
M.W. Mwatawala
Department of Crop Science and Production, Sokoine University of Agriculture, Morogoro, Tanzania
M. De Meyer
Entomology Section, Royal Museum for Central Africa, Tervuren, Belgium
R.H. Makundi
Pest Management Centre, Sokoine University of Agriculture, Morogoro, Tanzania
A.P. Maerere
Department of Crop Science and Production, Sokoine University of Agriculture, Morogoro, Tanzania
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 1 | Page No.: 18-27







ABSTRACT
The dominancy of introduced Bactrocera species (Diptera: Tephritidae) over the native fruit fly species was assessed based on data collected from a trapping and sampling program in Morogoro, Tanzania, from 2004 to 2006. Two invasions by Bactrocera species namely the invasive fruit fly Bactrocera invadens Drew, Tsuruta and White and the Solanum fruit fly Bactrocera latifrons (Hendel) have been recorded in 2003 and 2006, respectively. These add to an earlier introduced melon fly Bactrocera cucurbitae (Coquillett). Points and exact times of entry of these species are still unknown. Dominance of Bactrocera species over the native Ceratitis species has been speculated from other parts of the world. Results of this study also suggest the dominancy of Bactrocera species over native Ceratitis species in Tanzania. B. cucurbitae seems to dominate the other cucurbit infesters in terms of abundance and infestation rate. Similarly, B. invadens seems to dominate the native Ceratitis species in orchard fruits in terms of abundance, host range and infestation rate. B. latifrons, whose distribution in the country is still unclear, seems to be the dominant species in its main hosts from family Solananaceae. The outcome of the competition resulting from these introductions is speculated upon. Presence of these pests calls for strong surveillance systems and quarantine regulations to protect the infant fruit industry of Tanzania.
PDF Abstract XML References Citation
How to cite this article
M.W. Mwatawala, M. De Meyer, R.H. Makundi and A.P. Maerere, 2009. An Overview of Bactrocera (Diptera: Tephritidae) Invasions
and Their Speculated Dominancy over Native Fruit Fly Species in Tanzania. Journal of Entomology, 6: 18-27.
DOI: 10.3923/je.2009.18.27
URL: https://scialert.net/abstract/?doi=je.2009.18.27
DOI: 10.3923/je.2009.18.27
URL: https://scialert.net/abstract/?doi=je.2009.18.27
INTRODUCTION
Fruit flies (Diptera: Tephritidae) occur almost all over the world but the major pest genera have limited natural distribution (White and Elson-Harris, 1992). However, various Tephritid species have been accidentally or intentionally introduced into areas beyond their natural range. Examples include Ceratitis capitata (Widemann), which is native to tropical Africa, but is now one of the most widely distributed fruit flies because of introductions through human activities. Bactrocera cucurbitae, which is probably native to the Oriental Region, has been introduced into East Africa, Mauritius, the Ryukyu Islands of Japan, New Guinea and the nearby islands, Guam and Hawaii (Munro, 1984; Hooper and Drew 1989; Kakinohana, 1994).
Recently two Bactrocera introductions have been reported from Tanzania. First, B. invadens which was detected in 2003 (Mwatawala et al., 2004) and secondly, B. latifrons which was detected in 2006 (Mwatawala et al., 2007). Such introductions are an increasing threat, due to a worldwide increase of both commodity shipments (including fresh fruits from other continents) and the booming intercontinental tourist industry. A review (Duyck et al., 2004) has shown that, often, the presence of a newly introduced invasive polyphagous species results in a decrease in number and niche shift of the pre-established species. In most cases reviewed by Duyck et al. (2004), a Bactrocera species has invaded and numerically dominated a native Ceratitis species, thus assuming the key pest status. Integrated Pest Management (IPM) focuses on key pest(s) in an ecosystem, because by virtue of their numerical strength, they inflict heavy damages on crops. Understanding competition and dominancy in farming systems will help in developing a more focused IPM program for fruit flies. Studies in Réunion have shown the displacement of native species by Bactrocera zonata (Saunders) (Duyck et al., 2004, 2006b, 2008). The competitive displacement of C. cosyra by B. invadens has been suggested by Vayssières et al. (2005), Mwatawala et al. (2006b) and Ekesi et al. (2006). There are concerns about lack of data on pest status of B. cucurbitae in Africa despite its longtime presence. However, Vayssières et al. (2008) have shown that in reunion, B. cucurbitae has demonstrated more advantageous demographic characters related to competition, compared to D. ciliatus Loew, another major cucurbit infester on the island. Similarly, data on the recently introduced B. latifrons are still sketchy. This study examines the effect of the numerical dominancy of Bactrocera species over native Ceratitis species.
