
R. Cossio-Bayugar
Centro Nacional de Investigacion Disciplinaria en Parasitologia Veterinaria, Institut Nacional de Investigaciones Forestales Agricolas y Pecuarias INIFAP. Carr. Fed. Cuernavaca-Cuautla 8534, Jiutepec, Morelos ,CP 62550, Mexico
E. Miranda-Miranda
Centro Nacional de Investigacion Disciplinaria en Parasitologia Veterinaria, Institut Nacional de Investigaciones Forestales Agricolas y Pecuarias INIFAP. Carr. Fed. Cuernavaca-Cuautla 8534, Jiutepec, Morelos ,CP 62550, Mexico
D. Portilla-Salgado
Centro Nacional de Investigacion Disciplinaria en Parasitologia Veterinaria, Institut Nacional de Investigaciones Forestales Agricolas y Pecuarias INIFAP. Carr. Fed. Cuernavaca-Cuautla 8534, Jiutepec, Morelos ,CP 62550, Mexico
J. Osorio-Miranda
Departament of de Ectoparasitos y Dipteros, Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria SAGARPA
Journal of Entomology
Year: 2009 | Volume: 6 | Issue: 2 | Page No.: 117-123







ABSTRACT
This study was done with the objective of developing a methodology for cholinesterase and carboxylesterase expression measurement in acaricide resistant R. microplus, for that purpose the expression levels for these enzymes were measured by real time PCR quantification of mRNA specific detection, comparing acaricide sensitive with multiple acaricide resistant strains of ticks known as Mora and San Alfonso. Acaricide susceptible ticks were used as standard cholinesterase and carboxylesterase expression level and adjusted as a baseline of 1 Relative Expression Units (REU). A statistical significance was observed in cholinesterase gene expression level as 13.07 ± 3.49 REU for Mora strain and 10.81 ± 2.98 REU for San Alfonso strain compared to the susceptible strain. Also, carboxylestrase expression found statistically significant for Mora and San Alfonso strains (6.9 ± 1.14 and 12.11 ± 1.81 REU, respectively) compared to the susceptible strain. Present results proved that the carboxylestrase and cholinesterase genes expression increased by acaricide pesticide exposure in Mora and San Alfonso, R. microplus, which explained as an overexpression of AchE2 at the singanglion level for OP resistant ticks, as well as increased levels of esterase gene CzEST9 implicated in pyrethroid resistant strains.
PDF Abstract XML References Citation
How to cite this article
R. Cossio-Bayugar, E. Miranda-Miranda, D. Portilla-Salgado and J. Osorio-Miranda, 2009. Quantitative PCR Detection of Cholinesterase and Carboxylesterase Expression Levels in Acaricide Resistant Rhipicephalus (Boophilus) microplus. Journal of Entomology, 6: 117-123.
DOI: 10.3923/je.2009.117.123
URL: https://scialert.net/abstract/?doi=je.2009.117.123
DOI: 10.3923/je.2009.117.123
URL: https://scialert.net/abstract/?doi=je.2009.117.123
INTRODUCTION
Acetylcholinesterase (AChE) hydrolyzes the neurotransmitter acetylcholine regulating the cholinergic nervous transmission in a wide range of organisms across the phylogenetic tree including arthropods (Boublik et al., 2002). Organophosphorous (OP) pesticides react with a serine within the AChE active site, irreversibly phosphorylating it and rendering the enzyme inactive (Weill et al., 2002). As a result of AChE phosphorylation, synaptic transmission fails to take place, leading to paralysis and death of the arthropod (Kim et al., 2003). An altered AChE capable to resist phosphorylation of the active site has been referred as responsible for OP pesticides resistance mechanism in: Musca domestica (Kim et al., 2003), Culex pipiens (Weill et al., 2002) Anopheles albimanus (Dzul et al., 2007) and Drosophila sp. (Boublik et al., 2002). However, an altered AChE is not the only OP resistance mechanism, a secondary process is an overexpression of putative AChE enzyme directed to sequestering and detoxifying the excess of OP chemicals (Baxter and Barker, 2002). Present research indicates that some other close AChE related enzymes produces enhanced acaricide detoxification activity mediated by ester-hydrolyzing enzymes such as carboxylesterase (CE) (Hemingway and Karunaratne, 1998). AChE and CE are very close related enzymes and ubiquitous among arthropods, both enzymes specialize in hydrolyzing ester acaricide resistance has been reported that it is related to detoxification of synthetic pyrethroids mediated by ester bond hydrolysis of the acaricide by CE (De Jersey et al., 1985), other scientific reports have shown increased