
M. Srinivasa Rao
Biology Division, Indian Institute of Chemical Technology,
Uppal Road, Tamaka, Hyderabad 500 007, India
U.S. N. Murty
Biology Division, Indian Institute of Chemical Technology,
Uppal Road, Tamaka, Hyderabad 500 007, India
B. Gangadasu
0rganic Chemistry Division-Il, Indian Institute of Chemical Technology, Uppal Road, Tamaka, Hyderabad 500 007, India
B. China Raju
0rganic Chemistry Division-Il, Indian Institute of Chemical Technology, Uppal Road, Tamaka, Hyderabad 500 007, India
C.H. Ramesh
0rganic Chemistry Division-Il, Indian Institute of Chemical Technology, Uppal Road, Tamaka, Hyderabad 500 007, India
S. Bharat Kumar
0rganic Chemistry Division-Il, Indian Institute of Chemical Technology, Uppal Road, Tamaka, Hyderabad 500 007, India
V. Jayathirtha Rao
0rganic Chemistry Division-Il, Indian Institute of Chemical Technology, Uppal Road, Tamaka, Hyderabad 500 007, India
Journal of Entomology
Year: 2008 | Volume: 5 | Issue: 1 | Page No.: 45-50







ABSTRACT
Mosquito larvicidal efficacy of seven different synthesized imidacloprid compounds (IMD) with different substitutions (chlorine ion, methyl group, methoxy group, ester group, phenyl ring and pyridine ring) of Imidacloprid were studied on the IV stage larvae of mosquito, Culex quinquefasciatus. LC50 of each compound (IMD1-IMD-7) was determined using probit analysis. Effects of these compounds on biochemical toxicity were also determined by observing the changes in total protein, lipid and Acetylcholine esterase activity of the mosquito larvae. The results show that due to the action of these compounds, higher amount of protein production (p<=0.001) and AchE activity (p<=0.001) and significant decrease in total lipid activity (p<=0.001) were observed when compared to the control. Similar types of results were also reported on increase production of protein, lipid and acetylcholine activity due to the action of insecticides in other insects. Hence, it is concluded that, these seven synthesized compounds can be used as mosquito larvicides.
PDF Abstract XML References Citation
How to cite this article
M. Srinivasa Rao, U.S. N. Murty, B. Gangadasu, B. China Raju, C.H. Ramesh, S. Bharat Kumar and V. Jayathirtha Rao, 2008. Larvicidal Efficacy of Neonicotinoid Classes of Compounds on Culex quinquefasciatus. Journal of Entomology, 5: 45-50.
DOI: 10.3923/je.2008.45.50
URL: https://scialert.net/abstract/?doi=je.2008.45.50
DOI: 10.3923/je.2008.45.50
URL: https://scialert.net/abstract/?doi=je.2008.45.50
INTRODUCTION
The neonicotinoids are new insecticide class, which include the commercial products imidacloprid (IMD). Its chemical analogs are also being used as insecticides for many agricultural and public health pests (Yamamoto and Casida, 1999; Mason et al., 2000) which directly act on the nicotinic acetylcholine receptor (nAchr) (Tomizawa and Casida, 2001; Suchail et al., 2003).
Imidacloprid, 1(6-chloro-3-pyridinyl)methyl-N-nitro-2-imidazolidinimine, is a new neonicotinoid insecticide, has been widely used as pest control agent on many crops (Elbert et al., 1991). Imidacloprid causes blockage in neuronal pathway which leads to the accumulation of more amount of acetylcholine, resulting to paralysis and eventually leads to death of the insect (Casida, 1998; Caroline Cox, 2001; Suchail et al., 2003). Neurophysiological studies have also shown that imidacloprid has multiple agonist and antagonist effects on neuronal nicotinic acetylcholine receptor channels in clonal rat phaeochromocytoma cells (Nagata et al., 1998). However, these compounds are shown to have very less or no toxic effect against higher organisms including humans. This selective toxicity of imidacloprid is attributed to differences in the binding affinity at the nicotinic acetylcholine receptor (Chao and Casida, 1997) or due to structural differences in the neuronal nicotinic acetylcholine receptor binding sites of mammals and insects (Casida, 1998; Wang et al., 2007).
