
M.S. Shathele
Department of Microbiology, King Faisal University, P.O. Box 1757 Al-Ahsa 31982, Kingdom of Saudi Arabia
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 3 | Page No.: 167-175







ABSTRACT
Newly formulated Saudi-1 and Saudi-2 media provided highly conspicuous growth and detection of Lagenidium giganteum zoospores on initial plating. The selectivity of Saudi-type media was based on a lecithin (soybean) and antibiotics, streptomycin and ampicillin. The Saudi-1 media supported the vegetative growth of Lagenidium giganteum (under continuous 60 W light source to get conspicuous growth). The Saudi-2 media was more defined and selective than Saudi-1. The liquid proflo PB/3 media was the best growth medium for inducing sporulation. The LC50 values obtained in this study were higher than those reported previously. One possible reason for the discrepancies in LC50 values of L. giganteum zoospores against mosquito larvae might be due to the difference in the handling techniques of treatment concentrations. On the other hand, perhaps a more important factor is the type of water (original medium) used in the previous experiments. The present study utilized sterile volvic water (double distilled), whereas the previous studies used distilled water. In conclusion, new isolate of L. giganteum is reported along with their pathogenicity towards mosquito larvae and their zoosporogenesis pattern. The study showed an excellent potential for future investigations on the use of newly developed isolate Saudi media for mosquito pathogeneic fungus for its maintenance and zoospores release.
PDF Abstract XML References Citation
How to cite this article
M.S. Shathele, 2009. New Isolate Media of the Mosquito Pathogenic Fungus Lagenidium giganteum (Oomycetes: Lagenidiales) for Fungal Maintenance and Zoospores Release. Journal of Entomology, 6: 167-175.
DOI: 10.3923/je.2009.167.175
URL: https://scialert.net/abstract/?doi=je.2009.167.175
DOI: 10.3923/je.2009.167.175
URL: https://scialert.net/abstract/?doi=je.2009.167.175
INTRODUCTION
Biological control of mosquito larvae with predators and other bio-control agents seems to be more-effective and eco-friendly approach by avoiding the use of synthetic chemicals and concomitants damaging the environment. Introduction of an auto-reproducing predator into the ecosystem may provide sustained biological control of pest populations. Lagenidium giganteum Couch, a water mold of the Class Oomycetes, is a facultative parasite of mosquito larvae that showed to be a promising biological control agent (Brey and Remaudiere, 1985). Recently, its status has changed and it is available for mosquito control operations (Domnas and Warner, 1991). This pathogenic specie has been isolated mainly from North America. However, literature indicates that isolations were also performed in other places of the world (Kerwin et al., 1994; Kerwin and Peterson, 1997; Lacey and Undeen, 1986; Washino, 1977). Although, some of the cultures were not maintained for further evaluation and comparison. Recently, Kerwin (1994) and Washino (1977) isolated another strain from California. Lagenidium giganteum Couch is a naturally occurring entomopathogenic fungus that attacks a broad spectrum of mosquito species and appears to be restricted to this group (Berbee and Kerwin, 1993). The non-discriminating host ranges within the insect family Culicidae or mosquitoes resulted from the ability of fungus to differentiate between the cuticle of mosquito larvae and other aquatic animals (Kerwin, 1994). Upon application of media to a breeding site, the fungus is activated in water where motile biflagellate zoospores are produced. Unlike bacterial larvicides which must penetrate the foliage and be ingested by the larvae, the free-swimming zoospore actively seeks out its larval mosquito host and infects resident larvae. One to three days after infection by L. giganteum, the mosquito larvae die, new spores are produced and larva-to-larva transmission continues. The main objective of this study was to develop a medium that is more selective and supportive to vegetative growth of Lagenidium giganteum (under continuous 60 W light source) to get conspicuous growth and would promote the expression of observable zoospores upon initial isolation.
MATERIALS AND METHODS
An isolates strain of L. giganteum was found in mosquito habitats in the Uraba area Northwest, Louisiana (LA) infecting Culex quinquefasciatus sentinel larvae in semi-permanent pools isolated by using sentinel cages baited with Cx. quinquefasciatus. The experiment was carried out during 2006-2007.