MATERIALS AND METHODS
Studies were conducted between October 2004 and October 2006, in Morogoro region. Four sites representing the three agro-ecological zones of Morogoro including, Sokoine University of Agriculture (SUA) horticulture unit, Mikese, Mkindo and Nyandira, were selected. Descriptions of the study sites were given by Mwatawala et al. (2006a). Studies involved collecting fruits to determine emerging species and trapping of the adult flies. Additional sampling of fruits was done from August through December 2007 and this focussed mainly on cucurbitaceous and solanaceous fruits which were less collected during the 2004-2006 period.
The fruits were collected each week following the procedure described by Copeland et al. (2002). The rearing the flies followed the procedures used by Copeland et al. (2002). Specimens were preserved following methods described by White and Elson-Harris (1992). The identification of flies were done using standard keys presented by White and Elson-Harris (1992), CABI key (CABI, 2005) and White (2006). The data collected include fruit fly species, fruits they attack, geographical location and their reservoir host.
A split-plot design was used to compare the infestation rates of B. invadens, C. rosa and C. cosyra in the different fruit varieties during the peak fruiting season at SUA Horticultural Unit. The sources of variations were the fruit fly species (sub-plots) and the fruit varieties (main-plots). The mango varieties studied were kent, dodo tommy Atkins and keitt. Orange varieties included hamlin, jaffa, matombo sweet, valencia late and cassa grande. For guava two varieties, pink fleshed and white fleshed were studied. Independent experiments were set for mango, citrus and guava. Each experiment was replicated for two fruiting seasons except mango, whereby sampling was done for one season due to insufficient mango fruits obtained from Horticulture unit in 2006/2007 season. In this case, weekly samples formed replicates. Two trees of each variety were randomly selected and marked for sampling. The sample size consisted of 5-10 fruits of each variety depending on the availability of fruits.
Relative Abundance Index (RAI) was determined as described by Segura et al. (2006). In this case RAI has been defined as the abundance of B. invadens relative to the combined abundance of B. invadens and one of the Ceratitis species (C. rosa, C. cosyra or C. capitata). In determining RAI, fruits that are hosts of B. invadens as well as one or more of Ceratitis species were included.
Trapping was carried out for two years starting October 2004 at the SUA Horticulture unit. For B. invadens vs. Ceratitis species, only Protein Bait (PB) was used, while for B. cucurbitae versus the pre-established cucurbit infesters, PB and Cue Lure (CL) were used. The data were collected once a week and catches of flies were calculated as the number of flies/trap/week. The protein bait was replaced each week while other attractants were replaced after four weeks.
Mean weekly catches of B. cucurbitae, Dacus ciliatus, D. punctatifrons and D. bivittatus by each attractant (PB and CL) were compared in a split plot design experiment at the SUA Horticulture Unit from January 2005 to January 2006. This one-year cycle was first divided into specific periods of observation of four weeks, coinciding with the time of changing the attractants. A total of thirteen periods of observation were scheduled. The weekly catches of each of the species in a respective lure during each period of observation were first pooled and the average number of flies per trap per week was calculated. There were three sources of variation namely, periods of observation (main-plot), orchards (sub-plots) and fruit fly species (sub-sub-plots). Each treatment was replicated twice. The Analysis of Variance (ANOVA) followed by means separation using the Least Significant Difference (LSD) were used to compare fruit fly catches. The data were analyzed using SAS version 9 (SAS institute Inc., USA).