Mosquitoes (Diptera: Culicidae) pose the greatest threat to public health because of their ability to act as vectors of pathogens causing malaria, dengue, encephalitis and filariasis, etc. which affect many millions of people all over the world, particularly in South East Asian countries. Culex quinquefasciatus Say, is one of the potential vector of Wuchereria bancrofti, the causative agent of human lymphatic filariasis (Kabir et al., 2003). Recent estimates suggest that 120 million people infected with filariasis globally (Michael, 2000). It is reported that C. quinquefasciatus and Anopheles gambiae developed resistance to many effective insecticides (Guessan, 2003). So, there is urgent need to identify some effective insecticides to suppress the mosquito control.
MATERIALS AND METHODS
Seven different synthesized analogues of imidacloprid were produced from the Department of Organic Chemistry, Indian Institute of Chemical Technology (IICT), Hyderabad and used for the present study. The process of synthesis briefly is: various substituted nicotinaldehydes were synthesized from enamides by Vilsmeier reaction. These aldehydes were reduced by using Sodium borohydride to get alcohol. Alcohols were then converted to chloride by using thonylchloride in the presence of n-Heptane solvent and catalytic amount of Dimethylformamide (DMF). These chlorides were again coupled with tetrahydro 2-nitro imine imidazole in presence of Potassium carbonate, catalytic amount of Cesium chloride and solvent was acetonitrile and yielded analogue of IMDs (substituted -2-pyridyl methyl tetrahydro-1H-2-imidazolyliden)-1-oxo-1-hydraziniumolate) (Fig. 1). These products were well characterized by 1H Nuclear Magnetic Resonance (1HNMR), Mass, IR and 13C NMR and named as Imidacloprid (IMD 1-7).
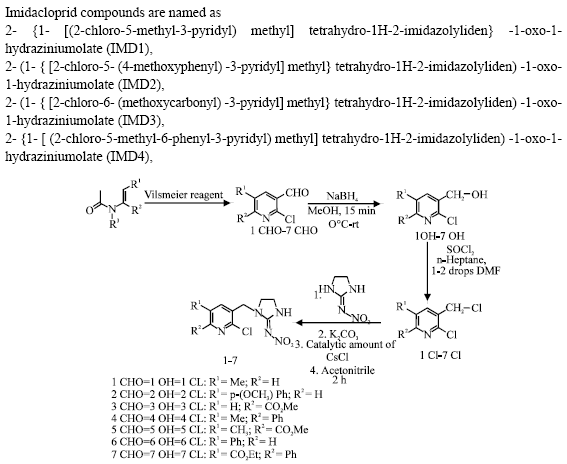 | |
| Fig. 1: | Step wise synthesized of imidacloprid analogues (IMD1-IMD7) |
Insect
Cultured stock mosquito, C. quinquefasciatus colonies from the insectary of Biology division, IICT, Hyderabad was used for this experiment. The temperature was maintained at 27±2°C and relative humidity (RH) at 60±5% throughout the experiment. Ten percent yeast suspension was provided as food source for the mosquito larvae.
Larvicidal Bioassay
The larval bioassay toxicity study was performed using standard protocol as described by World Health Organization (WHO, 1963), on 4th instar mosquito, C. quinquefasciatus larvae. For each group, 25 larvae were placed in a disposable plastic cup containing 250 mL of tap water. To determine the toxicity of the each analogue seven different concentrations, i.e., 1, 2, 3, 4, 5, 10 and 15 ppm were prepared by dissolving with acetone and were inoculated to respective cup. All the cups were coded and kept separately. Simultaneously, positive control (only IMD pure compound) and negative control (249 mL of tap water and 1 mL of acetone) were kept separately. For all the experiments, 3 replications were maintained. Larval mortality from each compound was recorded after 24 h of treatment and LC50 and LC90 were determined using Probit Analysis (Finney, 1971).
Selection of Doses
By observing the LC50 dose of each compound, one lowest and one highest concentration of each test compound were selected and tested for the biochemical analysis.
Biochemical Assays
From each set of experiment, total protein content by Lowry et al. (1951) method total lipids by Van Handel (1985) and Acetylcholine Esterase (AchE) activity using Ellman et al. (1961) method were determined. The results were subjected for statistical analysis (t-test).