Maintenance and Culture Conditions
A strain of Lagenidium giganteum was used in all the experiments. The stock cultures of Lagenidium giganteum were maintained at 28°C on a new media (Saudi-1) which supports vegetative growth of Lagenidium giganteum (under continuous 60 W light source to get conspicuous growth. The medium consisted off 200 mL of double purified sterile water (volvic water) in a 500 mL flask, 0.26 g. Bacto-peptone (Difco Laboratories, Detroit, MI), 0.26 g. Bacto-yeast extract (Difco), 0.66 g. glucose, 0.66 mL CaCl2 stock solution (0.075 g mL 1), 0.07 g cholesterol from sigma (St. Louis, Mo), 0.2 g lecithin (soybean), sigma, pre-dissolved in 5-7 mL volvic water using heat and stir bar and 2.66 g Bacto-agar (Difco). The whole mixture was autoclaved for 23 min and cooled slightly. Ten milliliter of antibiotics (1.0 g streptomycin/50 mL autoclaved dh2o, filtered into 1.0 g ampicillin/50 mL autoclaved dh2o, using Millipore filter) were added to the medium. The retention of a high level of pathogenicity and maximum capacity to produce infective units are primary considerations in culturing entomo-pathogenic fungi (Washino, 1977; Fargues and Robert, 1983; Berbee and Kerwin, 1993; Kerwin et al., 1988; Gotelli, 1943).
Protocol for obtaining Lagenidium giganteum Zoospores
Defined medium, designated as liquid proflo PB/3 media, was the best growth medium for inducing sporulation. This liquid PB-3 (Saudi-2) fungal medium consists off 1 L Volvic water; 2 g proflo cottonseed extract, sigma, 1.5 g glucose, 0.5 g lactalbumin hydrolysate (Sigma L-0375), 1.0 g dehydrated egg yolk, sigma, 3.0 mL CaCl2, stock solution (0.075 g mL 1), 3.0 mL MgC12, stock solution (0.10 g mL 1), 1.0 mL Fe (NO3)3, stock solution (0.81 g/100 mL), 1.0 mL corn oil. 0.1 g fish peptone was added to correct the pH to 7.5, adjusted with 1.0 M NaOH using ph meter. One litter PB-2 media in a 2 L Erlenmeyer wide bottom flasks was inoculated with eight of 4.0 mm square agar cuts of Lagenidium giganteum of 6 days old culture plates. The necessary precaution was taken not to exceed the designated cuts. Then the Erlenmeyer flasks were capped with aluminum foil. The medium was grown by shaking in an enclosed gyratory shaker for a minimum period of seven days at 125 rpm and under continuous light condition of 60 W lamps at 28°C. Necessary precautions were taken to avoid the expose of culture to any kind of noise or sound or air current, because this will significantly impede spore release. Zoospores were prepared by Filtering approximately 250 mL of 7 days old liquid culture media, collecting the large clumps mycelium on Wattman qualitative filter paper (Grade-415) by vacuum filtration. The collected hyphae were then suspended in de-ionized water. Then, the mycelium was divided among ten Pyrex glass pans, each filled with 460 mL volvic water. The Pyrex glass pan size was 11¾ inches long x 7½ inches wide x 1¾ inch high. An effort was made to keep the volume to 4 carefully minced small tea spoons (approx. equal to 20 mL re-suspended in original media of equal volume) of mycelial filtrate /pan. However, addition of excess fungus to the pans will significantly impede spore release. The Pyrex pans were kept at 28°C for a minimum period of 18 h. Approximately, after 24 h, visible gray white dense like clouds began to appear near the surface which were siphoned into 1500 mL glass beakers. Further precautions were also taken to avoid zoospore encystment including wide bore siphon tubing, two sided light source, minimized airflow and complete quietness were applied.
Mosquito Rearing
Mosquitoes Larvae, Culex quinquefasciatus, used in the experiments were obtained from the colony established from egg rafts in the laboratory. The Culex quinquefasciatus adults were maintained at 28°C under total darkness and kept in covered screen cages. Mosquito larvae were fed on a mixture of fish food flakes and brewer’s yeast and maintained in Pyrex glass pans measuring (9.8×14.2×2.1 cubic inches). Adult mosquitoes were fed on a 10% aqueous solution of honey and mice blood meals two to three times per week. Mosquitoes lay eggs one at a time and they float on the surface of the water. In the case of Culex species, the eggs are stuck together in rafts of 200 or more. Most eggs hatch into larvae within 48 h. Water (incubation media) is a necessary part of their habitat. The larva (larvae-plural) lives in the water and comes to the surface for breathing. Larvae shed (molt) their skins four times, growing larger after each molting. Most larvae have siphon tubes for breathing and hang from the water surface (Bidllngmayer, 1968). The larvae feed on microorganisms and organic matter in the water. On fourth molting, the larva changes into pupa (Beatty and Marquardt, 1996). Each first instars Culex quinquefasciatus larva consists off seven body segments i.e., head, neck, chest, (midgut; midgut and lower gut), gills and siphons and measures approximately 6 mm. larvae were used for assays 1-2 days after hatching, or immediately as new hatched larvae depending on experimental requirements.