RESULTS
B. invadens Versus the Native Infesters of Orchard Fruits
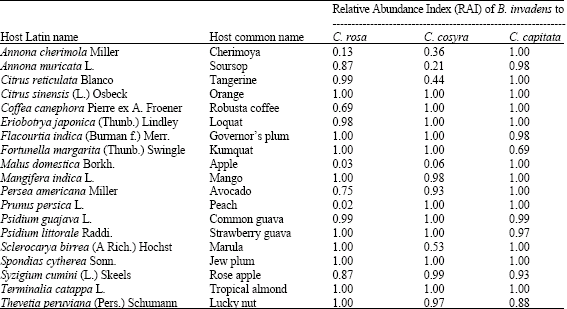
The dominancy of B. invadens over the native Ceratitis species is assessed in terms of Relative Abundance Index (RAI), infestation rates and seasonal abundance. Table 1 presents the relative abundance of B. invadens to the three Ceratitis species in 19 hosts. The fruit species included those attacked by B. invadens as well as by one or more of the Ceratitis species. Relative abundance of B. invadens to C. cosyra was high (more than 0.5) in all fruit species except tangerine, soursop and cherimoya, the last two being members of Annonanceae family. RAI of B. invadens was higher even in mango, a traditional host of C. cosyra. When comparing B. invadens to C. rosa, RAI was high in all fruit species except cherimoya, apple and peach, which are grown in high altitude areas, whose climate is favourable for C. rosa. When compared to C. capitata the relative abundance of B. invadens was high in all the fruit species (more than 0.5) with the lowest recorded in kumquat.
The infestation rates of the three fruit fly species in three commercial fruits were compared and results are shown in Table 2. Significant differences in infestation rates of the three fruit fly species in each of the three fruit species were observed. In all the fruit species, the infestation rate of B. invadens were highest compared to those of Ceratitis species. The incidences of each fruit fly species, in the varieties of each the studied fruits, were not significantly different.
| Table 1: | Relative abundance of B. invadens in fruit species also infested by one or more of the major Ceratitis species |
 | |
| Table 2: | Infestation rate of the major fruit flies in major fruit species |
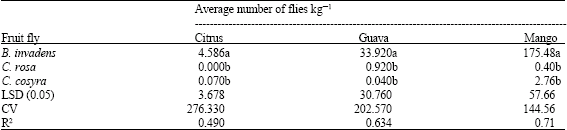 | |
| Means in a column followed by the same letter(s) are not significantly different (ANOVA and LSD) | |
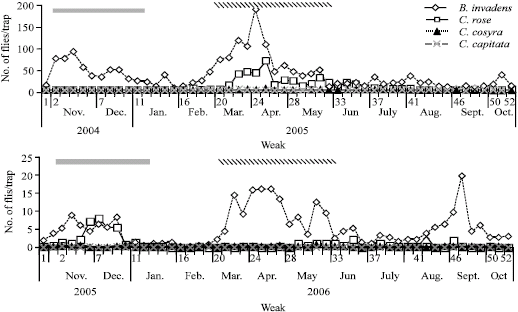 | |
| Fig. 1: | Seasonality of B. invadens and Ceratitis species at SUA Horticulture unit as recorded in PB (Filled bar indicates the short rainy season, stripped bar indicates long rainy season) |
Figure 1 presents the seasonal abundance of the three fruit fly species as determined by PB. The bait was chosen in the analysis because of its non-specificity to species and sex. In both years, the catches of B. invadens were high compared to the Ceratitis species. Catches of the latter were sporadic and in minimal. It should be noted that high populations were recorded during the rainy seasons when many fruits are available. It can be generally concluded that B. invadens numerically dominates the native Ceratitis species in terms of RAI, infestation rates and abundance.