RESULTS AND DISCUSSION
Larval Mortality
The LC50 values of Imidacloprid analogues were determined (IMD1, 2, 3, 4, 5, 6 and 7) were 7.15, 5.67, 6.18, 6.38, 8.86, 4.14 and 5.48 ppm, respectively. Among all the compounds, IMD-2, 6 and 7 showed higher larval mortality when compared to other 4 compounds on C. quinquefasciatus larvae (Table 1). This differential activity of these different analogues of IMD may be due to the position of the active functional groups in the native ring. When larvae were exposed to different groups of Imidcloprids analogs for 24 to 72 h post treatment of compounds, 100% of larval mortality was recorded from all the test groups including in the positive control Imidacloprid. This increased in larval mortality, may be due to the uptake of the active moiety of the compound, which could be time dependent. There is also every possibility of production of more toxic metabolites in the larval integument and alimentary canal leading to death of the larvae (Kabir et al., 2003).
Biochemical Analysis
Effect on Total Protein
The total protein content from the positive control batch exposed to only IMD was 30 μg mL-1 while in the negative control it was 23.2 μg mL-1. Higher amount of total protein were also recorded from all the different metabolites of IMD such as 54.7, 58.6, 51.8, 47, 43.2, 54.7 and 64.3 μg mL-1 in IMD-2 (5 ppm), IMD-3 (3 ppm), IMD-4 (4 ppm), IMD-5 (4 ppm), IMD-6 (2 and 3 ppm), IMD-7 (4 and 5 ppm), respectively (p≤0.001) (Table 2). This higher amount of total proteins production in presence of insecticides may be due to the over expression of the target gene (Mouches et al., 1990). A very less amount of total protein were found in IMD-3 (3 ppm)-19.2 μg mL-1 and IMD-5 (5 ppm)-16.3 μg mL-1, which may be due to the more toxic nature of these metabolites at specific concentration as per as the total amount of protein is concerned.
Effect on Total Lipid
Lipids are known to be the important constituent of the insect for the defending action against various kinds of pathogens by blocking the passage inside the body cavity. It is reported that various toxic chemicals interfere in the metabolism of lipids in animals. However, the physiological changes in insects depend upon the type of exposed toxic chemicals. In the present study, all the IMD analogues including pure IMD caused massive damage in the total lipids in mosquito. In control group, the total lipid was found to be 165.99 mg mL-1 while in pure IMD it was 45mg mL-1 (p≤0.001). In all the 7 different IMD compounds, drastic drop in the total lipid content was observed (p≤0.001) (Table 2). This decrease in the total lipid may be due to the increase in the lipid peroxidation in the plasma membrane with the exposure of these compounds.
| Table 1: | Toxicity pattern of different IMD compounds on Culex quinquefasciatus larvae |
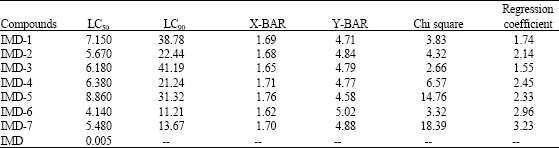 | |
Data collected from three independent replicates | |
| Table 2: | Effect of different concentrations of IMD analogues on biochemical parameters in IV stage Culex quinquefasciatus larvae |
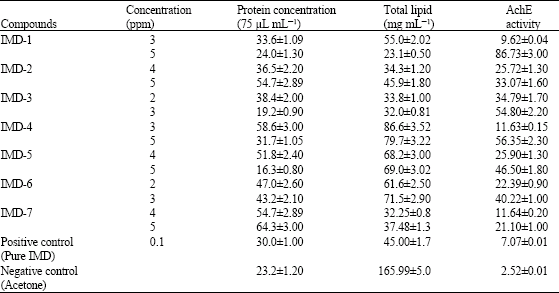 | |
Data collected from three replicates; ±is Standard Error of Mean | |
Effect on AchE
High amount of AchE were obtained with many folds in the presence of different metabolites of IMD when compared to control groups. In the negative control group, AchE was recorded 2.52 μL min-1 mg-1 while in the positive control (pure IMD), it was 7.075 μL min-1 mg-1. High concentration of the AchE were obtained in all the different IMD compounds. The amount of AchE was also increased with the increase in the concentration of the IMD metabolites tested (p≤0.001) (Table 2). This increase in the AchE in presence of the different metabolites of IMDs may be due to the inhibitory action of the AchE activity in post-synaptic region of the nerves.