Zoospore concentrations were measured by taking ten 100.0 µL samples, drawn carefully at a depth of 1 cm from the water surface and at different spots from the Pyrex pans containing the zoospore solutions. Each sample was fixed by adding 100 µL of 3% glutaraldehyde solution. There are two published LD50 values for Lagenidium giganteum against mosquito’s larvae of 400-1400 zoospores/mL (Lord and Undeen, 1985; Rueda et al., 1991; Washino et al., 1976; Domnas et al., 1994). Since, the number of zoospores obtained in the experiments at different times from fungus in pure culture was variable, therefore it was difficult to have standard inoculums for use in definitive experiments. Moreover, because it was a strain of L. giganteum, without a known killing dose, the following experiment was carried out.
Killing Dose Experiment
Zoospore concentration in a stock suspension was estimated with a hemacytometer under 40 x magnifications by taking the average of four readings. Aliquots of the stock suspension containing zoospores (<1 h old) were added to Pyrex glass pans containers (3.5 inch diameter x 2.5 inch height) containing 100 mL volvic water and 20 first instars C. quinquefasciatus larvae those were reared from non-sterile egg rafts to yield final concentrations of 2000, 1800, 1500, 1400, 1350, 1300, 1200, 1000, 850, 700, 500, 400, 350, 250, 100, 50 and 0.0 (control) zoospores/mL. It is pointed out that sometimes, the stock suspension will give high concentration of zoospores/mL. Therefore, the numbers of ml’s were adjusted by adding volvic water as a dilution to obtain the desired volume of 20 mL/container ranging from (0.03+0.97 mL volvic) to 1 mL of zoospore suspension.
Another batch was used for sterile egg rafts. For the first five days, the pans were checked every 12 h and the dead or moribund larvae were removed to prevent fungal recycling. The cadavers were examined microscopically for 48 h to verify the presence or absence of L. giganteum. Only visible infected cadavers were considered to be killed by the fungus and counted. The pans without zoospores served as controls for determining the direct effect of water on mosquito larvae. Beginning 24 h from initiation of the assay, live larvae were fed on commercial Tetramin fish flakes. The containers were kept at 26±3°C under continuous light. Two to six dead larvae per cup from each concentration were examined microscopically to verify fungal Infection. The experiment was conducted three times on six months intervals. There were six replicates in each trial. Mean number of dead larvae were reported.
RESULTS AND DISCUSSION
Killing Dose of Inoculums
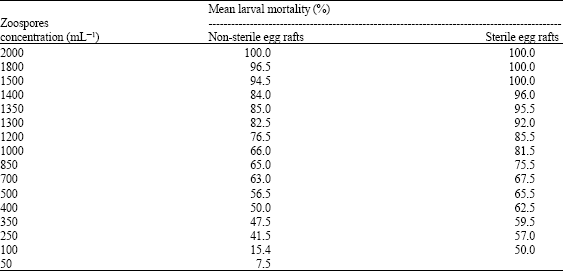
Results showed that a zoospore concentration of 2000 zoospores mL 1 produced 100% larval mortality (Table 1). However, the LC50 values with 95% confidence interval were (400 and 450 and (45 and 50) zoospores/ml for both the non-sterile and sterile egg rafts, respectively. Other researchers showed that a North Carolina isolate of L. giganteum, when assayed, produced LC50 values with 95% confidence intervals of (25.3-36.2) zoospores mL 1. The LC50 values found in this study were higher than those reported previously (Kerwin et al., 1990; Kramer, 1990; Patel et al., 1990). However, L. giganteum infectivity to mosquito larvae has been shown to vary among different isolates (Axtell et al., 1982; Jaronski and Axtell, 1983a, b; Jaronski et al., 1983; Domnas et al., 1994). It was observed in this investigation that one possible reason for the discrepancies in LC50 values of L. giganteum zoospores against mosquito larvae might be due to the difference in the handling techniques of treatment concentrations. The other most important factor could be the type of water (original medium) used in the experiment. In this study, sterile volvic water (double distilled water) was used, whereas the previous studies used only ordinary distilled water. Woodring et al. (1995) reported approximately 50% infection for all larval instars of Culex tarsalis with a concentration of 10 zoospores mL 1 and using rice-field water (Woodring et al., 1995). Lord and Undeen (19855) reported the LC50 value of 12.9 zoospores mL 1 in a 0.2 g L 1 peptone medium, 133 zoospores mL 1 in field water and 124 zoospores mL 1 in de-ionized water (Lord and Undeen, 1985). They also observed that dilutions of Lagenidium giganteum zoospores in de-ionized water caused rapid decrease in zoospore activity, thus reducing the Infection.