B. cucurbitae Versus the Native Cucurbit Infesters
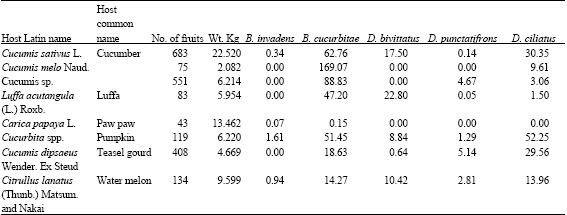
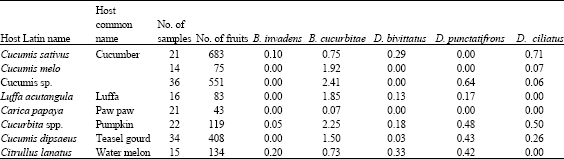
The dominancy of B. cucurbitae over the native cucurbit infesters is assessed in terms of infestation rates and seasonal abundance. Results of sampling of the major cucurbitaceous fruits (pumpkin, cucumber and water melon) are shown in Table 3 and 4. The infestation rate of B. cucurbitae were highest in all the sampled species, except pumpkin and teasel gourd. In pumpkin, the infestation rate was slightly lower that of Dacus ciliatus and a general conclusion cannot be made.
Mean weekly catches of the four cucurbit infesters by CL and PB were determined and results are presented in Table 5. Significant differences in mean weekly catches of the cucurbit infesters were observed. The catch of B. cucurbitae in CL was highest and significantly different from all Dacus species. This was followed by D. ciliatus while D. bivittatus was least caught. However the catches of the three Dacus species were not significantly different. Similarly in PB, the highest mean weekly catch was that of B. cucurbitae and was followed by D. ciliatus and D. punctatifrons. The catch of B. cucurbitae was significantly different from those of Dacus species, whose catches were not significantly different from each others. In both the parapheromone and the food bait, the mean weekly catch of B. cucurbitae was highest compared to the Dacus species.
| Table 3: | Infestation rates of cucurbit infesters in Cucurbitaceous fruits |
 | |
| Table 4: | Incidence of cucurbit infesters in Cucurbitaceous fruits |
 | |
| Table 5: | Mean weekly catches of cucurbit infesters |
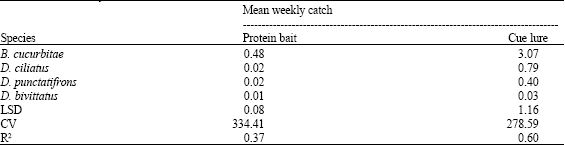 | |
| Means in a column followed by the same letters(s) ore not significantly different (ANOVA and LSD) | |
Figure 2 present the seasonal abundance of the cucurbit infesters determined by CL and PB during the two years trapping program. In CL, B. cucurbitae was recorded in highest numbers and less sporadically compared to Dacus species whose catches were minimal. In PB, catches of all cucurbit infesters were generally low, although B. cucurbitae appeared in substantial numbers. The results largely suggest numerical dominancy of B. cucurbitae over pre-established cucurbit infesters.
B. latifrons in Solanaceous Fruits
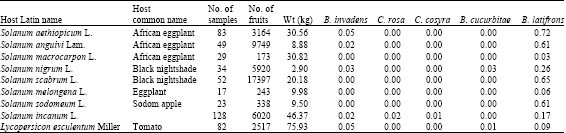
B. latifrons was detected for the first time in May 2006 in PB trap hung on a citrus tree. Latilure and cade oil, the known attractant for B. latifrons (McQuate and Peck, 2001), were not used up to the time of detection. The population of B. latifrons was mainly determined by sampling solanaceous fruits, mainly eggplant, Solanum aethiopicum, which seems to be its preferred host in this region. Other solanaceous fruits sampled include Black nightshade, Solanum nigrum and Solanum anguivi, as well as wild solanaceous fruits including Solanum incanum. The sampling was mainly conducted after the two years sampling programme. Infestation rates and incidence of fruit flies emerging from these fruits are presented in Table 6 and 7. In most cases, B. latifrons was the dominant species (with exception in tomato).