CONCLUSIONS
Nicotinoids are neurotoxic in nature. These molecules bind to the receptor molecule in the nervous system that normally receives the molecule acetylcholine. Toxicological studies of Imidacloprid shows the rapid appearance of neurotoxicity symptoms in honey bee (Suchail et al., 2003). It is also reported that, IMD metabolites are more toxic than IMD (Caroline, 2001; Axel Decourtye et al., 2004). From this study it is observed that, the toxicity pattern of all the tested compounds do not differ much and on par with the toxicity of the (IMD). Among all the analogues tested, IMD-6, IMD-7 and IMD-2 (chlorine atom with different position and contain phenyl substitution), (ester group and substitution of pyridine ring) and methoxy group) respectively showed higher toxic effect on C.quinquefasciatus larvae when compared to other four compounds. To use these compounds as mosquito larvicides, further studies on non-target organisms are required.
ACKNOWLEDGMENT
Authors are thankful to the Director, Indian Institute of Chemical Technology for his encouragement and support.
REFERENCES
- Chao, S.L. and J.E. Casida, 1997. Interaction of imidacloprid metabolites and analogs with the nicotinic acetylcholine receptor of mouse brain in relation to toxicity. Pestic. Biochem. Physiol., 58: 77-88.
CrossRefDirect Link - Decourtye, A., C. Armengaud, M. Renou, J. Devillers, S. Cluzeau, M. Gauthier and M. Pham-Delague, 2004. Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pesticide Biochem. Physiol., 78: 83-92.
CrossRefDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Finney, D.J., 1971. Probit Analysis. 3rd Edn., Cambridge University Press, London, UK., pp: 76-80.
CrossRefDirect Link - Guessan, N.R. and F. Darriet, P. Guillet and P. Carnevale et al., 2003. Resistance to carbosulfan in Anopheles gambiae from Ivory Coast based on reduced sensitivity of acetylcholinesterase. Med. Vet. Entomol., 17: 19-25.
Direct Link - Kabir, K.E., A.R. Khan and M.A. Mosaddik, 2003. Goniothalamin-a potent mosquito larvicide from Bryonopsis laciniosa L. J. Applied Entomol., 127: 112-115.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mason, G., M. Rancati and D. Bosco, 2000. The effect of thiamethoxam, a second generation neonicotinoid insecticide, in preventing transmission of Tomato yellow leaf curl geminivirus (TYLCV) by the whitefly Bemisia tabaci (Gennadius). Crop Prot., 19: 473-479.
Direct Link - Mouches, C., Y. Pauplin, M. Agarwal, L. Lemieux and M. Herzog et al., 1990. Characterization of amplification core and esterase B1 gene responsible for insecticide resistance in culex. Proc. Nat. Acad. Sci. USA., 87: 2574-2578.
PubMedDirect Link - Nagata, K., J.H. Song, T. Shono and T. Narahashi, 1998. Modulation of the neuronal nicotinic acetylcholine receptor-channel by the nitromethylene heterocycle imidacloprid. J. Pharmacol. Exp. Ther., 285: 731-738.
PubMedDirect Link - Suchail, S., L. Debrauwer and L.P. Belzunces, 2004. Metabolism of imidacloprid in Apis mellifera. Pest Manage. Sci., 60: 291-296.
Direct Link - Tomizawa, M. and J.E. Casida, 2001. Structure and diversity of insect nicotinic acetylcholine receptors. Pest Manage. Sci., 57: 914-922.
Direct Link - Van Handel, E., 1985. Rapid determination of glycogen and sugars in mosquitoes. J. Am. Mosquito Control Assoc., 1: 299-301.
PubMed - Wang, Y., Jiagao Cheng, Xuhong Qian and Zhong Li, 2007. Actions between neonicotinoids and key residues of insect nAChR based on an ab initio quantum chemistry study: Hydrogen bonding and cooperative π-π interaction. Bioorgan. Med. Chem., 15: 2624-2630.
Direct Link