaculatus and F: Cheilomenes sp. Bold values are infected vectors and plants in ELISA test | |
Virus transmission by insects is a common way for viruses to travel between different host plants and this is possibly as a result of a protein that plant viruses attach to as they hitch an insect ride between plants (Uzest et al., 2007). The leaf feeding beetles (Chaetocnema pulla, Trichispa sericea and Chnootriba similis), the leaf feeding grasshoppers (Oxya hyla, Zonocerus variegatus, Euscyrtus sp. and Parattetix sp.) and the sucking bugs (Cofana spectra, Cofana unimaculata, Locris rubra) play important role in transmitting the disease from rice to perennial wild rice. These vectors carried the virus in the field. Perennial hosts with rhizomes could play an important role, because they act as reservoirs for the spread of the disease.
| Table 5: | Natural sources of RYMV found in wild vectors and grass weeds at Gagnoa and Sakassou, Côte d’Ivoire |
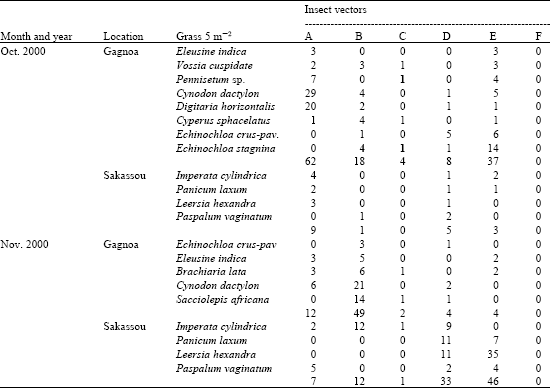 | |
| A: Chaetocnema pulla, B: Conocephalus longipennis, C: Oxya hyla, D: Cofana spectra, E: Cofana unimaculatus and F: Cheilomenes sp. Bold values are infected vectors and plants in ELISA test | |
| Table5: | Continued |
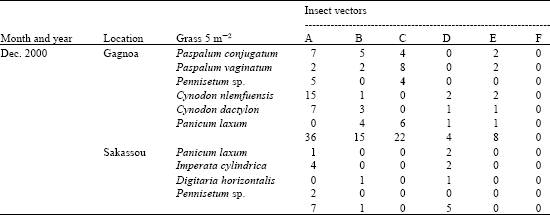 | |
| Table 6: | Natural sources of RYMV found in wild vectors and grass weeds at Gagnoa and Sakassou, Côte d’Ivoire |
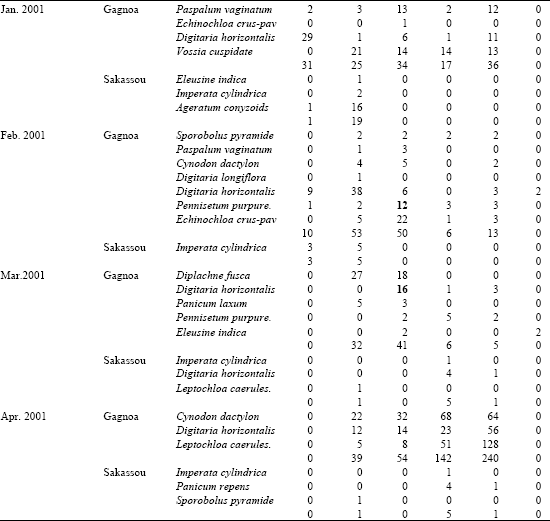 | |
| A: Chaetocnema pulla, B: Conocephalus longipennis, C: Oxya hyla, D: Cofana spectra, E: Cofana unimaculatus and F: Cheilomenes sp. Bold values are infected vectors and plants in ELISA test | |
| Table 7: | Natural sources of RYMV found in wild vectors and grass weeds at Gagnoa and Sakassou, Côte d’Ivoire |
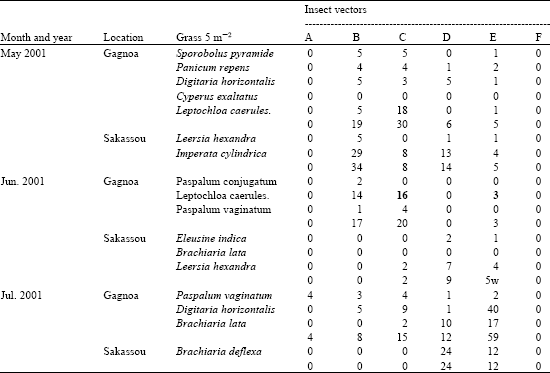 | |
| A: Chaetocnema pulla, B: Conocephalus longipennis, C: Oxya hyla, D: Cofana spectra, E: Cofana unimaculatus and F: Cheilomenes sp. Bold values are infected vectors and plants in ELISA test | |
However, the annual grass Digitaria horizontalis appears to be favoured over other grasses by the leaf-biting and leaf-sucking vectors. Furthermore, all the vectors transmitted RYMV to Bouaké 189 which was used as a control plant.
