
Hooshang Rafiee Dastjerdi
Department of Plant Protection, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Mir Jalil Hejazi
Department of Plant Protection, Faculty of Agriculture, University of Tabriz, Tabriz, Iran
Ghadir Nouri Ganbalani
Department of Plant Protection, Faculty of Agriculture, University of Mohaghegh Ardabili, Ardabil, Iran
Moosa Saber
Department of Plant Protection, Faculty of Agriculture, Higher Education Institute of Maragheh, Maragheh, Iran
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 3 | Page No.: 161-166







ABSTRACT
In this research, the effects of profenofos, thiodicarb, hexaflumuron and spinosad were studied on functional response of Habrobracon hebetor to different densities of last instar larvae of Anagasta kuehniella. Two day-old mated females were exposed to LC25 of the insecticides and distilled water as control. Host densities of 2, 4, 8, 16, 32 and 64 were offered to treated females wasps for 24 h in 10 cm petri dishes. Experiments were conducted in 8 replications and were carried out in controlled conditions, 26± 2°C, 70± 5% RH and 16:8 (L:D) h. The type of functional response was determined using logistic regression and the parameters, searching efficiency (a) and handling time (Th) were estimated by non-linear regression using SAS software. Functional response was type II in control and all of insecticide treatments. Searching efficiency in control and insecticide treatments were 0.0935, 0.0132, 0.0511, 0.0864 and 0.0905 h-1 and handling times were 0.4542, 1.0646, 0.5381, 0.5275 and 0.4896 h, respectively. The maximum attach rates (24/Th) were estimated 22, 52, 44, 45 and 49, respectively. Spinosad and hexaflumuron had the most and the lowest effect on searching efficiency of H. hebetor, respectively.
PDF Abstract XML References Citation
How to cite this article
Hooshang Rafiee Dastjerdi, Mir Jalil Hejazi, Ghadir Nouri Ganbalani and Moosa Saber, 2009. Effects of Some Insecticides on Functional Response of Ectoparasitoid, Habrobracon hebetor (Say) (Hym.: Braconidae). Journal of Entomology, 6: 161-166.
DOI: 10.3923/je.2009.161.166
URL: https://scialert.net/abstract/?doi=je.2009.161.166
DOI: 10.3923/je.2009.161.166
URL: https://scialert.net/abstract/?doi=je.2009.161.166
INTRODUCTION
There is increasing interest in studying the effects of insecticides on the biocontrol agents in IPM programs and reducing the application of insecticides in fields. The ectoparasitoid, Habrobracon hebetor (Say) is an important biocontrol agent of lepidopteran pests in fields and stored products. Among parasitoids, H. hebetor gets more attention from Iranian researchers. The effects of insecticides on efficiency of this wasp can be compared by estimation and comparison of functional response parameters. In this study, we determined if there is any difference between functional responses of wasp that are exposed to insecticides. Functional response is defined as the number of hosts attacked by an individual natural enemy in relation to host density over a given time interval (Solomon, 1949). The most widely used model for describing the functional response of predator/parasitoid is the disk equation of Holling (1959):
Na = aTtN0/(1+aThN0) |
Where:
| Na | = | No. of host attacked |
| N0 | = | Initial No. of hosts |
| Tt | = | Time of exposure between wasps and hosts |
| a | = | Rate of successful hosts attacked |
| Th | = | Handling time |
For modeling the type III functional response, attacked rate (a) must be in equation 1 as a function of prey/host density. In the simplest generalized form, attacked rate is a function of initial number of prey/host density:
a = (d+bN0)/(1+cN0) |
where, b,c and d are constants that must be estimated (Juliano, 1993; Juliano and Williams, 1987; Hassell et al., 1977).
Among the types of functional response, type II and III have received the most attention, because most natural enemies show these types. The functional response of parasitoid is a main factor regulating the population dynamics of parasitoid-host systems. It describes the rate at which a biocontrol agent parasitized its host at different host densities and can thus determine the efficiency of parasitoid in regulating parasitoid populations. However, certain predators and parasitoids exhibiting type II response have been successfully established and managed prey populations (Hughes et al., 1992; Fernández-Arhex and Corley, 2003). The functional response curves can be differentiated by evaluating the parameters, viz., coefficient of attack rate and handling time (time spent by predator in attacking, killing, subduing and digesting the prey). The coefficient of attack rate estimates the steepness of the increase in predation with increasing prey density and handling time helps estimate the satiation threshold (Pervez and Omkar, 2005). There are no studies on effects of insecticides on functional response of H. hebetor. In this research we assayed the effects of sublethal concentration of profenofos, spinosad, thiodicarb and field recommended concentration of hexaflumuron on functional response of ectoparasitoid H. hebetor.
MATERIALS AND METHODS
The study was conducted in 2007 and 2008 in the Department of Plant Protection, University of Tabriz, Iran.