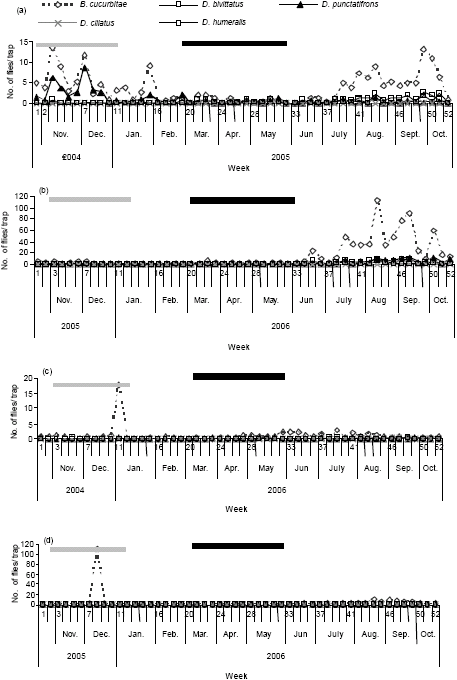 | |
| Fig. 2: | Seasonality of cucurbit infesters at SUA Horticulture unit as recorded by (a and b) CL (c and d) PB. (Filled bar indicates the short rainy season, stripped bar indicates long rainy season) |
| Table 6: | Incidence of fruit flies attacking solanaceous fruits |
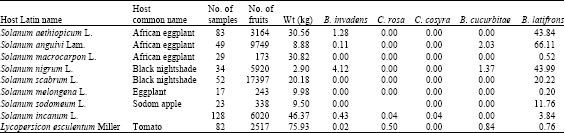 | |
| Table 7: | Incidence of fruit flies attacking solanaceous fruits |
 | |
DISCUSSION
Dominancy of Introduced Bactrocera Over Pre-Established Species
Results strongly suggest that the introduced Bactrocera spp. numerically dominate the native species in their niche. B. invadens dominates the Ceratitis species while B. cucurbitae dominates the native Dacus species. So far B. latifrons is the main infester of solanaceous fruits, but continued monitoring of this species is necessary. Where polyphagous tephritid species have been introduced into an area already occupied by other polyphagous tephritids, interspecific competition has resulted into a decrease in number and niche shift of pre-established species (Duyck et al., 2004). However, it should be noted that in most cases only the successful invasions are recorded since the less competitive introduced species fail to establish themselves and may become extinct before they are recorded. Most cases of tephritid invasions as described by Duyck et al. (2004), species of genus Bactrocera, invaded in the presence of and ultimately dominated numerically one or more species of the genus Ceratitis and the reverse was not observed. According to Duyck et al. (2004), invasive B. dorsalis has dominated the established C. capitata on at least two independent occasions while the reverse was not observed.
Invaders are generally assumed to be r-strategists and this means that during the colonization phase invaders are at an advantage but they have to compete at a later stage in order to establish a large stable population. In this regard, exotic invaders tend to be more competitive (Byers, 2000; Petren and Case, 1996) and they are able to quickly dominate the indigenous species. Co-existence between species can be promoted by competition-colonisation trade-offs among different species (Tillman, 1994), i.e., the bad competitors must be good colonizers because their maintenance depends on their being first to colonise empty spaces. It seems that B. invadens has been able to override the colonization- competition trade off. The whole genus of Bactrocera has a more K-oriented profile than Ceratitis and B. invadens has been able to display r- selected traits during the colonization phase and then later it has successfully competed with and probably excluded the pre-established species from their original niches. In this case, the proposition that a poor competitor is a good colonizer (or vice versa) could not hold. Duyck et al. (2007) reported that key traits for invasions are those that favour competition than colonization.
It can be proposed that competitors like Bactrocera spp. could exploit resources better than the pre-established spp., probably by denying them access to food or target sites. Interference competition implies that a more aggressive species gains access to resource to the detrimental effects on others. The larger body size of B. invadens (which is a K-selected trait) may be an advantage in exploitative as well as interference competition. K-selected species (such as Bactrocera spp.) can invade over r-selected species areas, taking over the most productive niche, while r-selected species (such as Ceratitis spp.) become restricted to a limited set of habitats (Duyck et al., 2004).
The B. invadens scenario in Tanzania and the whole of East Africa could be similar to the situation in Réunion islands where the invader B. zonata, was the best competitor and tended to occupy fruits and lay on them for more time than the Ceratitis species (Duyck et al., 2006a). The large body size of B. zonata may be an advantage in exploitative as well as in interference competition. The fact that B. invadens has a larger body size, wing length ranging from 5.4 to 6.9 mm (Drew et al., 2005) than C. rosa whose wing length ranges from 4.5 to 5.75 mm) (De Meyer, 1998) and C. cosyra whose wing length ranges from 3.4 to 5.2 mm (De Meyer and Freidberg, 2006). This suggests that a relatively K-like strategy may underlie the apparent directionality of interactions between the genus Bactrocera and Ceratitis, although further confirmations are needed.