carboxylesterase activity in a resistant pyrethroid Mexican strain of R. microplus ticks (Jamroz et al., 2000). CE are involved in the metabolism of xenobiotics compounds (Scott, 1999; Karunaratne and Hemingway, 2001), earlier reports attribute to CE an important role in pesticides resistance in a large variety of pest arthropods including the sheep ectoparasite Lucilia cuprina, (Newcomb et al., 1997; Hartley et al., 2006), the human blood sucking mosquitoes Culex sp., Aedes sp. and Anopheles sp. (Hemingway and Karunaratne, 1998; Paton et al., 2000; Karunaratne and Hemingway, 2001) and the peach-potato aphid Myzuz persicae (Field and Devonshire, 1998; Field et al., 1999). Earlier study on R. microplus CE also has identified a linkage between this enzyme and cattle tick acaricide resistance (Jamroz et al., 2000; Baffi et al., 2008). Both AChE and CE worked simultaneously on OP and pyrethroid (PS) acaricide resistance in Australian acaricide resistant R. microplus ticks (Baxter and Barker, 2002). In this study, we focused on the quantitative expression levels of AChE and CE in order to prove the linkage of these enzymes to acaricide resistance in cattle tick Rhipicephalus microplus strains (Mora and San Alfonso).
MATERIALS AND METHODS
This study was carried out in Centro Nacional de Investigación Disciplinaria en Parasitología Veterinaria, INIFAP, México and the Departamento de Ectoparásitos y Dípteros del Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA-SAGARPA) México during the months of September 2006 to April 2007.
Ticks
All ticks strains used in this study, have been cultured under laboratory controlled conditions for many generations and normally are used as reference for the tick acaricide resistance monitoring programs of the Mexican Federal Government, these ticks were reared and maintained at: Departamento de Ectoparásitos y Dípteros del Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA-SAGARPA). Tick strains used in this study included R. microplus reference ticks previously reported by Ortiz et al. (1995), Soberanes et al. (2002) and Cossio-Bayugar et al. (2005) with known acaricide resistance or susceptibility (Table 1). Multiple resistant strain for OP and PS resistant strain named Mora (MO) (Ortiz et al., 1995) and San Alfonso (SA) (Soberanes et al., 2002) (Table 1). The acaricide susceptible reference strain was used for comparison against acaricide resistant ticks during larvae package bioassays as previously reported by Stone and Haydock (1962). Susceptible strain was also used for cholinesterase and carboxylesterase gene expression baseline adjustment in order to compare against gene expression levels of the resistant strain of ticks according to protocol previously reported by Cossio-Bayugar et al. (2005).
| Table 1: | Acaricide bioassays result on different strains and isolates of ticks. Data is represented as percentage of mortality under a standard concentration of acaricide |
 | |
Each reference strain and tick isolate used was cultured by infesting a bovine with 2x104, 10-15 day old larvae, engorged females were collected 21 days after infestation, placed in Petri dishes for oviposition in groups of ten ticks for each strain, while incubating at 28°C in 80% relative moisture until complete oviposition. The eggs were collected, weighted and aliquoted in vials with 200 mg each.
Ticks were assayed and selected for their PS and OP resistant toxicological profile demonstrated by acaricide discriminant doses bioassays (Stone and Haydock, 1962)(Table 1). Bioassays were run by using trichloro ethylene diluted acaricides at the following concentration: coumaphos 0.2%, chlorphenvinphos 0.2%, diazinon 0.08%, chlorpiriphos 0.2%, cypermetrine 0.05%, deltametrine 0.09%, flumetrine 0.01%. One milliliter of each dilution was applied evenly to a 7 by 9 cm piece of filter paper. The trichloroethylene was allowed to evaporate from the filter paper for 2 h. The treated papers were then folded in half and sealed on the slides with clips, this formed a packet into which approximately 100 larvae were placed and then the top of the packet was sealed with another clip. The packets were kept at 27°C , 92% relative humidity for 24 h, after which the packets were removed from incubation and opened, live and death larvae were counted and the data was processed as percentage of mortality for each tick group under every acaricide concentration.