Insects
Adults of H. hebetor were obtained from an insectarium maintained by Plant Protection Bureau of Bilehsavar in Ardabil Province, Iran and reared on 5th instars of A. kuehniella in the laboratory. A. kuehniella cultures were maintained in clear plastic boxes (40x25x15 cm) containing 1 kg of wheat flour. A colony of H. hebetor was established into plastic petri dishes (10 cm in diameter) and then this colony was divided into two groups, one used to maintaining colony and the other group used in experiments. Fifth instars of A. kuehniella were used for both colony maintenance and experiments. Rearing conditions were 26± 2°C temperature, 70± 5% RH and a photoperiod of 16: 8 h (L:D).
Chemicals
Insecticides tested were hexaflumuron (Consult® 10EC) and spinosad (SpinTor® 25SC) both from Dow AgroSciences, (http://dowagro.com/uk/products); profenofos (40EC, Golsam, http://www.golsam.com) and thiodicarb (80DF, Gyah, http://www.gyah.ir).
Functional Response Study
Exposure cages (100x100x10 mm) were used for experiments (Saber et al., 2005). Glass surfaces of cages were sprayed with 2 mL of aqueous solutions of LC25 of profenofos, spinosad, thiodicarb and field recommended dose of hexaflumuron by potter spray tower. Concentrations used were 4.45, 5.04, 23.46 and 250 mg a.i. L-1. Triton X-100 was used as the wetting agent at a concentration of 555 ppm in this experiment. The control plates were sprayed with distilled water plus Triton X-100. The operating pressure was 0.5 bar and the mean spray deposit was 1.68 ± 0.04 μL cm-2. As adult wasps emerged then 30 females and 30 males of H. hebetor were left to mating up to 24 h in glass tubes. Adult females were exposed to LC25 of profenofos, spinosad, thiodicarb and field recommended dose of hexaflumuron. After 24 h, randomly selected 6 alive females were transferred individually to plastic petri dishes (30 mm in diameter) with the densities (2, 4, 8, 16, 32 and 64) of 5th instars of A. kuehniella. Petri dishes were kept in incubators under the same conditions as described above and the wasps were provided with honey. The numbers of parasitized and remaining alive larvae were registered after 24 h. Eight replicates were conducted for each insecticide treatments. The functional response parameters were estimated through the nonlinear least square method (procedure NLIN) with SAS for Windows® Release 9.0 (SAS Institute, 2002). The selection of appropriate model (type II or III functional response) requires logistic regression of the proportion of host parasitized versus the number of hosts present (Juliano 1993). Estimation and comparison of parameters are best done through non linear least squares regression techniques (Williams and Juliano, 1985, Juliano and Williams, 1987). Juliano (1993) described appropriate non-linear curb fitting procedures for functional response experiments.
RESULTS
The negative values for the linear parameters obtained in the present study confirm the type II response for all treatments used. The logistic regression model thus can be recommended as a tool for further analyzing functional response curves. The logistic regression for all treatments had a significant linear parameter (Table 1). The number parasitized by wasps in all treatments increased with increasing host density but the proportion of host parasitized by wasps declined with increasing host density (Fig. 1, 2). This suggests that all treatments exhibited a type II response. Their handling times and coefficient of attack rates are shown in Table 2, which shows that control had the shortest handling time and the highest coefficient of attack rate, followed by hexaflumuron, thiodicarb, profenofos and spinosad, respectively.
| Table 1: | Results of the logistic regression analysis of the proportion of A. kuehniella larvae parasitized by H. hebetor against the initial number offered |
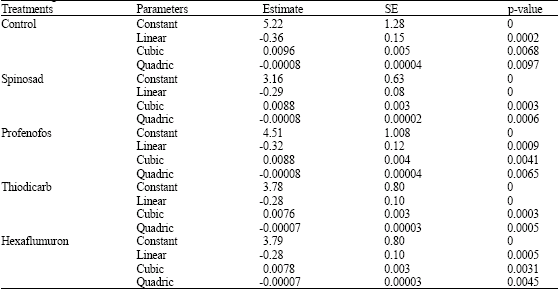 | |
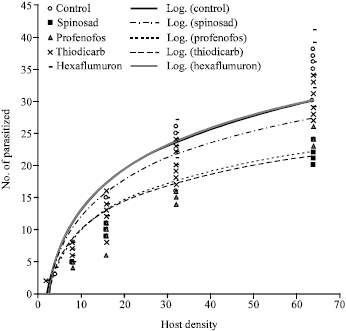 | |
| Fig. 1: | Functional response of the ectoparasitoid, H. hebetor exposed to different insecticides and in control. A type II response curve was fitted by least squares, following Holling’s (1959) equation in all treatments. Overlapping values are shown as a single dot, n = 8 for each density |
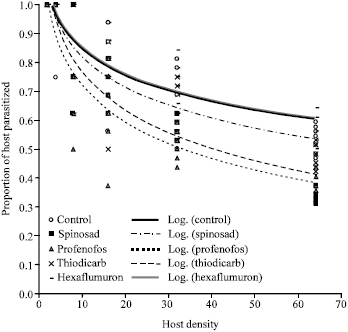 | |
| Fig. 2: | The proportion of host parasitized by ectoparasitoid, H. hebetor exposed to different insecticides and in control were declined with increasing host density. Overlapping values are shown as a single dot, n = 8 for each density |
Also, control had the highest coefficient of attack rate, followed by hexaflumuron, thiodicarb, profenofos and spinosad, respectively.
| Table 2: | Functional response parameters estimated for H. hebetor exposed to insecticides and in control |
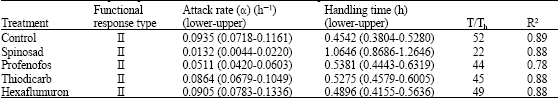 | |
Spinosad significantly affected handling time. Both handling time and coefficient of attack rate were dependent on the treatments (Table 2).