| Table 1: | Various zoospores concentration and mortality percentage of first instars larvae |
 | |
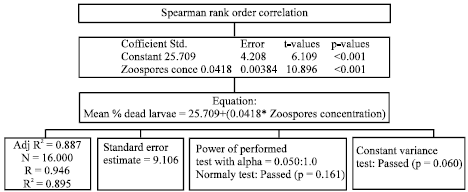 | |
| Fig. 1: | Spearman Rank Order Correlation |
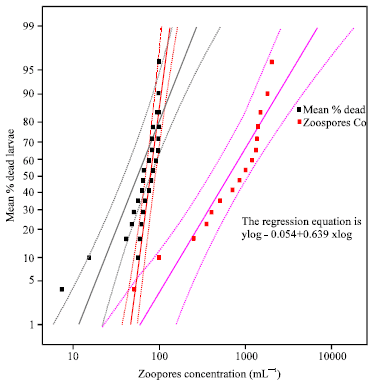 | |
| Fig. 2: | Log concentration probit regression lines for killing of Culex quinquefasciatus first instars larvae by Lagenidium giganteum |
| Table 2: | Analysis of variance |
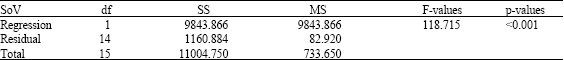 | |
They further suggested that osmotic stress produced by low solute concentrations inactivated the Lagenidium giganteum zoospores. This decline in zoospore activity was also observed in other fungal zoospores using distilled water (Kerwin and Washino, 1988; Kramer, 1990). In this study, the Spearman Rank Order Correlation (Fig. 1) show that the pair(s) of variables with positive correlation coefficients and p<0.05 tended to increase together. Table 2 shows the analysis of variance. Whereas, in the case of pairs with negative correlation coefficients and p-values below 0.05, one variable tended to decrease, while the other showed increases.
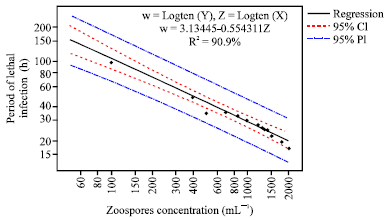 | |
| Fig. 3a: | Regression plot of lethal period of infection various. Zoospores concentration and Period of 100% mortality (h) of first instars larvae from non sterile egg rafts |
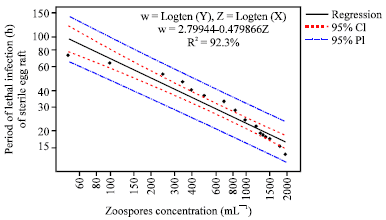 | |
| Fig. 3b: | Regression plot of various zoospores concentration and Period of 100% mortality (h) of first instars larvae from sterile egg rafts |
For the pairs with p-values greater than 0.05, the relationship was not significant between the two variables (Fig. 2) and the regression equation is ylog = -0.054+0.639 xlog. Also, the number of zoospores obtained at different times from fungus in pure culture was variable in the experiments. Researchers showed that large numbers of zoospores (107-108/L) could be obtained from L. giganteum (Woodring et al., 1995; Kerwin et al., 1994; Kerwin et al., 1988). Moreover, it was difficult to secure standard inoculums for use in definitive experiments and to have a high zoospore concentration of 2000 mL 1. Besides that, the culturing techniques may affect the results of concentration assays especially using the new strain. However, LC50 values differed between the isolates indicating that some strains of Lagenidium giganteum may be more virulent than others. In this study, 2000 Pores concentration 1500 mL 1 inoculums was considered as standard for all experiments.