Relative Quantification of Carboxylesterases and Cholinesterases
Total RNA was isolated from each R. microplus tick sample following the manufacturer’s instructions (Totally RNA kit™, Ambion, TX, USA) and treated with DNase according to manufacturer’s instructions (Turbo DNA- free™, Ambion, TX, USA). RNA was quantified spectrophotometrically by measuring the 260/280 nanometer ratio previously described by Sambrook and Russell (2001) RNA quality was verified by agarose gel electrophoresis according to earlier protocol (Sambrook and Russell, 2001). RNA was transcribed to cDNA using random decamer primers following the instructions of a commercial kit (RETROscriptTM Kit Ambion). The cDNA synthesis reaction included 5 μM Random decamers, 0.5 mM each dNTP, 10 units RNase Inhibitor, 100 units of MMLV-RT, 1X RT buffer (50 mM Tris HCl, PH 8.3, 75 nM KCl, 3 mM MgCl2, 5 mM DTT). The reaction was incubated at 44°C for 1 h, followed by a 10 min incubation at 92°C to inactivate the reverse transcriptase.
Cholinesterase (AChE) and carboxylesterase (CE) relative gene expression quantification was made by using real time PCR with taqman fluorescent probes specifically designed for CE and AChE R. microplus gene sequences (GenBank access AF182283, AF286096, DQ533868 for CE and AJ278345, AJ278344, AJ278343, AJ278342 and AF067771 for AChE). Measurement of CE and AChE mRNA in acaricide susceptible and acaricide resistant ticks by real-time PCR was carried out using a fluorogenic 5’ nuclease assay (TaqMan® system) on an ABI Prism 7300 Sequence Detector (Applied Biosystems, CA, USA). The reactions included 900 nM of each primer, 300 nM probe, 1X TaqMan® Universal Master Mix (TaqMan buffer A 1X, 200 μM dATP, dGTP, dUTP each, 400 μM dUTP, 0.01 U μL-1 AmpErase UNG, 0.05 U μL-1 AmpliTaq Gold, 0.05% gelatin, 5 mM MgCl2) (Applied Biosystems). The real-time PCR thermal program consisted of one cycle at 50°C for 2 min and one cycle at 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. AChE and CE gene-specific PCR primers and the TaqMan® probe labeled with 6-carboxyfluorescein (FAM)/MGB designed for AChE and CE R. microplus gene sequence were as follows: forward primer for AChE AChEFor 5’- GGCACTGAAATGGATCCAGGAA-3’, reverse primer AChE AChERev 5’-CGTGACTTCACCAGGGTTACC-3’ and AChE TaqMan® probe 5’-CCAAATG CAG CAATGTT-3’ as well as forward primer for CE CEFor 3'-CGACGCATTCCTTCCAAAGATG-5', reverse primer for CE CERev 3'- TCGACGGACGCGAAGAAG-5' and CE TaqMan probe-CACTTGTAGCCATGAATC-5'. The R. microplus eukaryotic 18S rRNA (Applied Biosystems, CA, USA) probe served as endogenous controls and was labeled with MGB®. A reaction without cDNA was used as negative control. To check reproducibility, the real-time PCR analysis including the endogenous control, 18S rRNA, was repeated twice in independent experiments, with five replicates for each experiment (n = 10).
The number of RNA transcripts detected for each strain and field isolate was determined by the 7300 SDS Software v1.2.2 (Applied Biosystems, CA, USA); quantitative measurements were normalized by measuring the expression levels of 18S ribosomal RNA as internal control. The susceptible strain was considered the basal level of CE and AChE expression and assigned a Relative value of 1 Expression Unit (1 REU).
Statistical Analysis
Means and dispersion measurements of relative CE and AChE gene expression in REU from acaricide resistant strains were compared against expression means and dispersion measurements of susceptible strains by an unpaired Student's t-test using the GraphPad Software (GraphPad Software Inc. CA, USA).
RESULTS
Sensitive reference strain showed 100% mortality when exposed to all acaricides, this ticks were considered as reference acaricide sensitive gene expression and their carboxylesterase and cholinesterase gene expression values were adjusted as 1 Relative Expression Unit (REU) (Table 1, Fig. 1). Mora and S. Alfonso strains showed high levels of Pyrethroids and Organophosphorous resistance with very low mortality when exposed to all PS and OP acaricides dosages capable of killing a 100% of the reference susceptible strain (Table 1). Both strains of resistant ticks showed a statistically significant increased levels of AChE expression (p<0.0001 unpaired t-test) 13.07 ± 3.49 REU for MO strain and 10.81 ± 2.98 for SA strain compared to the susceptible strain 1REU, (Fig. 1). The levels of CE expression statistically significant increased (p<0.0001 unpaired t-test) for MO as 6.901 ± 1.14 REU and 12.11 ± 1.81 for SA strain compared to the susceptible strain 1REU (Fig. 1).