Variable virus transmission efficiency by vector species has been demonstrated. For instance, Frankliniella schultzei was more efficient in transmitting Tomato spotted wilt virus (TSWV) than Scirtotrips dorsalis (Rosello et al., 1996). Burrow et al. (2006) indicated that the difference between populations in their ability to transmit virus was demonstrated for the first time with Ciccadiluna mbila and the Maize streak virus. Gray et al. (2002) indicated that clonal populations of Schizaphis graminum, a vector of Barley yellow dwarf virus, could differ in their ability to transmit viruses. Although the potential of three RYMV insect vectors, Oxya hyla, Locris rubra and Chnootriba similes has been evaluated (Sere et al., 2008), little is known about the role of other insects found in farmers’ rice fields in the transmission of the virus.
Natural sources of RYMV were found in grasses belonging to the annual species Digitaria horizontalis, Echinochloa crus-pavonis and Leptochloa caerulescens at Gagnoa and perennial species Imperata cylindrica at Sakassou from August 2000 to July 2001. In September 2000, the grass Digitaria horizontalis and all the vectors sampled from it were RYMV positive by ELISA test. This provides strong evidence that wild hosts must be the main source of infection for rice crop.
CONCLUSION
Out of 12 insect vectors found to transmit RYMV, Chaetocnema pulla, Oxya hyla and Conocephalus longipennis are the most important because they are short feeding insects. Insects that have short feeding periods before moving to another plant are highly mobile and are considered to be the most efficient vectors of viruses. The implications of leafhoppers (Cofana spectra and Cofana unimaculata) and spittle bug (Locris rubra) in RYMV spread suggest that virus transmission goes beyond the boundaries of biting and chewing insects in West Africa. In earlier study, the stalk eyed flies, leafhoppers and spittlebugs were unable to transmit RYMV to susceptible rice host. This could be as a result of emergence of resistance breaking virus and presence of two serotypes of Rice yellow mottle virus reported in Côte d’Ivoire.
The fact that the virus found in alternative hosts under artificial inoculation corresponds to virus carriers in the wild indicates that sources of virus infection for the spread of the disease to rice crop are mainly from wild hosts. The abundance of grasses found in the forest (Gagnoa) and derived savanna (Sakassou) zones reflects the diversity of the natural flora in these locations. This diversity, coupled with their overlapping maturity periods, may help maintain high population densities of RYMV vectors in these zones where the infection reservoir of the virus gradually increases under conditions of year-round cropping with irrigation.
ACKNOWLEDGMENTS
We express our appreciation to Dr. Sidi Sanyang and Mr. Guy Manners for their useful suggestions and reviews of the manuscript. The technical assistance of Mr. Lekpa Jean Baptiste and Mr. Etienne Dessieh is gratefully acknowledged.
REFERENCES
- Abo, M.E., A.A. Sy and M.D. Alegbejo, 1998. Rice Yellow Mottle Virus (RYMV) in Africa: Evolution, distribution, economic significance on sustainable rice production and management strategies. J. Sustainable Agric., 11: 85-111.
Direct Link - Abo, M.E., M.D. Alegbejo, A.A. Sy and S.M. Misari, 2000. An overview of the mode of transmission, host plants and methods of detection of Rice yellow mottle virus. J. Sustain. Agric., 17: 19-36.
CrossRefDirect Link - Rosello, S., M.J. Diez and F. Nuez, 1996. Viral diseases causing the greatest economic losses to the tomato crop. I. The Tomato spotted wilt virus: A review. Scientia Hortic., 67: 117-150.
CrossRef - Awoderu, V.A., 1991. The rice yellow mottle virus situation in West Africa. J. Basic Microbiol., 31: 91-99.
CrossRefDirect Link - Bakker, W., 1970. Rice yellow mottle, a mechanically transmissible virus disease of rice in Kenya. Neth. J. Plant Pathol., 76: 53-63.
CrossRefDirect Link - Bakker, W., 1971. Three new beetle vectors of rice yellow mottle virus in Kenya. Neth. J. Plant Pathol., 77: 201-206.
CrossRefDirect Link - Burrow, M.E., M.C. Caillaud, D.M. Smith, E.M. Benson, F.E. Gildow and S.M. Gray, 2006. Genetic regulation of poliovirus and luteovirus transmission in the aphid Scizaphis graminum. Phytopathology, 96: 828-837.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Gray, S.M., D. Smith, L. Barbierri and J. Burd, 2002. Virus transmission phenotype is correlated with host adaptation among genetically diverse populations of the Aphid Scizaphis graminum. Phytopathology, 92: 970-975.
CrossRefDirect Link - Johnson, D.E., 1997. Weeds of Rice in West Africa. West Africa Rice Development Association, Cote D'ivoire.
Direct Link - Sere, Y., A. Onasanya, F.E. Nwilene, M.E. Abo and K. Akator, 2008. Potential of insect vector screening method for development of durable resistant cultivars to rice yellow mottle virus disease. Int. J. Virol., 4: 41-47.
CrossRefDirect Link - Uzest, M., D. Gargani, M. Drucker, E. Hebrard and E. Garzo et al., 2007. A protein key to plant virus transmission at the tip of insect vector stylet. Proc. Natl. Acad. Sci. USA., 104: 17959-17964.
PubMedDirect Link