DISCUSSION
The coefficient of attack rate and handling time are the parameters used to find out the magnitude of functional responses. Based on overlapping, the coefficient of attack rate of spinosad and profenofos differed significantly from the two other insecticides and the control, indicating that spinosad and profenofos induced different response from the ectoparasitoid when the host densities are increased. This also indicates that treated parasitoid exhibiting similar functional response curves cannot be deemed to respond similarly. The attack rates coefficients of H. hebetor exposed to hexaflumuron and thiodicarb did not vary significantly as compared to those observed with ectoparasitoids exposed to water. This indicates that hexaflumuron and thiodicarb had the least effect on the searching efficiency of the wasps, as compared to the two other insecticides.
The wasps exposed to insecticides had higher handling time, but spinosad only significantly affected handling time. Similar insignificant effects of factors on functional response were reported in four heteropteran predators preying on white flies and thrips (Montserrat et al., 2000). The handling time estimate is the cumulative effect of time taken during capturing, killing, subduing and digesting the prey (Veeravel and Baskaran, 1997). Thus, significant variation in the estimates of handling times of a single predator on different prey species indicates that any one of these integral components of the handling time might have been negatively.
Claver et al. (2003) studied impact of cypermethrin on functional response of Acanthaspis pedestris and indicated that cypermethrin negatively affected the functional response events such as attack ratio, handling time and rate of discovery. This insecticide also reduced the predatory efficiency and prolonged the mating events in A. pedestris. The type II (decelerating curve) of functional response was altered into a type IV (dome-shaped curve) by cypermethrin (on the functional response). They indicated that the type of functional response in predatory thrips differed depending on the types and doses of the pesticides exposed. In general, between insecticides tested, hexaflumuron and spinosad had lowest and highest adverse effects on functional response paramters of H. hebetor, respectively.
ACKNOWLEDGMENTS
The study received financial support from Postgraduate Education Bureau of the University of Tabriz and University of Mohaghegh Ardabili.
REFERENCES
- Claver, M.A., B. Ravichandran, M.M. Khan and D.P. Ambrose, 2003. Impact of cypermethrin on the functional response, predatory and mating behaviour of a non-target potential biological control agent Acanthaspis pedestris (St�l) (Het., Reduviidae). J. Applied Entomol., 127: 18-22.
CrossRefDirect Link - Juliano, S.A. and F.M. Williams, 1987. A comparison of methods for estimating the functional response parameters of the random predator equation. J. Anim. Ecol., 56: 641-653.
Direct Link - Hassell, M.P., J.H. Lawton and J.R. Beddington, 1977. Sigmoid functional responses by invertebrate predators and parasitoids. J. Anim. Ecol., 46: 249-262.
Direct Link - Holling, C.S., 1959. Some characteristics of simple types of predation and parasitism 1. Can. Entomol., 91: 385-389.
CrossRef - Hughes, R.D., L.T. Woolcock and M.A. Hughes, 1992. Laboratory evaluation of parasitic hymenoptera used in attempts to biologically control aphid pests of crops in Australia. Entomol. Exp. Appl., 63: 177-185.
CrossRef - Fernandez-Arhex, V. and J.C. Corley, 2003. The functional response of parasitoids and its implications for biological control. Biocontrol Sci. Technol., 13: 403-413.
CrossRef - Montserrat, M., R. Albajes and C. Castane, 2000. Functional response of four heteropteran predators preying on greenhouse whitefly (Homoptera: Aleyrodidae) and western flower thrips (Thysanoptera: Thripidae). Environ. Entomol., 29: 1075-1082.
CrossRef - Pervez, A. and Omkar, 2005. Functional responses of coccinellid predators: An illustration of a logistic approach. J. Insect Sci., 5: 5-5.
PubMedDirect Link - Solomon, M.E., 1949. The natural control of animal populations. J. Anim. Ecol., 18: 1-35.
CrossRefDirect Link - Veeravel, R. and P. Baskaran, 1997. Functional and numerical responses of Coccinella transversalis Fab. and Cheilomenes sexmaculatus Fab. feeding on the melon aphid, Aphis gossypii Glov. Insect Sci. Applied, 17: 335-339.
CrossRefDirect Link - Williams, F.M. and S.A. Juliano, 1985. Further difficulties in the analysis of functional response experiment and a resolution. Can. Entomol., 117: 631-640.
Direct Link - Saber, M., Hejazi M.J., K. Kamali and S. Moharramipour, 2005. Lethal and sublethal effects of fenitrothion and deltamethrin residues on the egg parasitoid Trissolcus grandis (Hymenoptera: Scelionidae). J. Econ. Entomol., 98: 35-40.
Direct Link