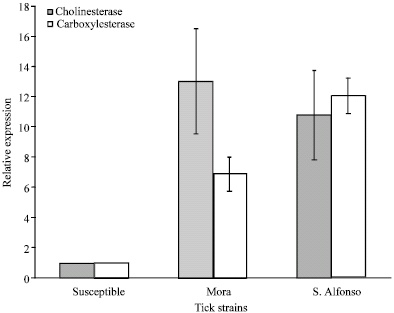 | |
| Fig. 1: | Cholinesterase and carboxylesterase gene expression relative quantification by real time PCR of acaricide susceptible and resistant R. microplus. Data is represented as Relative Expression Units (REU). Mean values and SD are shown |
DISCUSSION
Pesticide resistance in arthropods underlies a genetic mechanism that may involve several genes, achieving an inheritable pesticide resistance phenomenon. The phenotypes conferred by these genes included AChE with a low affinity for OP acaricides (Bull and Ahrens, 1988; Jamroz et al., 2000; Baxter and Barker, 2002). In present results, the statistically significant increase in AChE expression for both acaricide resistant strains of ticks when compared to susceptible reference strain, proved that the association of OP resistance of the analyzed ticks to high levels of AChE expression and may be compatible with OP acaricide sequestration scenario of cholinesterase excess which translate into enough free enzyme accomplished with the acetylcholine nerve impulse regulation, even after OP acaricide exposure, avoiding paralysis and death of the resistant ticks. AChE overexpression does not exclude that a low OP affinity AChE may be conferring resistance to present ticks strains, however to probe this phenomenon, a different set of experimental conditions beyond the reach of present assays, will be required. The AChE probes were designed to recognize the alleles AChE2 of R. microplus (accession No. AF067771.1, AJ278342.1, AJ278343.1, AJ278344.1, AJ278345.1). The AChE2 allele has been reported to be expressed within the tick's syngaglion, indicating that this AChE is participating in the actual regulation of nerve impulse transmission in synganglia, the main target for OP acaricides (Baxter and Barker, 2002). The CE probes, on the other hand, were designed to recognize the CzEST9 transcript (GenBank accession No. DQ533868.1 and AF182283.1), this enzyme transcript has been described to translate into a pyrethroid metabolizing carboxylesterase dubbed EST9, in Mexican pyrethroid resistant R. microplus (Jamroz et al., 2000; Hernandez et al., 2000; Pruett et al., 2002). This particular strain of ticks, showed a significant amount of metabolic esterase-based resistance when exposed to pyrethroid based acaricides (Jamroz et al., 2000; Pruett et al., 2002). Present experimental data showed that the CE probes detects increased transcription levels of a CzEST9 CE-like enzyme in two resistance R. microplus strains compared to the susceptible strain, although present acaricide resistant strains of ticks are different to the strain earlier reported by Jamroz et al. (2000) and Pruett et al. (2002), there are no earlier reports on the literature of Cholinesterase overexpression in OP resistance tick, there are, however earlier reports of increased levels of CEST9 on pyrethroid resistant R. microplus (Hernandez et al., 2000).
The Mexican resistant strains have been isolated from contiguous Mexican States with heavy cattle exploitation, transport of live untreated animals from one state to another may occur carrying acaricide resistant ticks with them, for we can assume that resistance genes coding for cholinesterase AChE2 allele as well as, esterase CzEST9 transcript are continuously spreading with the ticks and their cattle host, throughout cattle raising areas, this acaricide resistant spreading is in accordance to earlier reports by Soberanes et al. (2000). Present data support the hypothesis that, unless more rigorous constrains on the cattle commerce are enforced in Mexico, resistance genes and cattle tick acaricide resistance will grow to an ever bigger problem.
Based on the data presented in this study, we can conclude that part of the OP and pyrethroid acaricide resistance in R. microplus, is explained as an overexpression of AChE2 at the singanglion level for OP resistant ticks, as well as increased levels of esterase gene CzEST9 implicated in pyrethroid resistant strains. Further studies are necessary to analyze the expression levels of AChE, CE and other genes in reference and field ticks in order to probe the usefulness of these genes as molecular probes for diagnostic purposes.
ACKNOWLEDGMENT
This research was partially supported be SEP-CONACYT Grant J39752-Z.