Competitive Displacement
According to Duyck et al. (2004) data on tephritid invasions seem to support a hierarchical mode of competition (one species always dominates and excludes the other), although complete exclusion does not usually occur. Competitive displacement of C. cosyra by B. invadens has been suggested, although this is difficult to confirm due to lack of previous data on its abundance. As for B. latifrons, a concrete conclusion cannot be drawn since the species was more recently detected. There are several reported cases of competitive displacement in Tephritids. Ceratitis capitata was introduced to Australia from Europe around 1897 (Vera et al., 2002) and the species was gradually displaced around the Sydney area by the Queensland fruit fly Bactrocera tryoni (Froggatt), which invaded Australia from the north in the early 20th century (DeBach, 1966). The Oriental fruit fly largely displaced C. capitata from the coastal zones in Hawaii in 1945. C. capitata (which also invaded Hawaii in 1910) has now been restricted to the cooler climates at high altitudes where B. dorsalis is not found (Duyck et al., 2004). C. capitata became established in Réunion in 1939 and Mauritius in 1942, where the Mascarene fruit fly Ceratitis catoirii Guèrin-Mèneville was indigenous. A further invasion by C. rosa was also witnessed in Mauritius in 1953 and Reunion in 1955 (White et al., 2000). A similar inference can be made between B. invadens and C. cosyra, the latter seem to be confined to Annonanceae hosts. Along the same line, it seems C. rosa is more dominant in fruits grown in high altitude areas, where B. invadens occur in low numbers.
The peach fruit fly, Bactrocera zonata (Saunders), was found in Mauritius in 1987 and Réunion in 1991. C. rosa is dominant in high altitudes in Réunion while B. zonata continues to spread and has already colonized large portions of the niches used by other species at low altitudes.
CONCLUSION
Introduced Bactrocera species have shown numerical dominancy over the pre-established species sharing the same niche, as assessed by infestation rates in fruits as well as by abundance as determined by catches in traps. These species are now the key pests and should be the main target of any formulated IPM program. The results also suggest but could not confirm the competitive displacement of pre-established species by the introduced B. invadens. This is due to lack of earlier data on status of fruit flies in the study area. However, results strongly suggest the enforcement of quarantine and surveillance in order to defend the country from such devastating invasions.
REFERENCES
- Byers, J.E., 2000. Competition between two estuarine snails: Implication for invasions of exotic species. Ecology, 81: 1225-1239.
Direct Link - Copeland, R.S., R.A. Wharton, Q. Luke and M. De Meyer, 2002. Indigenous hosts of Ceratitis capitata (Diptera: Tephritidae) in Kenya. Annals Entomol. Soc. Am., 95: 672-694.
Direct Link - DeBach, P., 1966. Competitive displacement and coexistence principles. Annual Rev. Entomol., 11: 183-212.
Direct Link - Drew, R.A.I., K. Tsuruta and I.M. White, 2005. A new species of pest fruit fly (Diptera: Tephritidae: Dacinae) from Sri Lanka and Africa. Afr. Entomol., 13: 149-154.
Direct Link - Duyck, P.F., P. David and S. Quilici, 2004. A review of relationships between interspecific competition and invasions in fruit flies (Diptera: Tephritidae). Ecol. Entomol., 29: 511-520.
Direct Link - Duyck, P., P. David and S. Qulici, 2006. Climatic niche partitioning following successive invasions by fruit flies in La Réunion. J. Anim. Ecol., 75: 518-526.
Direct Link - Duyck, P., P. David, G. Junod, C. Brunel, R. Dupont and S. Qulici, 2006. Importance of competition mechanisms in successive invasions by polyphagous tephritids in La Réunion. Ecology, 87: 1770-1780.