Period of 100% Mortality
The lowest zoospores concentration caused the first larvae instars to have more life span among all the zoospore concentrations (50.0 mL 1), F = 139.69; p<0.00. The difference in period of lethal infection among the different zoospore concentrations was significant with an increase in concentration (Fig. 3a, b). The period of lethal infection decreased with the increase inzoospore concentration from 50 to 2000 mL 1.
 | |
| Fig. 4: | Fluorescence microscopy image of cells incubated with fluorescent dye. Zoospores, cysts (A). Bar = 5µm larvae of Culex quinquefasciatu s were introduced to a suspension of Lagenidium giganteum zoospores germination tubes (B) of known concentration and then examined with phase-contrast optics, Lagenidium giganteum rapidly colonized the host cuticle |
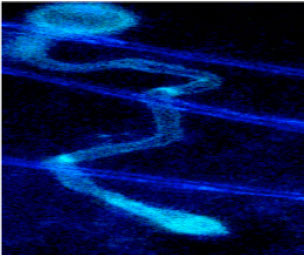 | |
| Fig. 5: | Fluorescence microscopy image of cells incubated with fluorescent dye. Zoospores, cysts (A). Bar = 5µm larvae of Culex quinquefasciatu s Show that a single germ tube (B)emerged from most of the zoospores and typically extended up to a certain distance of up to 6 mm before further branching or penetrating. By 2 h, more individuals had produced appressorial and the expansion of these structures shows that Penetration of the host cuticle was accomplished by a very narrow (approx. 0.3 µm in diameter) tube (D), which grew out from that part of the appressorium (C) in intimate contact with the cuticle surface (E) |
This could be attributed to the fast depletion of larvae nutrients due to the presence of large number of zoospores (Fig. 4, 5).
ACKNOWLEDGMENTS
The author is indebted to J. Kerwin, University of California, Davis and R. Robinson, Department of Microbiology, Arizona State University, Arizona, USA.
REFERENCES
- Axtell, R.C., S. Jaronski and T. Merriam, 1982. Efficacy of the mosquito fungal pathogen Lagenidium giganteum (Oomycetes: Lagenidiales). Proc. Calif. Mosq. Vector Cont. Assoc., 50: 41-42.
Direct Link - Berbee, M. and J.L. Kerwin, 1993. Ultra structural and light microscopic localization of carbohydrates and peroxidase/catalases in Lagenidium giganteum zoospores. Mycologia, 85: 734-743.
Direct Link - Domnas, A., S.S. Biswas and P.A. Gallagher, 1994. Squalene metabolism in two species of Lagenidium. Canadian J. Microbiol,. 40: 523-531.
Direct Link - Domnas, A.J. and S.A. Warner, 1991. Biochemical activities of entomophagous fungi. Crit. Rev. Microbiol., 18: 1-13.
Direct Link - Kerwin, J.L., D.D. Dritz and R.K. Washino, 1994. Pilot scale procution and application in wildlife ponds of Lagenidium giganteum (Oomycetes: Lagenidiales). J. Am. Mosq. Control Assoc., 10: 451-455.
Direct Link - Kerwin, J.L., D.A. Dritz and R.K. Washino, 1990. Confirmation of the Safety of Lagenidium giganteum (Oomycetes: Lagenidiales) to Mammals. J. Econ. Entomol., 81: 374-376.
Direct Link - Kerwin, J.L. and R.K. Washino, 1988. Field evaluation of Lagenidium giganteum (Oomycetes: Lagenidiales) and description of a natural epizootic involving a new isolate of the fungus. J. Med. Entomol., 25: 452-460.
Direct Link - Kramer, V.I., 1990. Laboratory evaluation of Lagenidium giganteum (Oomycetes: Lagenidiales) in water from Contra Costa County, California, mosquito sources. J. Am. Mosq. Control Assoc., 6: 79-83.
Direct Link - Lacey, L.L. and A.H. Undeen, 1986. Microbial control of black flies and mosquitoes. Annu. Rev. Entomol., 31: 265-296.
CrossRefDirect Link - Lord, J.C. and A.H. Undeen, 1985. Solute effects on Lagenidium giganteum zoospore motility and bioassay reproducibility. J. Invertebrate Pathol., 46: 160-165.
CrossRef - Patel, K.J., L.M. Rueda and R.C. Axtell, 1990. Comparisons of different types and concentrations of alginates for encapsulation of Lagenidium giganteum (Oomycetes: Lagenidiales), a fungal pathogen of mosquito larvae. J. Am. Mosq. Control Assoc., 6: 101-104.
Direct Link - Rueda, L.M., K.J. Patel and R.C. Axtell, 1991. Comparison of floating and sinking encapsulated formulations of the fungus Lagenidium giganteum (Oomycetes: Lagenidiales) for control of Anopheles larvae. J. Am. Mosq. Control Assoc., 7: 250-254.
Direct Link - Woodring, J.L., H.K. Kaya and J.L. Kerwin, 1995. Lagenidium giganteum in Culex tarsalis larvae: Production of infective propagules. J. Invertebrate Pathol., 66: 25-32.
Direct Link