REFERENCES
- Baxter, G.D. and S.C. Barker, 2002. Analysis of the sequence and expression of a second putative acetylcholinesterase cDNA from organophosphate-susceptible and organophosphate-resistant cattle ticks. Insect Biochem. Mol. Biol., 32: 815-820.
CrossRef - Boublik, Y., P. Saint-Aguet, A. Lougarre, M. Arnaud, F. Villatte, S. Estrada-Mondaca and D. Fournier, 2002. Acetylcholinesterase engineering for detection of insecticide residues. Protein Eng., 15: 43-50.
PubMed - Bull, D.L. and E.H. Ahrens, 1988. Metabolism of coumaphos in susceptible and resistant strains of Boophilus microplus (Acari: Ixodidae). J. Med. Entomol., 25: 94-98.
CrossRefDirect Link - Dzul, F.A., R. Patricia Penilla and A.D. Rodriguez, 2007. Susceptibility and insecticide resistance mechanisms in Anopheles albimanus from the Southern Yucatan Peninsula, Mexico. Salud Publica Mex., 49: 302-311.
Direct Link - Hemingway, J. and S.H. Karunaratne, 1998. Mosquito carboxylesterases: A review of the molecular biology and biochemistry of a major insecticide resistance mechanism. Med. Vet. Entomol., 12: 1-12.
PubMed - Hernandez, R., H. He, A.C. Chen, S.D. Waghela, G. Wayne Ivie, J.E. George and G.G. Wagner, 2000. Identification of a point mutation in an esterase gene in different populations of the Southern cattle tick, Boophilus microplus. Insect Biochem. Mol. Biol., 30: 969-977.
CrossRef - Jamroz, R.C., F.D. Guerrero, J.H. Pruett, D.D. Oehler and R.J. Miller, 2000. Molecular and biochemical survey of acaricide resistance mechanisms in larvae from Mexican strains of the southern cattle tick, Boophilus microplus. J. Insect. Physiol., 46: 685-695.
CrossRef - Karunaratne, S.H. and J. Hemingway, 2001. Malathion resistance and prevalence of the malathion carboxylesterase mechanism in populations of mosquito vectors of disease in Sri Lanka. Bull. World Health Organ., 79: 1060-1064.
Direct Link - Kim, C.S., W.T. Kim, K.S. Boo and S.I. Kim, 2003. Cloning, mutagenesis and expression of the acetylcholinesterase gene from a strain of Musca domestica: The change from a drug-resistant to a sensitive enzyme. Mol. Cells, 15: 208-215.
PubMed - Ortiz, E.M., V.M. Santamaria, N.A. Ortiz, C.N. Soberanes and M.J. Osorio et al., 1995. Caracterizacion de la resistencia de B. microplus a ixodicidas en México. Proceedings of the 3rd International Seminar of Parasitologia Animal, Acapulco, Mexico, October 11-13, 1995, Iconica Impresores, Morelos, Mexico, pp: 58-66.
- Paton, M.G., S.H. Karunaratne, E. Giakoumaki, N. Roberts and J. Hemingway, 2000. Quantitative analysis of gene amplification in insecticide-resistant Culex mosquitoes. Biochem. J., 346: 17-24.
PubMed - Pruett, J.H., F.D. Guerrero and R. Hernandez, 2002. Isolation and identification of an esterase from a Mexican strain of Boophilus microplus (Acari: Ixodidae). J. Econ. Entomol., 95: 1001-1007.
PubMed - Scott, J.G., 1999. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol., 29: 757-777.
CrossRefDirect Link - Soberanes, M.J., V.M. Santamaria, S.H. Fragoso and Z.V. Garcia, 2002. First case reported of Amitraz resistance in the cattle tick Boophilus microplus in Mexico. Tec. Pecu. Mex., 40: 81-92.
Direct Link - Weill, M., P. Fort, A. Berthomieu, M.P. Dubois, N. Pasteur and M. Raymond, 2002. A novel acetylcholinesterase gene in mosquitoes codes for the insecticide target and is non-homologous to the ace gene Drosophila. Proc. Biol. Sci., 269: 2007-2016.
CrossRefPubMedDirect Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Cossio-Bayugar, R., E. Miranda and P.J. Holman, 2005. Molecular cloning of a phospholipid-hydroperoxide glutathione peroxidase gene from the tick, Boophilus microplus (Acari: Ixodidae). Insect Biochem. Mol. Biol., 35: 1378-1387.
CrossRef