CrossRefDirect Link - Duyck, P., P. David and S. Qulici, 2007. Can more selected species be better invaders? A case of fruit flies in La Réunion. Diversity Distributions, 13: 535-543.
Direct Link - Duyck, P., P. David, S. Pavoine and S. Quilici, 2008. Can host range allow niche differentiation of invasive polyphagous fruit flies (Diptera: Tephritidae) in La Réunion. Ecol. Entomol., 33: 439-452.
Direct Link - De Meyer, M., 1998. Revision of the subgenus Ceratitis (Ceratalaspis) Hancock (Diptera: Tephritidae). Bull. Entomol. Res., 88: 257-290.
Direct Link - Ekesi, S., P.W. Nderitu and I. Rwomushana, 2006. Field infestation, Life history and demographic parameters of the fruit fly Bactrocera invadens (Diptera: Tephritidae) in Africa. Bull. Entomol. Res., 96: 379-386.
CrossRefDirect Link - McQuate, G.T. and S.L. Peck, 2001. Enhacement of attraction of alpha-ionol to male Bactrocera latifrons (Diptera: Tephritidae) by addition of synergist, cade oil. Ecol. Behav., 94: 39-46.
CrossRef - Mwatawala, M.W., M. De Meyer, R.H. Makundi and A.P. Maerere, 2006. Biodiversity of fruit flies (Diptera, Tephritidae) at orchards in different agro-ecological zones of the Morogoro region, Tanzania. Fruits, 61: 321-332.
Direct Link - Mwatawala, M.W., M. De Meyer, R.H. Makundi and A.P. Maerere, 2006. Seasonality and host utilization of the invasive Bactrocera invadens (Diptera: Tephritidae) in Central Tanzania. J. Applied Entomol., 130: 530-537.
Direct Link - Mwatawala, M.W., M. De Meyer, R.H. Makundi and A.P. Maerere, 2007. Detection of the Solanum fruit fly, Bactrocera latifrons (Hendel) in Tanzania (Dipt., Tephritidae). J. Applied Entomol., 131: 501-503.
Direct Link - Mwatawala, M.W., I.M. White, A.P. Maerere, F.J. Senkondo and M. De Meyer, 2004. A new invasive Bactrocera species (Diptera: Tephritidae) in Tanzania. Afr. Entomol., 12: 154-156.
Direct Link - Petren, K. and T.J. Case, 1996. An experimental demonstration of exploitation ompetition in an ongoing invasion. Ecology, 77: 118-132.
Direct Link - Segura, D.F., V. Teresa, M., Cagnoti, C.L. Vacaro and N.O. De Coll et al., 2006. Relative abundance of Ceratitis capitata and Anastrepha fraterculus (Diptera: Tephritidae) in diverse host species and localities in Argentina. Ecology Popul. Bio., 99: 70-83.
CrossRefDirect Link - Tillman, D., 1994. Competition and biodiversity in spatially structured habitats. Ecology, 71: 2-16.
Direct Link - Vayssières, J., G. Goergen, O. Lokossou, P. Dossa and C. Akponson, 2005. A new Bactrocera species in Benin among mango fruit fly (Diptera: Tephritidae) species. Fruits, 60: 371-377.
Direct Link - Vayssières, J.F., Y. Carel, M. Coubes and P. Duck, 2008. Development of immature stages and comparative demographic of two cucurbit-attacking fruit flies in Reunion islands: Bactrocera cucurbitae and Dacus ciliatus (Diptera: Tephritidae). Physiol. Ecol., 37: 301-314.
Direct Link - Vera, M.T., R. Rodriguez, D.F. Segura, J.L. Cladera and R.W. Sutherst, 2002. Potential distribution of the mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae), with emphasis on Argentina and Australia. Environ. Entomol., 31: 1009-1022.
CrossRef - White, I.M., M. De Meyer and J.M. Stonehouse, 2000. A review of native and introduced fruit flies (Diptera: Tephritidae) in the Indian Ocean islands of Mauritius, Réunion, Rodriguez and Seychelles. Proceedings of the Indian Ocean Commission Regional Fruit Fly Symposium, June 5-9, 2000, Flic en Flac, Mauritius, pp: 15-21.